
Thouarella moseleyi Wright and Studer, 1889
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3602.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:10304FBF-3969-4EFA-83F1-BB8A5E2B37F3 |
|
persistent identifier |
https://treatment.plazi.org/id/EE36E867-FFC9-FFBB-FF0A-ADB5FCEF0FA9 |
|
treatment provided by |
Felipe |
|
scientific name |
Thouarella moseleyi Wright and Studer, 1889 |
| status |
|
19. Thouarella moseleyi Wright and Studer, 1889 View in CoL
Figs 30 View FIGURE 30 , 31 View FIGURE 31
Thouarella moseleyi Wright & Studer, 1889: 61–62 View in CoL , pl. 14, fig. 1, 1a, pl. 21, fig. 2; Versluys 1906: 29–30, figs 26, 27 (in text) Thouarella (Euthouarella) moseleyi Kükenthal 1915: 150 View in CoL ; 1919: 417–418; 1924: 292, 294; Cairns & Bayer 2009: 28 (listed) Thouarella moseleyi var. spicata Thomson & Henderson, 1906: 38 , 42–43, pl. 3, figs 2, 4 (sample not seen)
Material examined: Holotype, NHM 1889.5 .27.39, H.M.S. Challenger, sta. 171, Kermadec Islands , NE of New Zealand, 28˚33’S, 177˚50’E, 1097 m, 15 Jul 1874, 6 cm colony.
Description
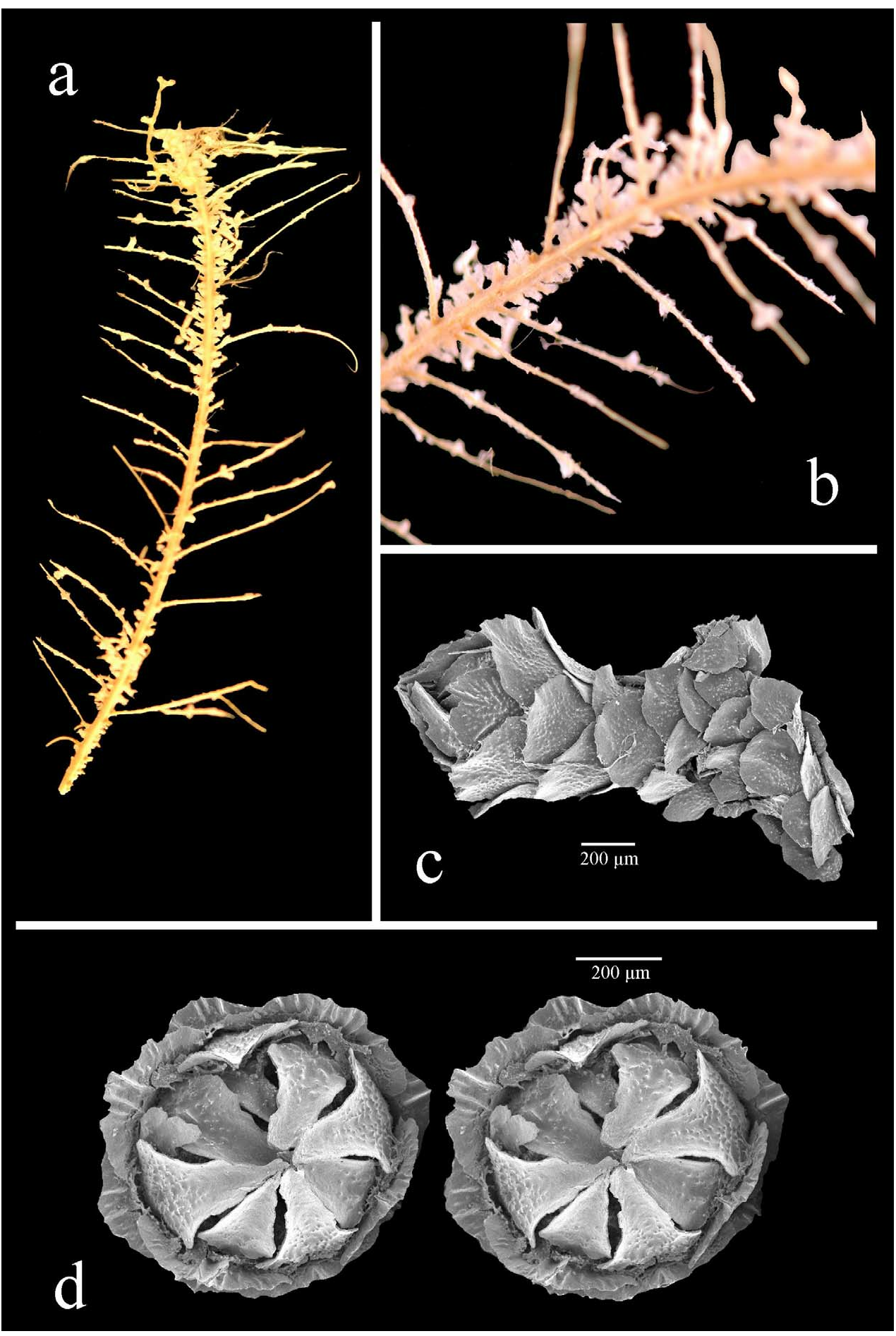
The holotype was originally approximately 160 mm long (from Wright & Studer 1889: pl. 14, fig. 1) but only a 60 mm fragment now remains. The branchlets are pinnate, possibly alternately pinnate ( Fig. 30a,b View FIGURE 30 ), although the colony can appear bottlebrush as branchlets are flexible and curve in all directions. The branchlets are mostly simple, some with secondary branching and they are 15–20 mm long. The axis is thin, calcareous, flexible, and “somewhat flattened” ( Wright & Studer 1889).
The polyps are clavate with a tall, conical operculum ( Fig. 30c View FIGURE 30 ), 1.5 mm high, paired, standing perpendicular to the branchlet, with some on the main stem. Each polyp has 6 longitudinal rows of body-wall scales and that number reduces rapidly adaxially from the marginal. There are 4–5 scales in the longitudinal abaxial row, and 3–4 in the adaxial rows.
The thin, lanceolate operculars (can be wider with rounded distal edge, Fig. 31b View FIGURE 31 ) measure 450–640 µm high (average 510 µm), 140–270 µm wide (average 200 µm), with a H:W of 2.2–3.1 (average 2.6). The inner surface is smooth with a simple keel running longitudinally from the tip for three-quarters of the scale length ( Fig. 31c View FIGURE 31 ) whilst the proximal quarter is tuberculate. Wider operculars have a wider, ridged area on the inner surface rather than a distinct keel ( Fig. 31b View FIGURE 31 ). The outer scale surface is longitudinally concave and smooth, with granules arranged radially from the proximal centre and tubercles are visible at the proximal edge. The opercular edge appears smooth, but may be worn and thus not representative.
The marginals are twice as wide as the operculars, 400–410 µm (average 406 µm), 455–510 µm high (average 480 µm), with a H:W of 1.1–1.3 (average 1.2), and are diamond-shaped ( Fig. 31d–f View FIGURE 31 ). The inner surface has 4 or 5 tall, thin keels ( Fig. 31d,f View FIGURE 31 ) with smooth areas lateral to the keels whilst the remainder is tuberculate. The outer surface is longitudinally concave and smooth, with granules distributed radially from the proximal centre with some tubercles being visible at the proximal edge ( Fig. 31e View FIGURE 31 ). The distal edge is irregular, possibly worn, and the proximal edge is coarsely lobate.
The submarginals are fan-shaped to elliptical ( Fig. 31g,h View FIGURE 31 ), 320–420 µm high (average 383 µm), 470–480 µm wide (average 475 µm), with a H:W of 0.7–0.9 (average 0.8). The inner surface is tuberculate, the distal edge having a wide area of perpendicular ridges rather than a keel. The outer surface and scale edges are as the bodywall scales, below.
The body-wall scales are circular to fan-shaped ( Fig. 31i–l View FIGURE 31 ), 300–400 µm high (average 340 µm), 300–475 µm wide (average 370 µm), with a H:W of 0.7–1.2 (average 0.9). The inner surface is tuberculate with a narrow smooth band along the distal edge, and occasionally, small teeth perpendicular to the distal edge. The outer scale surface is smooth with sparse granules across the proximal area whilst the distal edge is smooth and the proximal edge is coarsely lobate.
There are 2 layers of coenenchymal scales ( Fig. 31m View FIGURE 31 ): sclerites from the inner layer are circular in shape and sclerites from the outside layer are elliptical. The inner surface of the coenenchymal scales is tuberculate with a smooth band running along the distal edge whilst the outer surface is smooth, sometimes with sparsely placed granules. The distal edge is smooth and the proximal edge is coarsely lobate.
Distribution
This species is known from 2 locations (excluding var. spicata ), northwest of the Kermadec Islands in the South Pacific and north of Sumbawa Island in Indonesia, 6600 km away. It has been recorded from depths of 794–1097 m.
Remarks
The number of polyps per cm was impossible to tell from the small holotype fragment that remains. More material of this species from the type locality is required to confirm the differences between it and T. laxa , which has a very similar polyp morphology and pinnate branching.
A variety of T. moseleyi ( var. spicata ) was described in Thomson and Henderson (1906). The differences between the polyps of the holotype and those of the variant are mainly that the latter has a taller conical operculum and marginal scales with a more elongated, pointed spine (which may be bifid). The tall conical operculum is similar to that of the polyps of T. grasshoffi . However, colonies of T. grasshoffi have a true bottlebrush branching arrangement.
Comparisons
Within Group 2 only two other species, T. laxa and T. tydemani , have pinnate colonies or colonies that appear pinnate. The ridges running perpendicular to the distal edge of the body-wall scales on the polyps of T. moseleyi are similar to those reported on the body-wall scales in a recent description of T. tydemani ( Zapata-Guardiola and López-González 2010a) . The polyps of T. tydemani , however, have long spinose marginals and are distally flared, whereas polyps of T. moseleyi are clavate with shorter marginals.
The polyps of T. moseleyi are more robust than those of T. laxa , with wider marginals that are not as spinose, and a fitted opercular cone. There are also several ridges running perpendicular to the distal edge of the submarginals, which are absent in the polyps of T. laxa .

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Thouarella moseleyi Wright and Studer, 1889
| TAYLOR, M. L., CAIRNS, S. D., AGNEW, D. J. & ROGERS, A. D. 2013 |
Thouarella moseleyi
| Cairns, S. D. & Bayer, F. M. 2009: 28 |
| Kukenthal, W. 1924: 292 |
| Kukenthal, W. 1919: 417 |
| Kukenthal, W. 1915: 150 |
| Versluys, J. 1906: 29 |
| Thomson, J. A. & Henderson, W. D. 1906: 38 |
| Wright, E. P. & Studer, T. 1889: 62 |