
Woodworthia korowai, Winkel & Wells & Harker & Hitchmough, 2023
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5374.2.7 |
|
publication LSID |
lsid:zoobank.org:pub:973FE7A1-91F3-40DC-8CC7-A3DAA36C046C |
|
DOI |
https://doi.org/10.5281/zenodo.10248451 |
|
persistent identifier |
https://treatment.plazi.org/id/8A9D11D1-82AC-4C37-86BE-2FA7D156B736 |
|
taxon LSID |
lsid:zoobank.org:act:8A9D11D1-82AC-4C37-86BE-2FA7D156B736 |
|
treatment provided by |
Plazi |
|
scientific name |
Woodworthia korowai |
| status |
sp. nov. |
Woodworthia korowai sp. nov.
Figs. 5–10 View FIGURE 5 View FIGURE 6 View FIGURE 7 View FIGURE 8 View FIGURE 9 View FIGURE 10 .
ZooBank registration of Woodworthia korowai sp. nov.: urn:lsid:zoobank.org:act:8A9D11D1-82AC-4C37-86BE-2FA7D156B736 ..
Woodworthia aff. maculata “Muriwai” Hitchmough et al. (2016)
Woodworthia aff. maculata “Muriwai” van Winkel et al. (2018)
Woodworthia aff. maculata “Muriwai” van Winkel et al. (2020)
Woodworthia aff. maculata “Muriwai” Hitchmough et al. (2021)
Woodworthia aff. maculata “Muriwai” Purdie (2022)
Woodworthia aff. maculata “Muriwai” Melzer et al. (2022)
Holotype. AWMM LH02445 (adult female) from Muriwai Beach , Auckland, New Zealand (36°47’49.69”S, 174°24’32.34”E; ~ 1 km accuracy; ~ 5 m a.s.l.) collected by S. E. Thorpe on 6 September 2003. GoogleMaps
Paratypes (three specimens). AWMM LH4067 (adult female) from Oaia Island , Muriwai, Auckland, New Zealand; 36°50’26.32”S, 174°24’39.79”E; 20 m a.s.l.; collected by D. van Winkel and C. J. Wedding on 5 April 2014 GoogleMaps . AWMM LH4068 (adult male) from Muriwai Beach , Auckland, New Zealand; 36°47’4.27”S, 174°24’1.19”E; 5 m a.s.l.; by D. van Winkel on 18 November 2020 GoogleMaps . AWMM LH4069 (adult male) from Woodhill Forest , Auckland, New Zealand; 36°41’59.61”S, 174°21’20.37”E; 19 m a.s.l.; collected by an anonymous person, surrendered to N. Harker in December 2012 GoogleMaps .
Diagnosis. A combination of transversely expanded and V-shaped rather than straight lamellae, small body size (≤ 100mm SVL), and rostral scale excluded from nares, as well as its phylogenetic position identify the species as a member of the genus Woodworthia . Woodworthia korowai sp. nov. can be distinguished from all other Woodworthia species by the following combination of characters: adult body length up to 68 mm SVL; rostral scale not in contact with nares, separated by suture between anterior supranasal and first infralabial; rostral scale (<2.5x versus> 2.5x as wide as deep); 13–15 subdigital lamellae under the fourth digit of the pes; and 3–5 (versus> 5) scale rows under the distal phalanx of the fourth toe ( Fig. 9A View FIGURE 9 ). It is distinguished from W. chrysosiretica and W. “southern mini” by a narrower and deeper rostral scale (<2.5x versus> 2.5x as wide as deep), and further from W. “southern mini” by the higher number of subdigital lamellae under the fourth digit of the pes (13–15 versus 8–13). It can be distinguished from W. “Mount Arthur” by the following suite of characters: 13–15 (versus 9–12) lamellae under the fourth toe; 3–5 scale rows (versus 5–6, Fig. 9E View FIGURE 9 ) under the distal phalanx of the fourth toe; and on average: relatively narrower head (0.66 versus 0.71 HW/HL); shorter body proportions (0.86 versus 1.04 SF/AG); and shorter distal phalanx on fourth toe (0.24 versus 0.34 4TDPL/4FTL). It is distinguished from Woodworthia maculata sensu stricto by the following suite of characters: 9–11 supralabial scales (versus 10–14); 8–11 infralabial scales (versus 9–14); 3–5 scale rows (versus 5–8; Figs 9C & D View FIGURE 9 ) under the distal phalanx of the fourth toe; and on average: relatively narrower head (0.66 versus 0.72 HW/HL); smaller eye (0.19 versus 0.21 ED/HL); shorter fourth toe length (0.10 versus 0.12 4TL/SVL); and shorter distal phalanx on fourth toe (0.24 versus 0.30 4TDPL/4FTL). Table 6 View TABLE 6 compares characters of Woodworthia korowai sp. nov. with its two most closely related congeners. Diagnostics are not given for W. chrysosiretica , nor for any other undescribed Woodworthia species as formal taxonomic descriptions for these are in preparation (R. Hitchmough, S.V. Nielsen and A. Bauer, unpub. data). None of the undescribed taxa resemble W. korowai sp. nov. and all are allopatrically distributed.
Description of holotype ( Figs. 5 View FIGURE 5 & 6 View FIGURE 6 ). Adult female SVL 57.4 mm; head large, moderate in length (HL/SVL 0.28) and width (HW/HL 0.70), moderately depressed (HD/HL 0.5), distinct from neck, and triangular in dorsal profile; snout elongate (SNT/HL 0.39), broader than long, rounded in dorsal profile and truncate in lateral profile; eye to nostril distance greater than eye diameter (EN/ED 1.47); loreal region weakly inflated, prefrontal region slightly concave; canthus rostralis flat, weakly defined; eye large (ED/HL 0.20), pupil vertical, supraciliaries extending from anteroventral to posterodorsal edge of orbit; ear opening inverted oblong-shaped (EW/ED 0.44), longer than wide (EL/EW 1.43); eye-to-ear distance greater than diameter of eye (EE/ED 1.75); rostral trapezoid-shaped, wider than high (<2.5x as wide as high), with medial suture extending ventrally approximately halfway from dorsal edge, bordered dorsally by two domed supranasals and one small inverted pentagonal internasal, broader than long; rostral not in contact with nares, separated by suture between anterior supranasal and first infralabial (0.57x length of the nares); nares bordered anteriorly by large supranasal (2x size of other supranasals), dorsally and posteriorly by three postnasals and ventrally by first supralabial; 11 square supralabials, rounded/ curved dorsally, extending to and tapering smoothly below posterior margin of orbit, 8 to midpoint of eye; 10 infralabials tapering smoothly posteriorly to below posterior margin of orbit; scales of rostrum, loreal region, top of head, and occiput small and granular, those of the rostrum and loreal region approximately 1.5x the width of those on top of head, occiput, and posterior nuchal regions; superciliaries forming a brillar fold of small spiniform scales extending along the dorsal border of the orbit from anteroventral to posterodorsal corners; mental triangular, longer than wide, bordered laterally by first infralabials and posteriorly by medial triangular postmental; one row of slightly enlarged chinshields tapering posteriorly to fifth infralabial; and gular and throat scales small, granular, grading posteriorly into sub-flattened, mostly juxtaposed but few subimbricate pectoral and ventral scales. Body relatively short (AG/SVL 0.51) with no obvious ventrolateral folds; dorsal scales small, granular, homologous; ventral scales sub-flattened, mostly juxtaposed but some subimbricate, distinctly broader than long, much larger than dorsal scales, increasing in size medially, arranged in approximately 35 rows at midpoint of body. Forelimbs robust in stature, short (HUM+FEM/SVL 0.22); granular scales of dorsal and anterior margins of forearm larger (2.5x) than those on body; palmar scales weakly domed to sub-flattened, juxtaposed; digits relatively short, well-developed, moderately expanded, all bearing claws; those on digit I reduced, remaining claws long and strongly recurved; claws sheathed by a dorsal and ventral scale; relative length of digits of manus: IV ~ III>V ~ II> I; subdigital lamellae straight or slightly curved becoming strongly V-shaped distally on the dilated portion, smooth, undivided; lamellar counts from right (and left) sides 6-10-12-14-10 (5-9-12-12-11) manus (excludes apical scansors of digit I); reduced claw of digit I of manus situated in a groove in the apical lamella between a larger medial scansor and a smaller lateral scansor. Hind limbs short (FEM+TIB/SVL 0.31), more robust and longer than forelimbs, covered dorsally by granular scales and anteriorly by larger (2.5x than those on body), weakly domed, sub-imbricate scales; ventral scales of femora flattened, juxtaposed, equal in size to those of ventral scales; small postfemoral scales form an abrupt union with larger ventral scales of posteroventral margin of thigh; subtibial scales small, granular, juxtaposed; plantar scales weakly domed to sub-flattened, juxtaposed; digits relatively short (4TL/SVL 0.10), well developed, moderately expanded, all bearing claws; those on digit I reduced and partially sheathed, remaining claws long and strongly recurved; claws sheathed by a dorsal and ventral scale; relative length of digits of pes: IV> V> III> II> I; subdigital lamellae straight or slightly curved becoming strongly V-shaped distally on the dilated portion ( Fig. 9A View FIGURE 9 ), smooth, undivided, lamellar counts from right (and left) sides 5-9-13-14-12 (5-10-12-14-11) pes (excludes apical scansors of digit I); reduced claw of digit I of pes situated in a groove in the apical lamella between a larger medial scansor and a smaller lateral scansor.
Eight rows of precloacal scales, equal in size to ventral scales, extending approximately halfway along the length of the femora; no precloacal or femoral pores (some medial scales weakly dimpled); nine rows of smaller (0.5x precloacal scales) post-precloacal scales; six rows of enlarged, flattened hexagonal or roundish postcloacal scales; single enlarged, round, flattened postcloacal spur (2–3x surrounding scales) on each side of the tail base. Complete (unregenerated) tail, longer than SVL (TL/SVL 1.16); tail thick, roughly round in cross section, tapering to a point. Caudal scales flat, juxtaposed to weakly subimbricate, squarish with rounded free margins, arranged in regular rows. Surface of tail weakly segmented, caudal scale rows forming whorls, each whorl 6 ventral scale rows long; ventral caudals 2x larger than dorsals, midventral caudal scales not enlarged.
Colour of holotype in preservative. Ground colour of the head, body, and limbs tawny brown, and tail a lighter tan or buff; top of head with broad buff inverted ‘U’ marking running posterior to the eyes and onto the nape; pale buff dorsolateral stripes, continuous from the nares over the eyes to the base of the tail, and converging along the length of the tail; darker brown medial stripe from tip of snout to in line with the anterior margins of the eye; darker brown pre- and postorbital stripes, commencing at the nares and continuing the length of the body, bordering the dorsolateral stripes ventrally, to the base of the tail; lateral surfaces speckled with lighter and darker flecks and gradually fading to uniform cream ventrally; indistinct and broken buff mid-dorsal stripe from the nape to the base of the tail; limbs speckled with light and dark blotches and flecks; ventral surfaces, including base of pedes and manus, uniform yellow cream; dorsal surface of tail with a series of irregular light blotches; labial scales buff with murky brown irregular blotches; eyes brown with dark filigree pattering.
Colouration in life (non-vouchered specimens photographed in wild; Fig. 10 View FIGURE 10 ). Ground colour of the head, body, and limbs tawny brown, olive brown or grey-brown, and tail a lighter shade of body colour; top of head either indistinctly marked or with distinct dark inverted ‘V’ marking between the eyes often continuing posteriorly to form a heart-shaped, circular, or inverted ‘V’ marking on the back of the head; broad buff inverted ‘U’ marking running posterior to the eyes and onto the nape; pale buff dorsolateral stripes, continuous from the nares over the eyes to the base of the tail, and converging either at the tail base or along the length of the tail; dorsolateral stripes often wavy or with transverse projections that contact the mid-dorsal stripe; dorsal surface rarely without dorsolateral stripes, rather with transverse irregular-shaped pale blotches between the nape and the tail base; darker brown medial stripe from tip of snout to in line with the anterior margins of the eye; darker brown pre- and postorbital stripes, commencing at the nares and continuing the length of the body, bordering the dorsolateral stripes ventrally, to the base of the tail; lateral surfaces speckled with lighter and darker and occasionally mustard yellow flecks, and gradually fading to uniform cream ventrally; indistinct and broken buff mid-dorsal stripe from the nape to the base of the tail, or mid-dorsal absence; limbs speckled with light and dark blotches and flecks; ventral surfaces, including base of pedes and manus, uniform yellow cream; subdigital lamellae often lighter shade of grey; dorsal surface of tail with a series of irregular light blotches; regenerated tails often longitudinally striped with alternating darker and lighter brown stripes; labial scales buff coloured with murky brown irregular blotches; eye colour green, olive green, or yellow green with black web or filigree patterning.
Variation. The paratypes closely resemble the holotype in all aspects of colouration and pattern ( Fig. 7 View FIGURE 7 ), except AWMM LH4067 differs by having much less pronounced markings on the snout and top of head. AWMM LH4067 has damage to the outer integument on the left forearm and over the pelvis, which are artifacts of wild collection. Two of the paratypes (AWMM LH4069 and AWMM LH4068) have partially regenerated tails. AWMM LH4068 is missing the fifth digit of the left pes and there is a small hole on the ventral surface of the abdomen, anterior to the vent, that is an artifact of post-mortem decomposition prior to preservation. Two of the paratypes are males with precloacal pores: AWMM LH4069 has seven rows of precloacal scales; 62 pore bearing scales in total, in four rows, 3 extending onto the thigh, 2 extending approximately halfway along the length of the femora, equal in size to the ventral scales; ten rows of smaller (0.5x precloacal scales) post-precloacal scales; eight rows of enlarged, flattened hexagonal or roundish postcloacal scales; single enlarged, round, flattened postcloacal spur (3x surrounding scales) on each side of the tail base. AWMM LH4068 has seven rows of precloacal scales; 81 pore bearing scales in total, in four rows, 3 extending onto the thigh, 2 extending approximately halfway along the length of the femora, equal in size to the ventral scales; ten rows of smaller (0.5x precloacal scales) post-precloacal scales; eight rows of enlarged, flattened hexagonal or roundish postcloacal scales; single enlarged, round, flattened postcloacal spur (4x surrounding scales) on each side of the tail base ( Fig. 8 View FIGURE 8 ). AWMM LH4067 has seven rows of precloacal scales, equal in size to ventral scales, extending approximately halfway along the length of the femora; no precloacal or femoral pores; eight rows of smaller (0.5x precloacal scales) post-precloacal scales; six rows of enlarged, flattened hexagonal or roundish postcloacal scales; single enlarged, round, flattened postcloacal spur (2–3x surrounding scales) on each side of the tail base. no pore bearing scales in total, in four rows, 3 extending onto the thigh, 2 extending approximately halfway along the length of the femora, equal in size to the ventral scales; ten rows of smaller (0.5x precloacal scales) post-precloacal scales; nine rows of enlarged, flattened hexagonal or roundish postcloacal scales; no visible cloacal spurs. Mensural and meristic differences among specimens of the type series are presented in Table 6 View TABLE 6 .
Distribution. Woodworthia korowai sp. nov. is currently known only from the western coastline from Muriwai township north onto Te Korowai-o-Te-Tonga/ South Kaipara Peninsula (reaching 4.5 km south of the Kaipara Harbour entrance), in Auckland Region, New Zealand. A small, isolated population also occurs on Oaia Island, approximately 1.3 km off the coast of Muriwai township. Woodworthia korowai sp. nov. has not been recorded on any of the other small islands that lie off the west Auckland coastline between Muriwai and the Manukau Harbour entrance to the south. The presence of Woodworthia korowai sp. nov. elsewhere on the Auckland mainland (e.g., areas further south to the Manukau Inlet or northwards along the Kaipara Harbour coast) cannot be excluded, nor can its potential presence in southern Northland. However, dedicated surveys outside of the existing known range (e.g., southern Auckland Region, Kaipara, and southern Northland) have thus far failed to detect this species (D. van Winkel & Auckland Zoo, unpub. data). Until demonstrated otherwise Woodworthia korowai sp. nov. is considered endemic to the Auckland Region.
Etymology. The specific epithet is from the te reo Māori word korowai, referencing Te Korowai-o-Te-Tonga, the traditional name for South Head or South Kaipara Peninsula where the stronghold of this species occurs. Korowai is also a Māori term for a cloak, some of which the colour and patterns closely resemble those of the gecko. Furthermore, the English translation of Te Korowai-o-Te-Tonga (“the cloak of the south”) represents a metaphor for the covering, hiding, or concealing the species’ existence until relatively recently. The specific epithet was given by Ngāti Whātua o Kaipara, the mana whenua (the indigenous Māori people of New Zealand who have historic and territorial rights over the land) of Te Korowai-o-Te-Tonga.
Suggested vernacular name. The currently accepted vernacular name is ‘Muriwai gecko’ ( Bell 2014; van Winkel et al. 2018; Hitchmough et al. 2021; Purdie 2022) however, we propose changing it to ‘Korowai gecko’ to align with the specific epithet.
Environment. The Auckland Region lies at S36.8° (approximately 13° of latitude south of the Tropic of Capricorn) and experiences a subtropical climate characterised by warm and humid summers (December to February) and relatively mild winters (June to August). Mean annual temperatures range between 14 °C and 16 °C and relative humidity is high in all seasons (78–92%) due to the influence of the surrounding sea and the lack of any large mountain masses ( Chappell 2013). Most parts of Auckland receive around 2,000 hours of bright sunshine per year and median annual total rainfall for the west coast areas of the region range between 1,100 –1,500 mm, with most of the rainfall falling in the winter months ( Chappell 2013). The region regularly encounters storms of tropical origin, and the west coast is exposed to ocean swells emanating from the Tasman Sea and the Southern Ocean, which are further disturbed by the prevailing southwest winds. The coastal distribution of Woodworthia korowai sp. nov. means populations are frequently exposed to high winds, storm surges, and salt spray off the ocean. During the largest storms, Oaia Island is partially washed over by waves approaching from the west/ southwest.
Habitat. Woodworthia korowai sp nov. is largely restricted to coastal habitats such as duneland, coastal shrubland, and exotic Monterey pine ( Pinus radiata ) plantation on mainland New Zealand and exposed rock and prostrate herbfields on Oaia Island ( Figs. 1 View FIGURE 1 & 10C–E View FIGURE 10 ). The mainland duneland environment is characterised by large accumulations of sand build up that are stabilised by low-growing coastal vegetation. The vegetation comprises a relatively low diversity of drought-tolerant plant species including spinifex ( Spinifex sericeus ), pīngao ( Ficinia spiralis ), sand tussock ( Poa billardierei ), shore bindweed ( Calystegia soldanella ), tātaraheke/ sand coprosma (Coprosma acerosa ), tauhinu ( Ozothamnus leptophyllus ), and coastal toetoe ( Austroderia splendens ). The vegetation is typically dense and low growing through the foredunes but becomes noticeable thicker and shrubbier as it progresses into the hind dunes. The vegetation then develops abruptly into Monterey pine plantation forest with predominantly an exotic weed-dominated understory (e.g., woolly nightshade, Solanum mauritianum and pampas grass, Cortaderia selloana ). Wilding Monterey pine encroaches part way into the hind dunes and in some areas the sand accumulations have reached the canopy of the trees. The coastal foredunes are scattered with large sand blows. These accumulations of sand are dynamic and mostly devoid of vegetation, although isolated sand hummocks supporting clumps of coastal toetoe are occasionally present, and these are often occupied by Woodworthia korowai sp. nov. Around the perimeter of the sand blows, dense vegetation such as coastal toetoe, small-leaved pōhuehue ( Muehlenbeckia complexa var. complexa ) and exotic pampas and tree lupin ( Lupinus arboreus ) grow. The species has been reported on the edges and in clearings within Monterey pine plantation forest that abuts coastal duneland and from more urban settings (e.g., residential gardens and dwellings) in Muriwai township.
On Oaia Island (0.53 ha), the habitat is vastly different and is dominated by exposed rock, with a relatively small area (approximately 0.07 ha) of low-growing horokaka ( Disphyma australe ) and pigweed ( Chenopodium trigonon subsp. trigonon ) and low-spreading, woody stemmed taupata ( Coprosma repens ) skirting the mid-eastern flank of the island. Oaia Island is steep-side rock comprised of early Miocene-aged pillow lava, dated at c. 16-17 million years ago ( Cameron & Taylor 1989). The island is situated within the breaker/ surf zone and is 1.4 km from the nearest projection of the Waitakere coast and is partially washed over by large waves during the heaviest storms. The distribution and abundance of Woodworthia korowai sp. nov. on the island is clearly influenced by environmental factors, particularly the exposure to wave action and sea spray. The geckos largely occupy the lee (eastern) side of the island and are very scarce or absent on the western (ocean) side, except within some deep crevices on the upper third of the western rock faces. The total area of occupancy on the Island is estimated at less than half the total area of the island.
Natural History. Woodworthia korowai sp. nov. is primarily nocturnal but will sun-bask cryptically during the day, by exposing small portions of its body or head to direct sunlight from beneath dense vegetation or refuge structures. When active, this species can move with speed and is able to leap short distances (N. Harker, pers. obs.). Little is known about its movements and home range area; however, individuals have been reported moving 10–20 m between vegetation clumps and one individual was documented moving 20 m across an open sand dune between two patches of coastal toetoe over a period of 24 hours (D. van Winkel, unpub. data). Woodworthia korowai sp. nov. is viviparous and females give birth to 1–2 offspring annually in late austral summer/ early Autumn (February–April). Offspring size at birth is approximately 35 mm SVL. Age to maturity is unknown but presumed to be around 3–4 years, like its closely related sister species Woodworthia maculata . Life expectancy is unknown but likely to be several decades as reported in other Woodworthia species ( Lettink & Whitaker 2006; Cree & Hare 2016).
The diet consists of small invertebrates, including beetles (e.g., Mimopeus elongatus ), cockroaches (e.g., Drymaplaneta heydeniana ) and the small fruits (berries) of plants such as Muehlenbeckia complexa (presence of seeds confirmed in scats; D. van Winkel, pers. obs.) and possibly tātaraheke/ sand coprosma ( Coprosoma acerosa ) and native spinach ( Tetragonia trigyna ) when seasonally available. On Oaia Island, the diversity and abundance of invertebrates is much more limited, and the diet is likely dominated by few invertebrates such as matā/ seashore earwig ( Anisolabis littorea ) and other flighted insects (e.g., dipterans). The omnivorous diet is a common feature of many New Zealand geckos ( Hare et al. 2016; Wotton et al. 2016).
TABLE 6. (continued)
TABLE 6. (continued)
Syntopic lizard species that occur in the duneland habitat of W. korowai sp. nov. at the type locality are Oligosoma aff. smithi “Three Kings, Te Paki, Western Northland ” ( Hare, Daugherty & Chapple, 2008) and introduced and naturalised Lampropholis delicata ( De Vis, 1888) . These species share the same micro-habitats in the duneland ecosystem, being closely associated with complex low-growing vines and dense vegetation clumps (e.g., pampas and coastal toetoe). On Oaia Island, W. korowai sp. nov. is the only lizard present.
Potential predators of W. korowai sp. nov. include birds (e.g., pūkeko, Porphyrio melanotus and kōtare/ sacred kingfisher, Todiramphus sanctus ) and introduced mammals (house mouse, Mus musculus ; European hedgehog, Erinaceus europaeus ; ship rat, Rattus rattus ; Norwegian rat, R. norvegicus ; feral pig, Sus scrofa; ferret, Mustela furo ; stoat, M. erminea ; weasel, M. nivalis ; feral cat, Felis catus ; and potentially brushtail possum, Trichosurus vulpecula ).
Conservation status. Woodworthia korowai sp. nov. is currently listed (under the tag-name Woodworthia aff. maculata “Muriwai ”) in the New Zealand Threat Classification System ( Townsend et al. 2008) as ‘Nationally Vulnerable’ (with Qualifiers: Climate Impact, Conservation Research needed, Data Poor Size, Data Poor Trend, Range Restricted) on the basis of criterion C (1) (moderate population, 1,000 –5,000 mature individuals, predicted decline 10–50% over three generations) (Hitchmough et al. 2021). Generation time is estimated at about 8–10 years, based on an estimate of three to four years to reach sexual maturity.
Woodworthia korowai sp. nov. is currently listed (under the tag-name Woodworthia aff. maculata “Muriwai ”) in the Tāmaki Makaurau/ Auckland regional threat classification system as ‘Regionally Vulnerable’ (Data Poor Trend, Range Restricted, Climate Impact, Conservation Research needed, National Stronghold, Regional Endemic) on the basis of criterion C (1) (moderate population, 1,000 –5,000 mature individuals, predicted decline 10–50% over three generations)) ( Melzer et al. 2022).
In the IUCN red-list system (IUCN 2012), we suggest that W. korowai sp. nov. justifies listing as Endangered, on the basis of the criteria B1a,b(v), B2a,b(v); i.e., extent of occurrence estimated to be less than 5,000 km 2, area of occupancy estimated to be less than 500 km 2, it is known to exist at no more than five locations, and with a continuing decline inferred in numbers of mature individuals. Decline is inferred based on impacts of invasive mammalian predators, which are ubiquitous in their negative impacts on native lizards throughout mainland New Zealand ( Whitaker 1973; Nelson et al. 2016), and on-going degradation of coastal habitats as a result of anthropogenic activities (e.g., illegal four-wheel drive vehicles), exotic herbivores (e.g., Dama dama ; Oryctolagus cuniculus ), wilding pine encroachment, fires, and climate change impacts (e.g., coastal erosion).
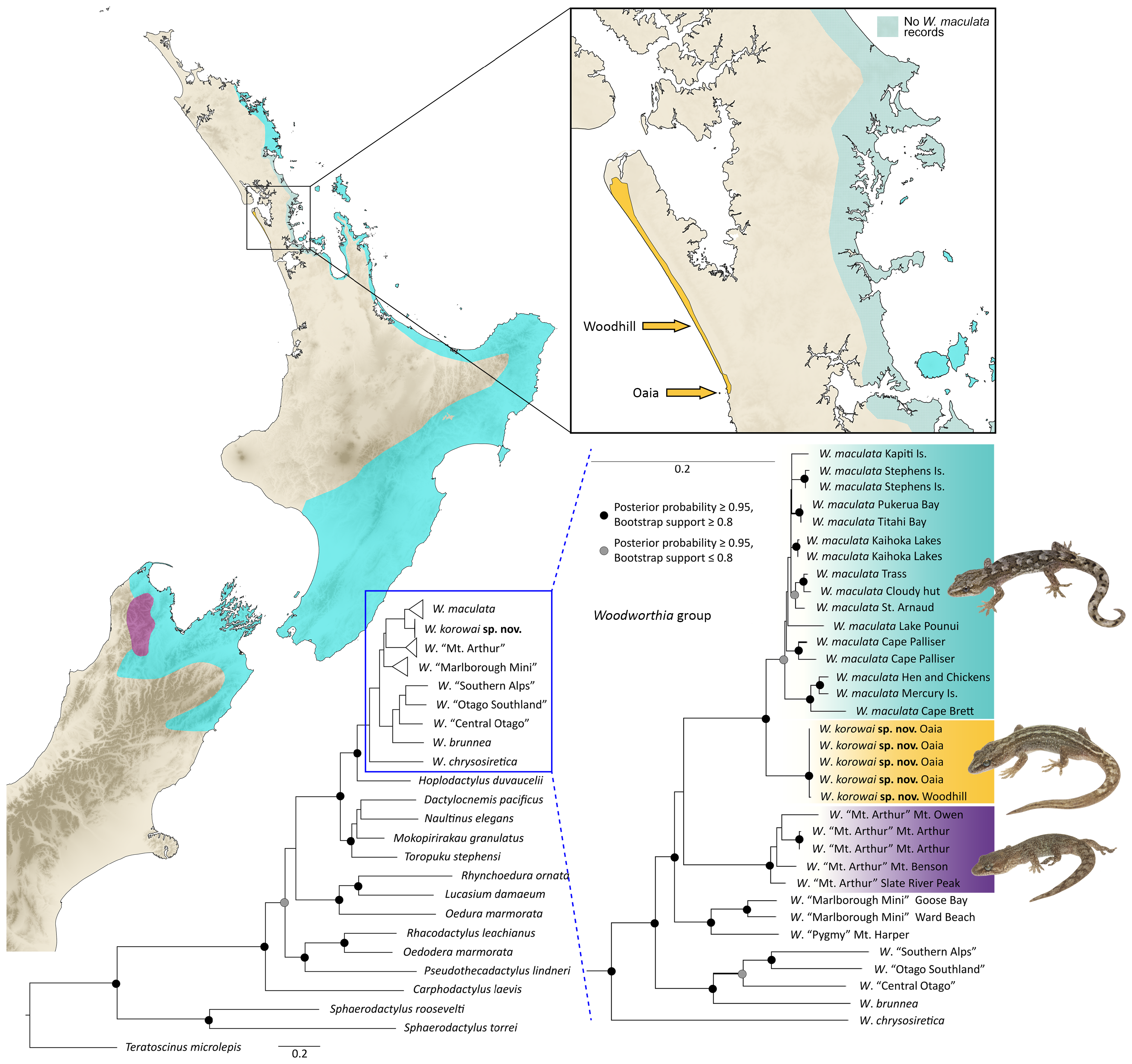



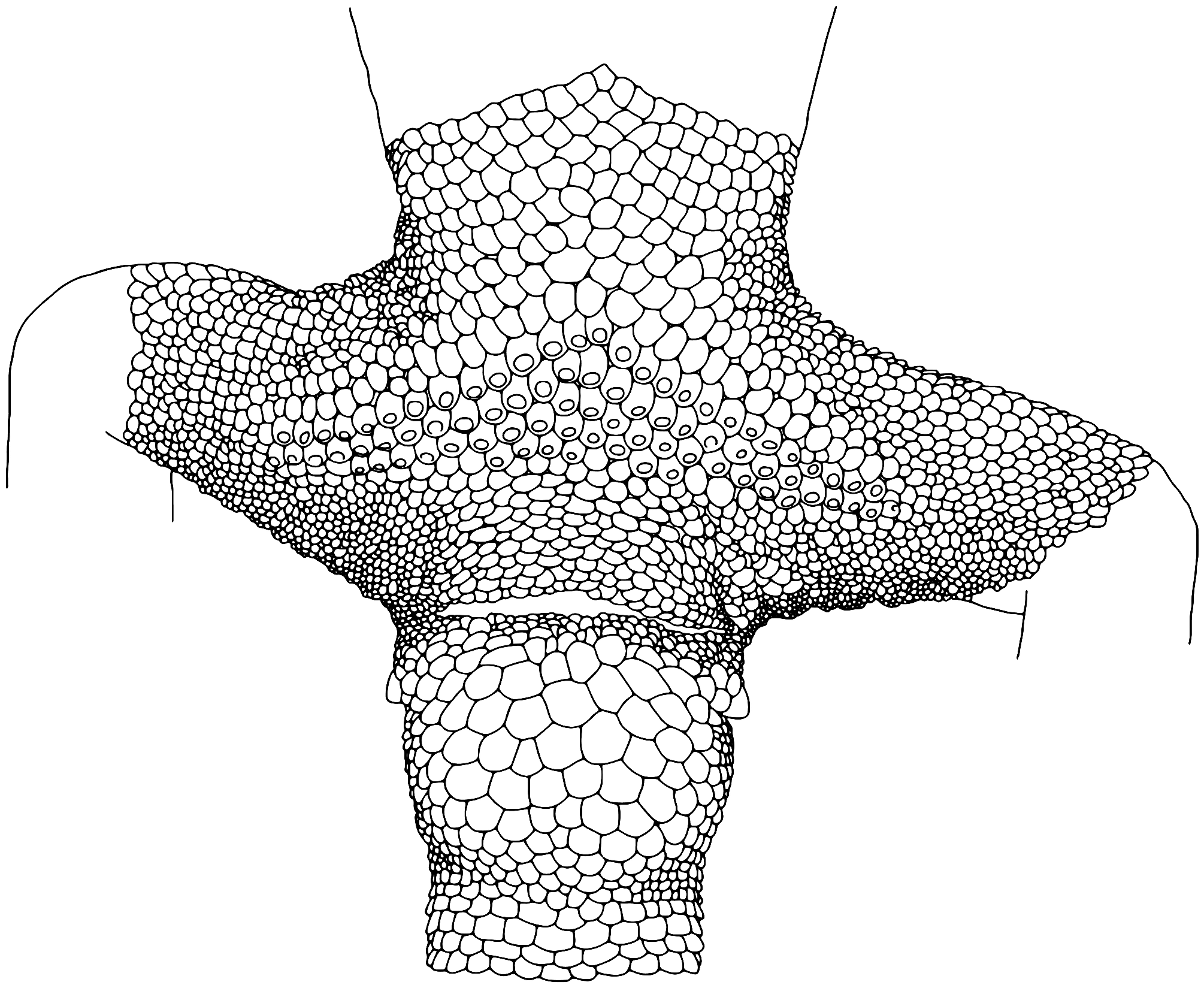
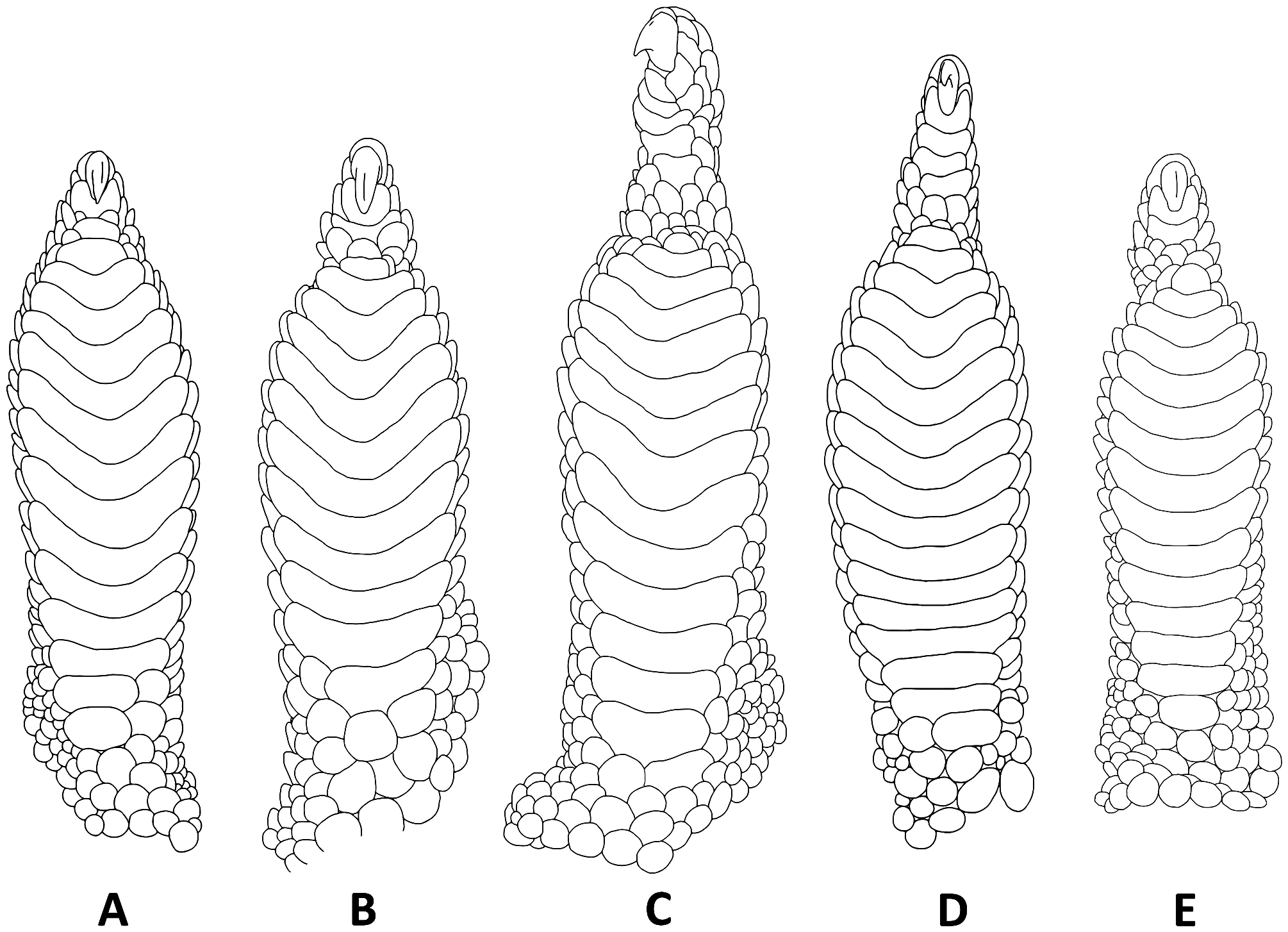

TABLE 6. Meristic, mensural (mm), proportional value, and character data from the type series of W. korowai sp. nov. (n = 4). Data (mean ± standard deviation, range in parentheses below) for all examined W. korowai sp. nov. specimens (type series and two additional in-life examined specimens) and geographically close and genetically closely related congeners (W. maculata, n = 26 and W. “Mt Arthur”, n = 4). Abbreviations are listed in the Materials and methods. Key: ND = no data, na = not applicable. Individuals with regenerated tails were excluded from pooled (mean) data for TL, TW, and TH characters. Bold text indicates characters with statistical significance among the three taxa.
| Woodworthia korowai sp. nov. | W. maculata (n = 26) | W. “Mt Arthur” (n = 4) | |||||
|---|---|---|---|---|---|---|---|
| Holotype | Paratypes | All examined W. korowai sp. nov. (n = 6, including in- life specimens) | |||||
| AWMM LH2445 | AWMM LH4069 | AWMM LH4067 | AWMM LH4068 | ||||
| Sex | f | m | f | m | |||
| Mensural | |||||||
| SVL | 57.4 | 55.8 | 47.0 | 68.0 | 58.7 (± 6.52) (47.0–68.0) | 62.1 (± 6.67) (48.8–74.0) | 52.0 (± 4.38) (46.6–57.5) |
| TL | 66.4 | 54.8 | 49.0 | 61.0 | 57.7 (± 8.7) (49.0–66.4) | 71.1 (± 6.36) (60.5–81.0) | ND |
| TW | 6.5 | 8.0 | 4.4 | 4.4 | 5.45 (± 1.05) (4.4–8.0) | 7.1 (± 0.98) (5.3–8.3) | 7.2 (± 0.00) (7.2) |
| TH | 5.9 | 6.4 | 4.2 | 4.6 | 5.05 (± 0.85) (4.6–6.0) | 6.2 (± 0.88) (4.7–7.3) | 6.4 (± 0.00) (6.4) |
| Regen. tail | No | Yes | No | Yes | - | - | - |
| BH | 11.2 | 6.5 | ND | 5.4 | 7.8 (± 1.97) (5.4–11.2) | 9.4 (± 1.77) (6.6–12.6) | 9.7 (± 1.37) (8.3–11.8) |
| BW | 14.8 | 13.4 | 10.1 | 10.5 | 12.1 (± 1.63) (10.1–14.8) | 13.2 (± 2.10) (10.0–18.2) | 13.2(± 0.96) (12.1–14.7) |
| HL | 16.2 | 17.9 | 15.8 | 18.9 | 17.2 (± 1.04) (15.8–18.9) | 17.2 (± 1.68) (14.2–20.4) | 15.3 (± 0.69) (14.5–16.3) |
| HW | 11.4 | 11.1 | 9.4 | 12.3 | 11.4 (± 1.08) (9.4–12.8) | 12.4 (± 1.25) (10.0–14.8) | 10.8 (± 0.58) (9.9–11.4) |
| HD | 8.1 | 8.7 | 5.9 | 5.5 | 7.2 (± 1.16) (5.5–8.7) | 8.2 (± 0.81) (6.5–9.4) | 7.9(± 0.62) (7.3–8.7) |
| EN | 4.7 | 4.3 | 4.0 | 4.4 | 4.5 (± 0.3) (4.0–4.9) | 4.6 (± 0.47) (3.6–5.4) | 4.1 (± 0.28) (3.7–4.4) |
| SNT | 6.3 | 6.5 | 5.7 | 6.3 | 6.2 (± 0.26) (5.7–6.5) | 6.5 (± 0.70) (5.2–7.7) | 5.7 (± 0.29) (5.3–6.1) |
| EE | 5.6 | 6.0 | 4.6 | 5.4 | 5.5 (± 0.47) (4.6–6.0) | 5.6 (± 0.53) (4.3–6.5) | 5.2 (± 0.43) (4.6–5.7) |
| ED | 3.2 | 3.3 | 2.5 | 3.8 | 3.2 (± 0.38) (2.5–3.8) | 3.6 (± 0.51) (2.5–4.5) | 2.9(± 0.24) (2.6–3.2) |
| EW | 1.4 | 1.17L | 1.5 | 0.9 | 1.2 (± 0.28) (0.7–1.5) | 0.9 (± 0.29) (0.4–1.5) | 0.9 (± 0.12) (0.7–1.1) |
| EL | 2.0 | 1.34L | 1.1 | 2.5 | 2.0 (± 0.57) (1.1–2.6) | 1.9 (± 0.44) (0.9–2.9) | 1.6 (± 0.12) (1.5–1.8) |
| IN | 2.8 | 2.4 | 2.3 | 2.5 | 2.6 (± 0.25) (2.3–3.0) | 2.8 (± 0.30) (2.2–3.4) | 2.4 (± 0.20) (2.2–2.7) |
| SF | 21.2 | 23.8 | 20.6 | 25.8 | 22.9 (± 1.82) (20.6–25.8) | 24.4 (± 2.82) (18.5–29.4) | 21.5 (± 0.51) (21.1–22.4) |
......Continued on the next page
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.