
Tityopsis sheylae, Teruel & Rodríguez-Cabrera, 2020
|
publication ID |
https://doi.org/10.5281/zenodo.4648293 |
|
publication LSID |
lsid:zoobank.org:pub:AC17B8E0-177D-48CD-8F24-878214033EAE |
|
DOI |
https://doi.org/10.5281/zenodo.4770010 |
|
persistent identifier |
https://treatment.plazi.org/id/7DD2D5D2-87BC-49B5-A268-D6C2FB94746E |
|
taxon LSID |
lsid:zoobank.org:act:7DD2D5D2-87BC-49B5-A268-D6C2FB94746E |
|
treatment provided by |
Carolina |
|
scientific name |
Tityopsis sheylae |
| status |
sp. nov. |
Tityopsis sheylae View in CoL sp. n.
( Figures 2 View Figure 2 , 23–32 View Figures 23–24 View Figures 25–26 View Figures 27–28 View Figures 29–30 View Figures 31–32 , 36 View Figure 36 , 40 View Figures 39–40 , Tables 3–4)
http: //zoobank. org/urn: lsid: zoobank. org: act: 7DD2D5D2- 87BC-49B5-A268-D6C2FB94746E
TYPE LOCALITY AND TYPE DEPOSITORY. Cuba, Mayabeque Province, border between San José de Las Lajas and Jaruco Municipalities, Tapaste, Escaleras de Jaruco, Mogote La Jaula, Cueva del Indio, 190 m a. s. l.; RTO .
TYPE SPECIMENS. Cuba, Mayabeque Province, border between San José de Las Lajas and Jaruco Municipalities, Tapaste , Escaleras de Jaruco , Mogote La Jaula , Cueva del Indio , 190 m a. s. l., 3 April 2019, leg. R. Teruel, S. Yong & T. M. Rodríguez, 1♂ ( holotype), RTO, 13 December 2014, leg. T. M. Rodríguez & A. Longueira, 3♂ 7♀ 2♂ juvs. 3♀ juvs. ( paratypes), RTO, 12 May 2017, leg. S. Yong & L. Forcelledo, 1♀1♀ juv. ( paratypes), RTO, 5 January 2018, leg. R. Teruel, S. Yong & L. Forcelledo, 5♂ 9♀ 2♂ juvs. 3♀ juvs. ( paratypes), RTO, 3 April 2019, leg. R. Teruel, S. Yong & T. M. Rodríguez, 2♂ 6♀ ( paratypes), RTO .
OTHER MATERIAL EXAMINED. Cuba, Mayabeque Province, border between San José de Las Lajas and Jaruco Municipalities, Tapaste, Escaleras de Jaruco, La Jaula, Cueva del Indio, 190 m a. s. l., 23 December 1995, leg. A. Pérez, 1♂ (topotype), IES, 27 September 1999, leg. E. Fonseca, 1♀ (topotype), IES .
Note. Both specimens were repeatedly seen by one of us (RT) between 2005 and 2011, but are currently missing from IES collection. As their whereabouts is unknown (there is no official record of a loan, destruction or loss in the collection log), both were excluded from the type series.
ETYMOLOGY. The selected epithet is a Latinized matronym honoring the senior author’s wife Sheyla Yong, who enthusiastically assisted us all along the present revision, both in the field (she actually collected several of the type specimens) and in the lab (part of the data gathering and processing, as well as photographic work).
DIAGNOSIS. Adult size moderately large for the genus ( 24– 27 mm in males, 32–34 mm in females). Adult coloration: base light yellowish brown, slightly darker on metasomal segments IV–V and slightly paler on legs, venter and telson; dark patterns essentially absent, except for faint infuscation in carapace (mostly around eyes) and metasomal segments II–IV (in some specimens only, mostly females). Juvenile coloration: base pale yellowish brown, paler on pedipalp chelae, legs, venter and telson; dark patterns essentially absent, except for diffuse gray to dark brown reticulation in chelicerae, carapace (along anterior margin and around eyes) and metasomal segments II–V (mostly on posterior part and around carinae); tergites with posterior margins translucent whitish; pedipalps fingers infuscate. Pedipalps remarkably elongate and robust for the genus (length/width ratio of femur, patella and chela: 2.94–3.11, 2.58–2.65 and 3.50–3.66 in males, 2.79–2.96, 2.64–2.71 and 3.51–3.57 in females); chelae with manus incrassate oval and very slightly longer in males (length/width ratio: 1.50–1.56 in males, 1.46–1.53 in females) and with fingers very long (movable finger/manus length ratio: 1.33–1.34 in males, 1.33–1.40 in females), fixed/movable fingers with 12–13/12–13 principal rows of denticles (mode 12/12). Tegument of carapace, tergites, sternites, metasoma and telson sexually dimorphic between adults: matt in male vs. with a satin sheen in females. Carapace and tergites with many medium-sized to coarse granules scattered (similar in both sexes). Pectines with 13–16 teeth in males, 13–15 in females (mode 14 and 13–14, respectively); basal middle lamella slightly enlarged and teardrop-shaped in males, greatly enlarged and widely oval in females. Sternite V with median smooth patch widely cordiform and about as long as wide in both sexes, very large in males and moderately large in females; lateral smooth patches oval, oblique and bulky in both sexes, moderately large in males and small in females. Metasoma slightly elongate in both sexes (length/width ratio of segments I–V: 1.12–1.16, 1.42–1.44, 1.54–1.58, 1.76–1.78 and 2.22–2.25 in males vs. 1.14–1.15, 1.43–1.45, 1.54–1.64, 1.70–1.71 and 2.22–2.24 in females), with 10/10/8/8/7 complete to almost complete, coarsely serrate to serrato-crenulate carinae; lateral inframedians on II obsolete only on basal one-sixth (males) to one-fifth (females); intercarinal tegument of segment V with many small (males) to medium-sized (females) conical granules. Telson with vesicle less globose than standard for the genus, subaculear tubercle small.
DESCRIPTION (adult male holotype; Figs. 2 View Figure 2 , 23 View Figures 23–24 , 25 View Figures 25–26 , 27 View Figures 27–28 , 29 View Figures 29–30 , 31 View Figures 31–32 , 36a View Figure 36 ; Tabs. 3–4).
Coloration ( Figs. 23 View Figures 23–24 , 36a View Figure 36 ) base light yellowish brown, slightly darker on metasomal segments IV–V (medium brown) and slightly paler on legs and venter. Chelicerae manus very sparsely reticulate with brown (more densely distally); fingers very faintly and irregularly infuscate. Pedipalps immaculate, with carinae and fingers slightly darker due to heavier sclerotization; fingers infuscate. Carapace largely immaculate, with faint symmetrical infuscation around all three eye groups; eyes and ocular tubercles black. Tergites immaculate, with carinae of VII slightly darker due to heavier sclerotization. Pectines immaculate pale yellowish brown, with basal portion and basal plate progressively darker due to heavier sclerotization. Sternites immaculate; V with median and lateral smooth patches bright white. Legs immaculate, with carinae slightly darker due to heavier sclerotization. Metasoma with segments IV–V slightly darker (medium brown), with carinae darker due to heavier sclerotization and without any annulated pattern; all surfaces immaculate. Telson immaculate, with distal half of aculeus dark reddish brown.
Chelicerae ( Fig. 27a View Figures 27–28 ). With dentition typical for the genus, teeth standard-sized and sharp. Tegument glossy but with minute granulation scattered, dorsodistal portion of manus with coarse, glossy granules irregularly arranged transversally around dorsodistal depression. Setation very dense ventrally, but essentially lacking dorsally, except for five rigid macrosetae around dorsodistal depression of manus.
Pedipalps ( Fig. 25a–c View Figures 25–26 ). Remarkably enlarged for the genus, i.e., all segments longer and chelae also heavier than standard. Femur essentially straight (very slightly bent inwards distally), with all carinae very strong, coarsely denticulate to serrate; intercarinal tegument matt, very densely and irregularly granulose; internal ( i) trichobothria surrounding a large, irregularly triangular spur. Patella straight, with all carinae very strong, coarsely granulose to serrato-crenulate; intercarinal tegument matt, very finely and densely granulose, internally with many conical denticles. Chela with manus incrassate oval (1.56 times longer than wide), much wider than patella (ratio 1.26) and with the basal half slightly widest, all carinae strong, coarsely serrate to serrato-crenulate, intercarinal tegument matt, very finely and densely granulose on all surfaces; fingers very long for the genus (movable finger 1.34 times longer than underhand) and evenly curved, fixed finger with 12/12 principal rows of denticles, movable finger with 12/12 plus an apical subrow of four denticles and a large internal accessory denticle (large terminal denticle not included), basal lobe/notch combination absent.
Carapace ( Fig. 27a View Figures 27–28 ). Wider than long (length/width ratio = 0.93). Anterior margin widely V-shaped, with two pairs of dark macrosetae. Carinae: anterior medians anteriorly divergent and formed by coarse, isolated granules and irregularly fused to the superciliaries, which are composed of smaller and much closer granules; posterior medians straight, parallel and formed by medium-sized granules; other carinae absent or indistinct from general granulation. Furrows: anterior marginal, anterior median, median ocular, lateral oculars, central transverse, central median, posterior median and posterior marginal all fused, wide and moderately deep; posterior laterals long, oblique, wide and moderately deep, almost fused to posterior marginal; posterior transverse narrow and moderately deep; other furrows absent or indistinct. Tegument matt, very densely and irregularly but finely granulose, with many medium-sized, rough granules scattered. Median eyes relatively large and separated by about one ocular diameter; lateral eyes noticeably smaller and moderately exposed beyond anterolateral margin.
Sternum ( Fig. 29a View Figures 29–30 ). Standard for the genus, although more pentagonal in shape, with three pairs of dark macrosetae. Tegument very densely and irregularly granulose.
Genital operculum ( Fig. 29a View Figures 29–30 ). Standard for the genus, with three pairs of dark macrosetae. Valves roundly subtriangular and not separated medially; tegument minutely and densely granulose. Genital papillae large, thick and not protruding.
Pectines ( Fig. 29a View Figures 29–30 ). Moderately large for the genus (but not reaching coxa-trochanter joint of leg IV). Tooth count 14/15, teeth straight and slightly swollen. Fulcra large and bulky, paraboloid to round. Basal middle lamella slightly enlarged and teardrop-shaped. Pre-pectinal plate well visible, heavily sclerotized, very wide and short; tegument coriaceous to smooth. Basal plate normally sclerotized, about as long as wide; anterior margin with a very wide, V-shaped anteromedian notch, posterior margin convex; tegument smooth.
Legs. Slender, with all carinae strong, finely denticulate to granulose. Intercarinal tegument minutely and densely granulose, with a row of slightly coarser granules defining an additional carina (externomedian) on lateral surface of femur of all legs. Claws short and strongly curved.
Mesosoma ( Figs. 27b View Figures 27–28 , 29 View Figures 29–30 ). Tergites matt, very densely and irregularly but finely granulose, with many medium-sized to coarse, rough granules scattered; carinae: median longitudinal strong, serrato-crenulate to serrate and progressively longer from I–VII, submedians absent on I–VI, complete, straight and strongly serrato-denticulate on VII, laterals absent on I–VI, complete, straight and strongly serrato-denticulate on VII. Sternites matt, very densely and irregularly but finely granulose, with some dark macrosetae scattered all over; lateral margins of all sternites subgranulose to granulose; posterior margin of all sternites smooth to vestigially granulose, on III–IV vestigially bilobed, on V bilobed (due to protuberance of lateral smooth patches), on VI essentially straight, on VII widely concave; carinae: III–V acarinate, VI-VII with paired laterals and submedians which are long, straight and finely crenulate; spiracles small, narrowly oval to short slit-like; sternite V with median smooth patch very large, widely cordiform (about as long as wide), bulky and glossy, not protruding from posterior margin; lateral smooth patches moderately large, oblique, oval, bulky and glossy, moderately protruding from posterior margin.
Metasoma ( Fig. 31 View Figures 31–32 ). Slightly longer than standard for the genus (4.23 times longer than carapace) and slightly narrower distally. Segments I–II with ten complete to almost complete carinae, III–IV with eight, V with seven: dorsal laterals strong, coarsely serrate to serrato-denticulate on I–III, coarsely crenulate-denticulate on IV, absent on V but subtly indicated by a few granules; lateral supramedians strong, coarsely serrate to serrato-crenulate on I–V; lateral inframedians strong, coarsely serrato-crenulate on I–II (obsolete on basal one-sixth of the latter), absent on III–V (but indicated in each segment by an incomplete row of granules that never form a true, raised carina); ventral laterals strong, coarsely crenulate to serrato-crenulate on I–V; ventral submedians strong, coarsely serrate to serrato-crenulate on I–IV, indicated by an incomplete but raised row of coarse granules on less than basal half of V; ventral median absent on I–IV, strong, coarsely serrato-crenulate on V. Intercarinal tegument matt, very finely and densely granulose, with many medium-sized granules scattered on dorsal surface of all segments and many small, comical granules on lateral surfaces of V; dorsal furrow absent or indistinct on segments I–III, complete, narrow and moderately deep on IV–V; setation sparse, with two pairs of dark macrosetae on essentially all carinae.
Telson ( Fig. 25d View Figures 25–26 ). Vesicle less globose than standard for the genus (1.48 times longer than wide, 1.02 times wider than deep) and with several dark and whitish setae scattered; tegument coriaceous and matt, smooth even ventrally; ventral median carina very weak but coarsely subgranulose, continued into the subaculear tubercle that is small, broadly conical and unarmed (i.e., without any granules). Aculeus standard-sized, sharp, shorter than vesicle and moderately curved.
FEMALE (adult paratopotypes; Figs. 24 View Figures 23–24 , 26 View Figures 25–26 , 28 View Figures 27–28 , 30 View Figures 29–30 , 32 View Figures 31–32 , 36c–d View Figure 36 ; Tabs. 3–4). Sexual dimorphism well-marked, differing from described male by: 1) size conspicuously larger (32.1–34.0 mm); 2) tegument of carapace, tergites, sternites, metasoma and telson with a satin sheen; 3) pedipalps relatively shorter, slightly more robust (especially chela manus) and with carinae finer; 4) carapace slightly wider posteriorly; 5) mesosoma relatively wider, with lateral sides markedly convex; 6) pre-pectinal plate absent; 7) genital papillae absent; 8) pectines relatively smaller, with teeth shorter, narrower and slightly fewer in number (13–15, mode 13–14), and with basal middle lamella greatly enlarged, widely oval in shape and whitish; 9) sternite V with median and lateral smooth patches much smaller, less bulky and translucent; 10) metasoma with carinae slightly stronger; 11) telson vesicle with many medium-sized granules scattered ventrally and ventrolaterally, and ventral median carina stronger.
VARIATION.The 44 paratopotypes are remarkably homogeneous for most morphological characters such as coloration, tegument sculpture, carination, setation and morphometric ratios, where variation was only minor and mostly due to eval differences ( Fig. 36 View Figure 36 ), e. g., older adults have slightly darker coloration and setation, granulation and claws worn-out to different degrees. The only noteworthy exceptions are discussed as follows.
Adult size ( Tab. 3) varied from 23.6–26.8 mm in males and 32.1–34.0 mm in females. There are two size classes even distinguishable to unaided eye in adult males (separated by a maximum gap of 3.14 mm, i.e., a 12% size difference), but only one in females (5% maximum size difference).
Number of principal rows of denticles varied from 12–13 on both fixed and movable fingers, with a clearly defined mode of 12 in each case. Fixed, symmetrical counts for both pedipalps (i.e., same number of rows in all four fingers of the same individual), were the dominant occurrence by far: of 35 specimens analyzed (10 paratopotypes excluded due to having at least one finger broken or too damaged for reliable counting), 25 had 12/12 rows (including the holotype) and one had 13/13 rows (an adult female). The cases of asymmetry did not show any bias towards a defined finger, i.e., they occur in similar frequencies on fixed and movable fingers, and on left and right pedipalps.
Pectinal tooth counts ( Tab. 4 View Table 4 ) varied from 13–16 in males and 13–15 in females, with clearly defined modes of 14 and 13– 14, respectively. Single-tooth asymmetry was a very common occurrence: 7 out of 15 males and 11 out of 25 females (two adults of this sex had a pecten either missing or too damaged for counting); there were no specimens with two or more teeth asymmetry.
AFFINITIES. By adults exhibiting the unusual combination of a size large for the genus (over 23 mm in males and 32 mm in females) with a light yellowish, essentially immaculate coloration, this species resembles only T. inaequalis . All other congeners (either described or not) are combined conversely, i.e., if coloration is pale, then size is small and vice versa (actually, most of them are medium-sized and reddish-colored with well-defined darker patterns).
Adults of T. inaequalis can be easily separated from those of T. sheylae sp. n. by the following characters: 1) size slightly but consistently larger ( 26–29 mm in males, 33–36 mm in females); 2) female pedipalp chela slightly shorter and more robust (length/width ratio: 3.43–3.51 in females); 3) female pectines with basal middle lamella slightly smaller and more oval in shape; 4) male sternite V with median smooth patch narrower, longer than wide and lanceolate in shape.
Besides, both species are widely allopatric in isolated orographic systems: T. inaequalis in Sierra de Los Órganos ( Pinar del Río Province) and T. sheylae sp. n. in Escaleras de Jaruco ( Mayabeque Province), with their nearest occurrences separated by 170 km air-distance.
ECOLOGICAL NOTES. T. sheylae sp. n. has been found so far only inside a single cave ( Fig. 40 View Figures 39–40 ). It occurs scattered all along it, from the first chamber right beyond the entrance (not completely dark, relatively cold, less humid and essentially devoid of bat guano), through the deepest corridors (in absolute darkness, very hot, with water-saturated air and densely covered by bat guano).
We always used the UV-detection technique to find it and were able to notice that apparently because of the different microclimatic conditions, the scorpions changed their behavior accordingly: those from the first chamber were mostly found hidden under rocks (only one adult of each sex was found walking between the rocks, in separate collecting events), while all those living in the deep were totally or partially exposed on the walls and floor, either walking slowly or sit-and-wait hunting.
It lives syntopically with the buthid Centruroides gracilis (Latreille, 1804) , but one species always outnumbers the other depending on the cave site. In the hottest chamber T. sheylae sp. n. is rare (only three specimens found so far), while hundreds of C. gracilis virtually cover all available surfaces. Conversely, in the colder chambers and corridors we have found less than 15 individuals of C. gracilis in total, against 42 of T. sheylae sp. n. Interestingly, no cases of mutual predation have been observed yet, in spite of the coexistence and abundance of both scorpion species all along the cave.
On 3 April 2019, we found a small-sized adult male that was being predated upon by an adult female of the spider Ciboneya nuriae Huber & Pérez, 2001 (Pholcidae) . The scorpion had just molted (its exoskeleton was still soft and not completely stretched yet, see Fig. 36f View Figure 36 ) and was already dead about 1 cm away from its exuvium, while the spider sucked it somewhere between the chelicerae and pedipalp coxae. We deduce the scorpion anchored to molt beside the web and once it came out of the exuvium, it became entangled and was then killed by the spider.
According to its collecting label, the male collected on 1995 (see above in Other Material Examined section) was also found dead, entangled in a spider’s web. Unfortunately, this sample is currently missing from its repository and the identity of the spider cannot be established, but this second finding evidences that spiders are a routine predator of this scorpion.
COMMENTS. The habitat outside the cave is mesophylous semicaducifolious forest on limestone karstic soil ( Fig. 40a View Figures 39–40 ), typical for most members of the genus. Nevertheless, repeated intensive searches conducted by us and some collaborators there and in other mogotes of this plateau, never yielded any evidences of an epigean occurrence of T. sheylae sp. n. (e.g., live specimens or remains of carcasses and exuvia). Only two females tentatively referable to T. inexpectata were found, one of them from IES collection and which vouchers the Escaleras de Jaruco record by Armas (1974: 5). Thus, we classify T. sheylae sp. n. herein as a true troglobite.
DISTRIBUTION ( Fig. 2 View Figure 2 ). This species is apparently a troglobite, known only from the type locality. Cueva del Indio is located in the southern mogotes of Escaleras de Jaruco, which is a medium-sized (15 x 5 km), roughly east-west oriented, karstic limestone plateau, isolated in the middle of the Habana-Matanzas Plain.
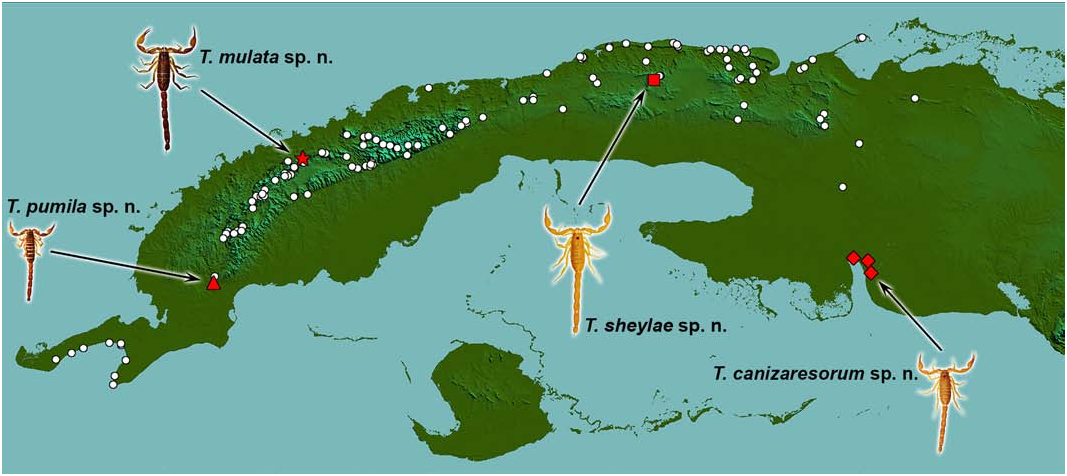

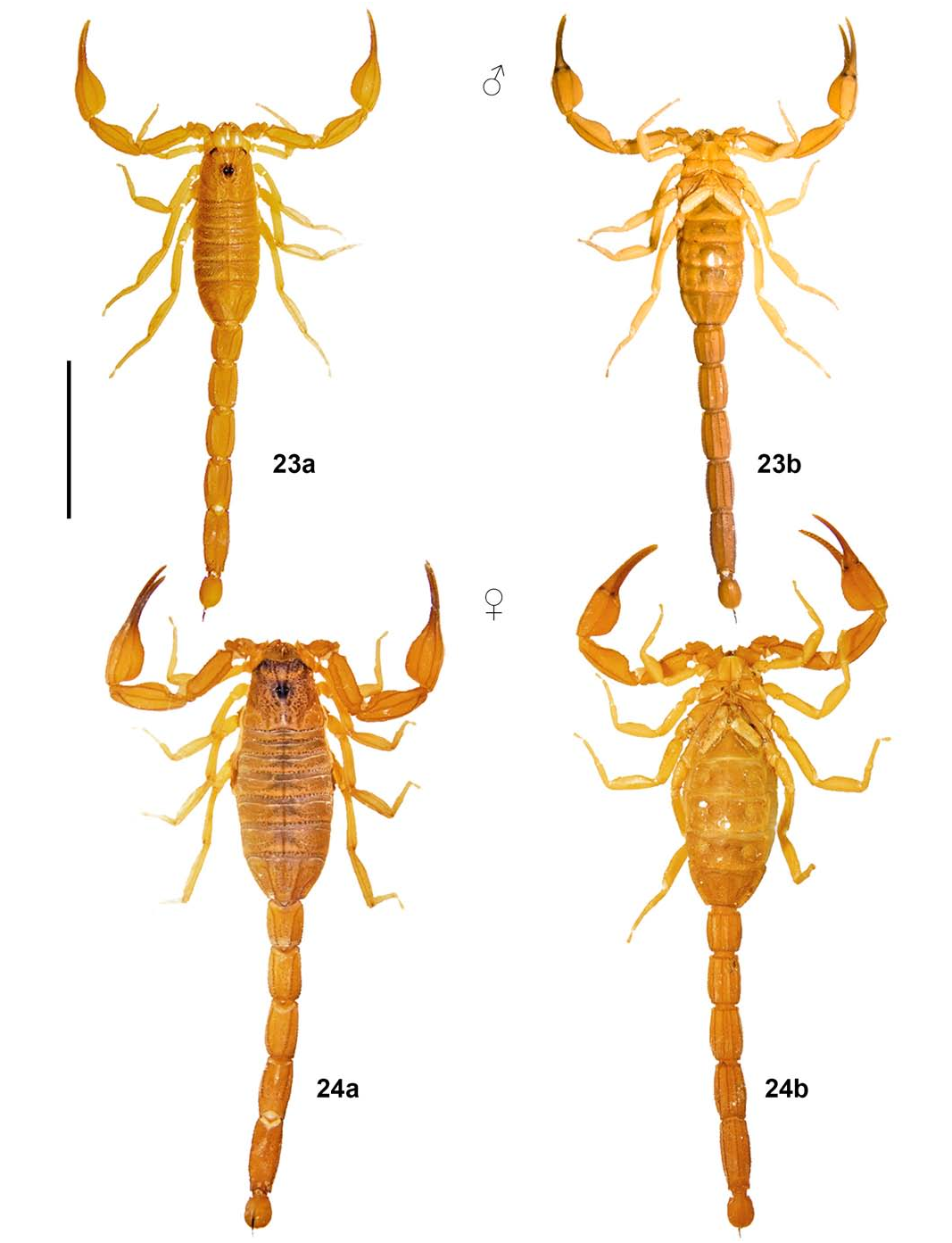
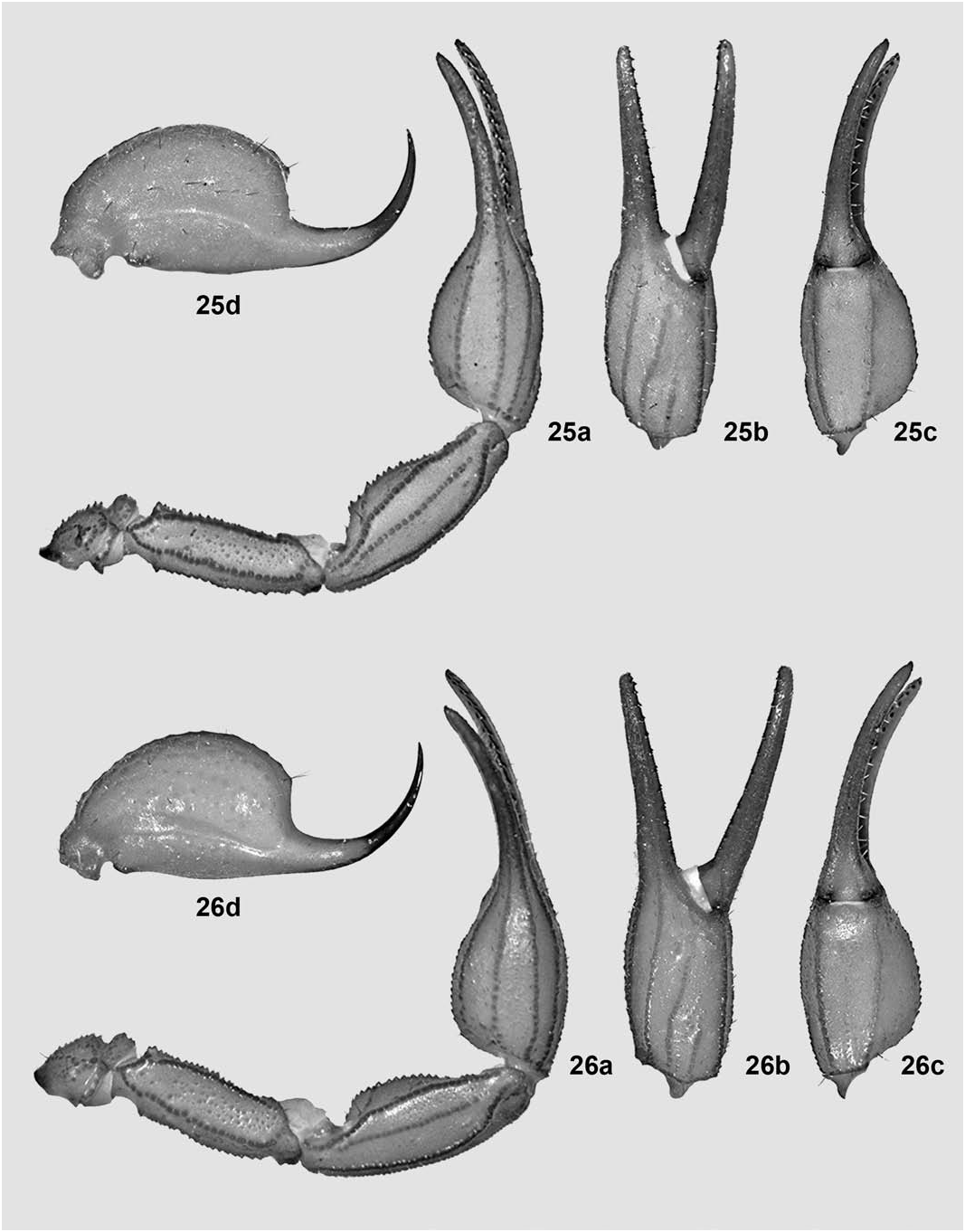
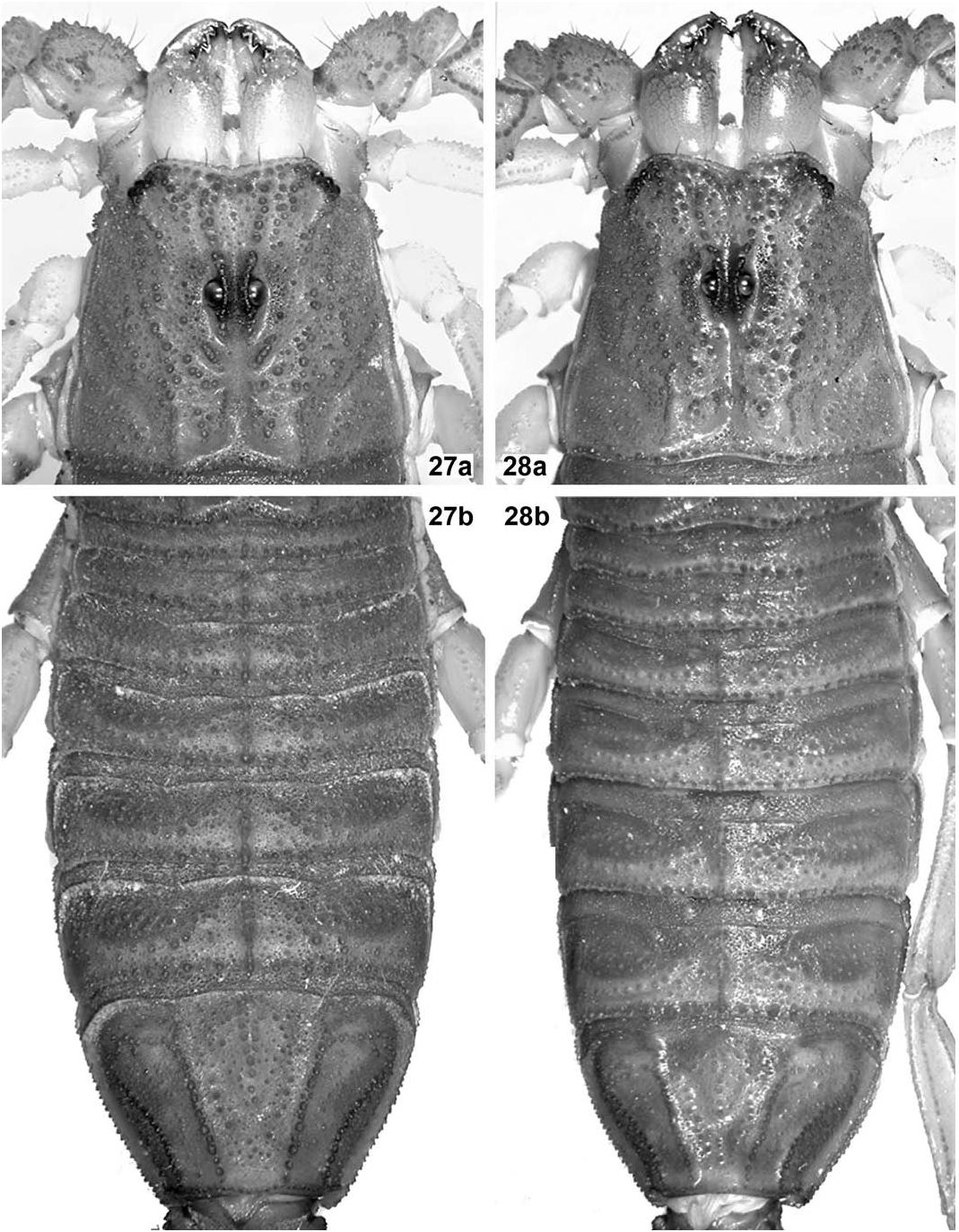
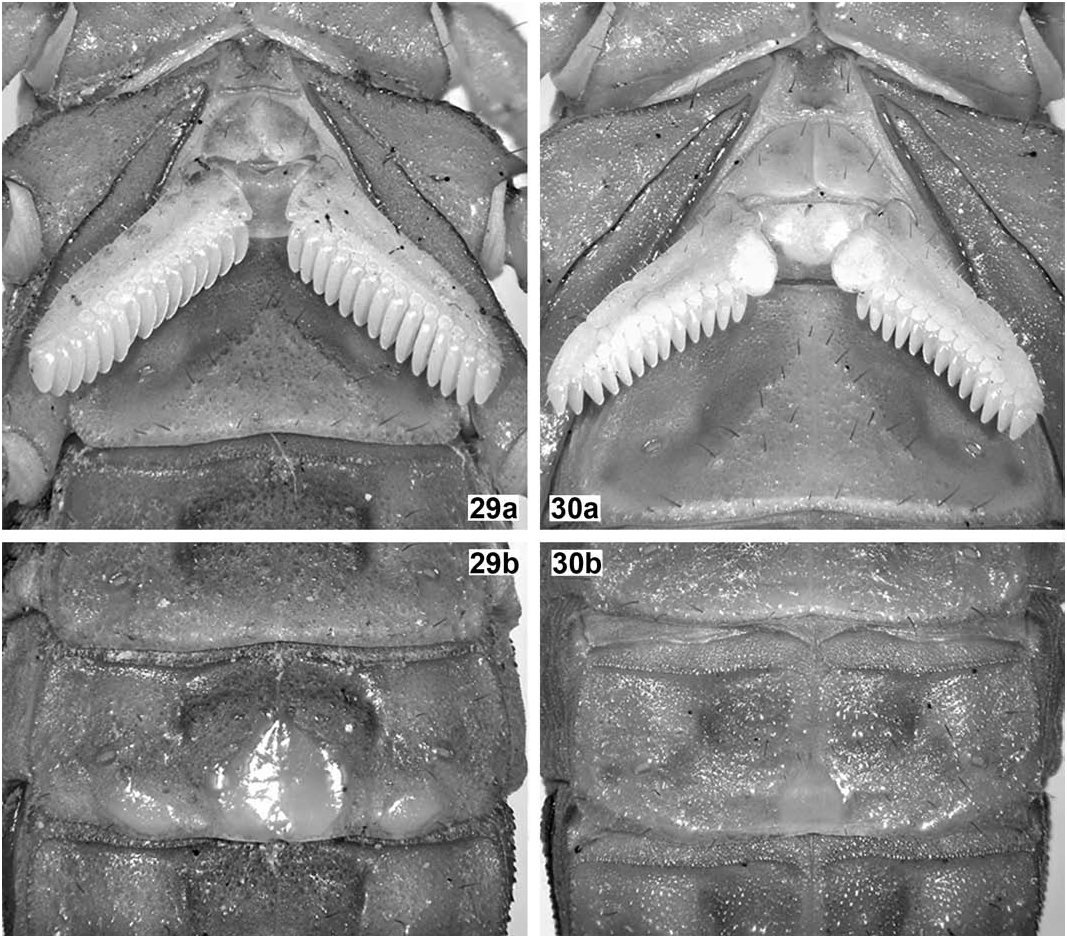
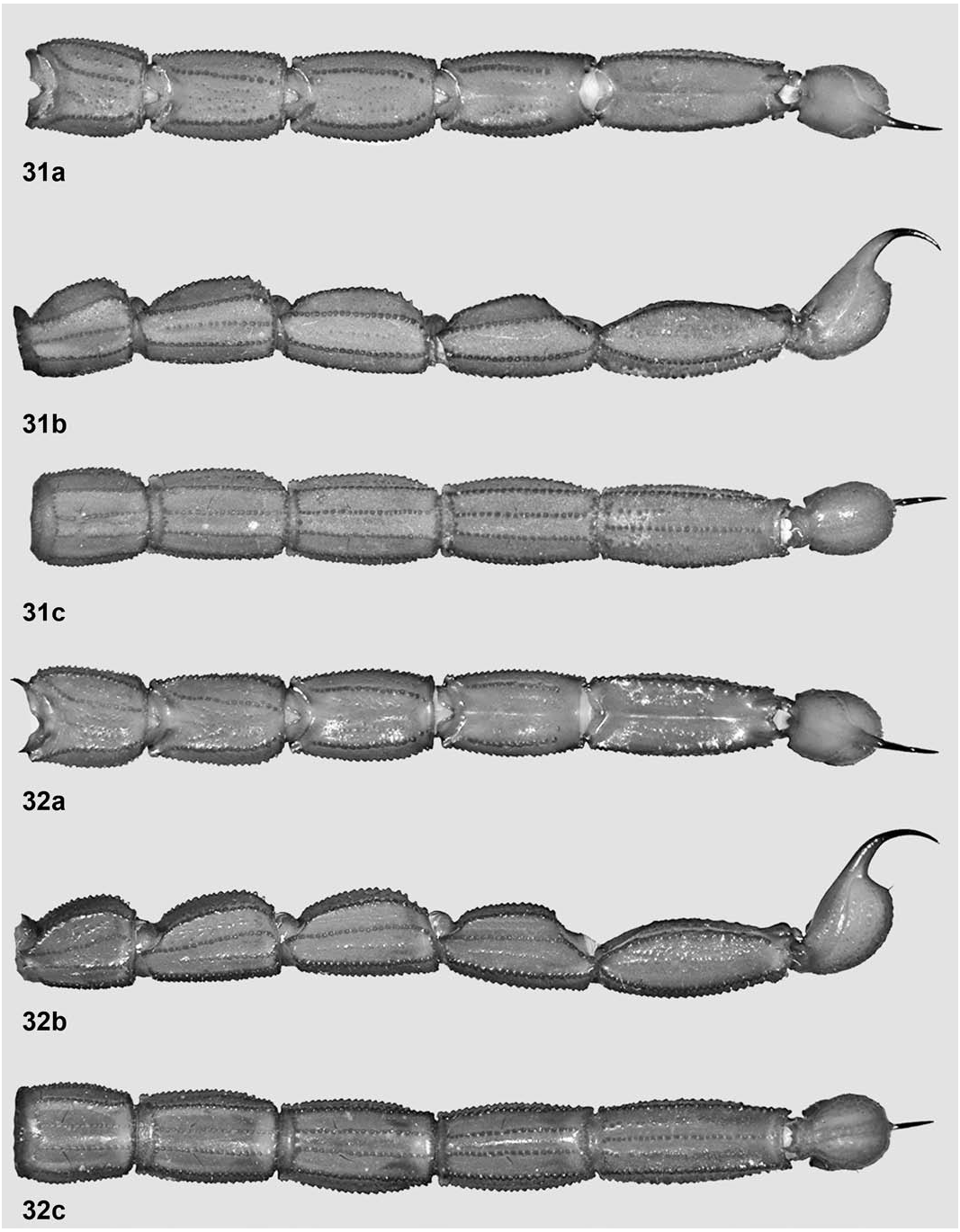

Table 4. Pectinal tooth count variation in the four species of the genus Tityopsis treated in this paper. Abbreviations: number of pectines (N),
| Species | Sex | N | 11 | Pectinal tooth count 12 13 14 15 | 16 | Ave. | SD | |||
|---|---|---|---|---|---|---|---|---|---|---|
| Tityopsis mulata sp. n. | ♂ ♀ | 8 12 | 1 | 3 | 3 5 | 4 3 | 1 | 14.75 13.83 | ± 0.66 ± 0.90 | |
| Tityopsis pumila sp. n. | ♂ | 2 | 2 | 14.00 | ± 0.00 | |||||
| Tityopsis canizaresorum sp. n. | ♀ | 76 | 2 | 33 | 39 | 2 | 12.54 | ± 0.59 | ||
| Tityopsis sheylae sp. n. | ♂ ♀ | 30 52 | 2 27 | 21 24 | 5 1 | 2 | 14.23 13.50 | ± 0.67 ± 0.54 | ||
| R |
Departamento de Geologia, Universidad de Chile |
| T |
Tavera, Department of Geology and Geophysics |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |