
Tityopsis canizaresorum, Teruel & Rodríguez-Cabrera, 2020
|
publication ID |
https://doi.org/ 10.5281/zenodo.4648293 |
|
publication LSID |
lsid:zoobank.org:pub:AC17B8E0-177D-48CD-8F24-878214033EAE |
|
DOI |
https://doi.org/10.5281/zenodo.4770006 |
|
persistent identifier |
https://treatment.plazi.org/id/AC88BB73-DE45-4045-890D-74C4D6E2A98B |
|
taxon LSID |
lsid:zoobank.org:act:AC88BB73-DE45-4045-890D-74C4D6E2A98B |
|
treatment provided by |
Carolina |
|
scientific name |
Tityopsis canizaresorum |
| status |
sp. nov. |
Tityopsis canizaresorum View in CoL sp. n.
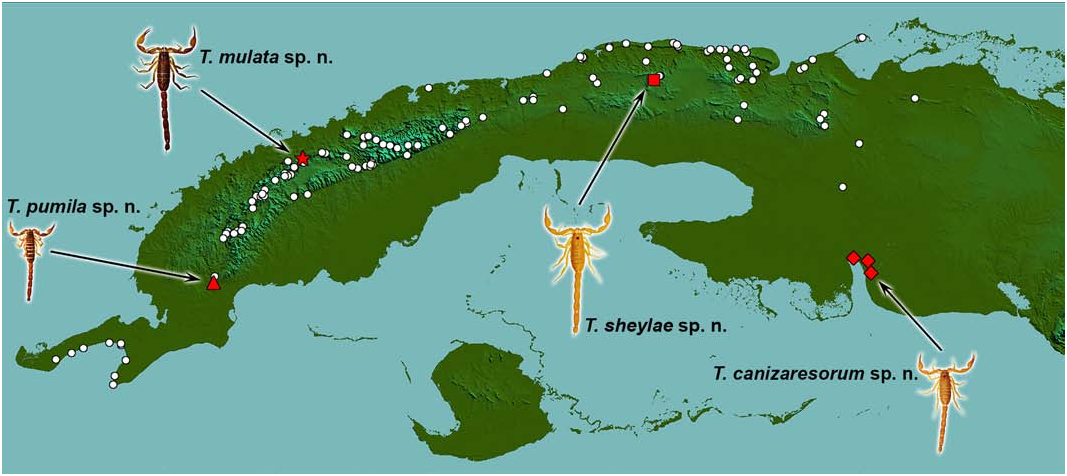
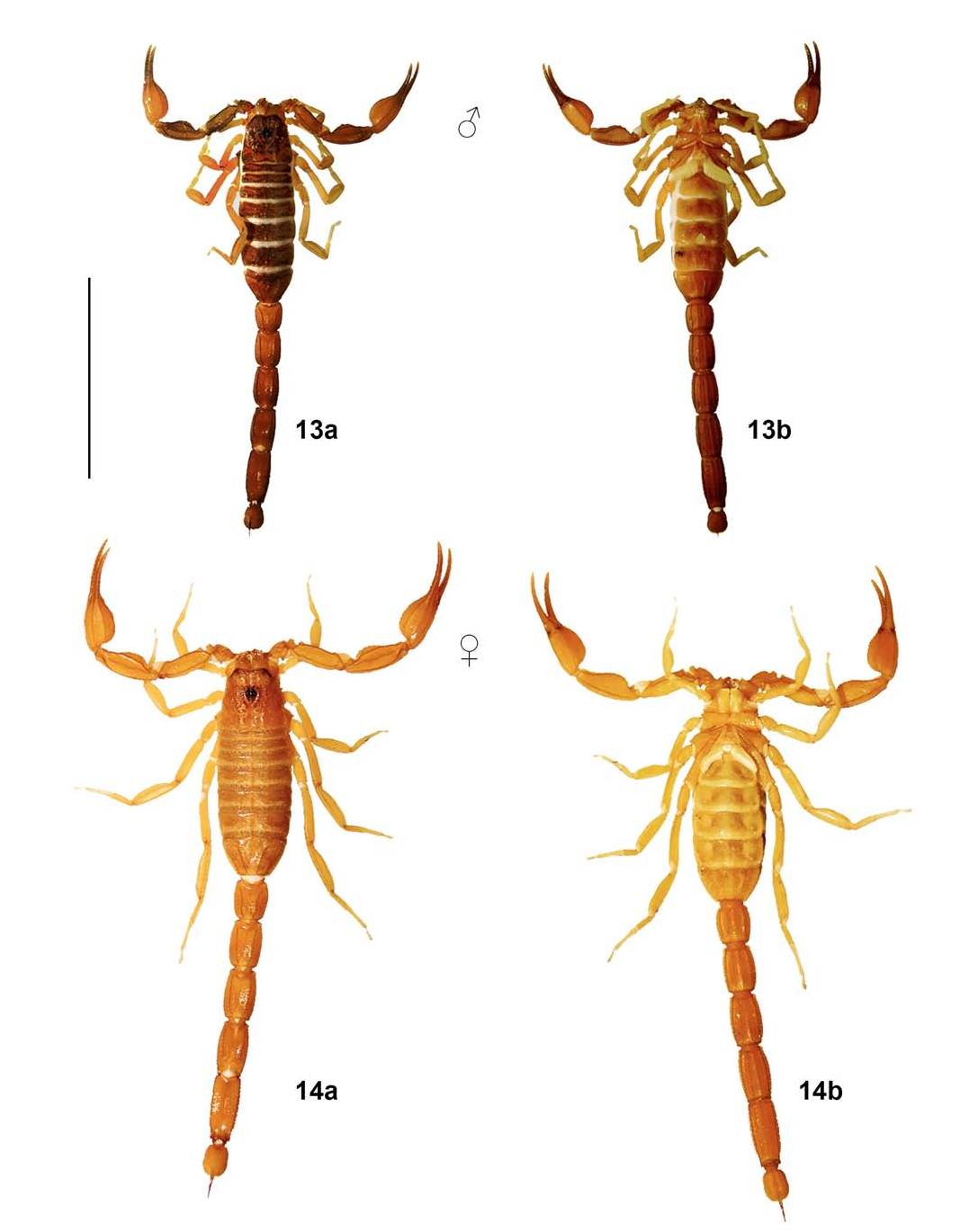
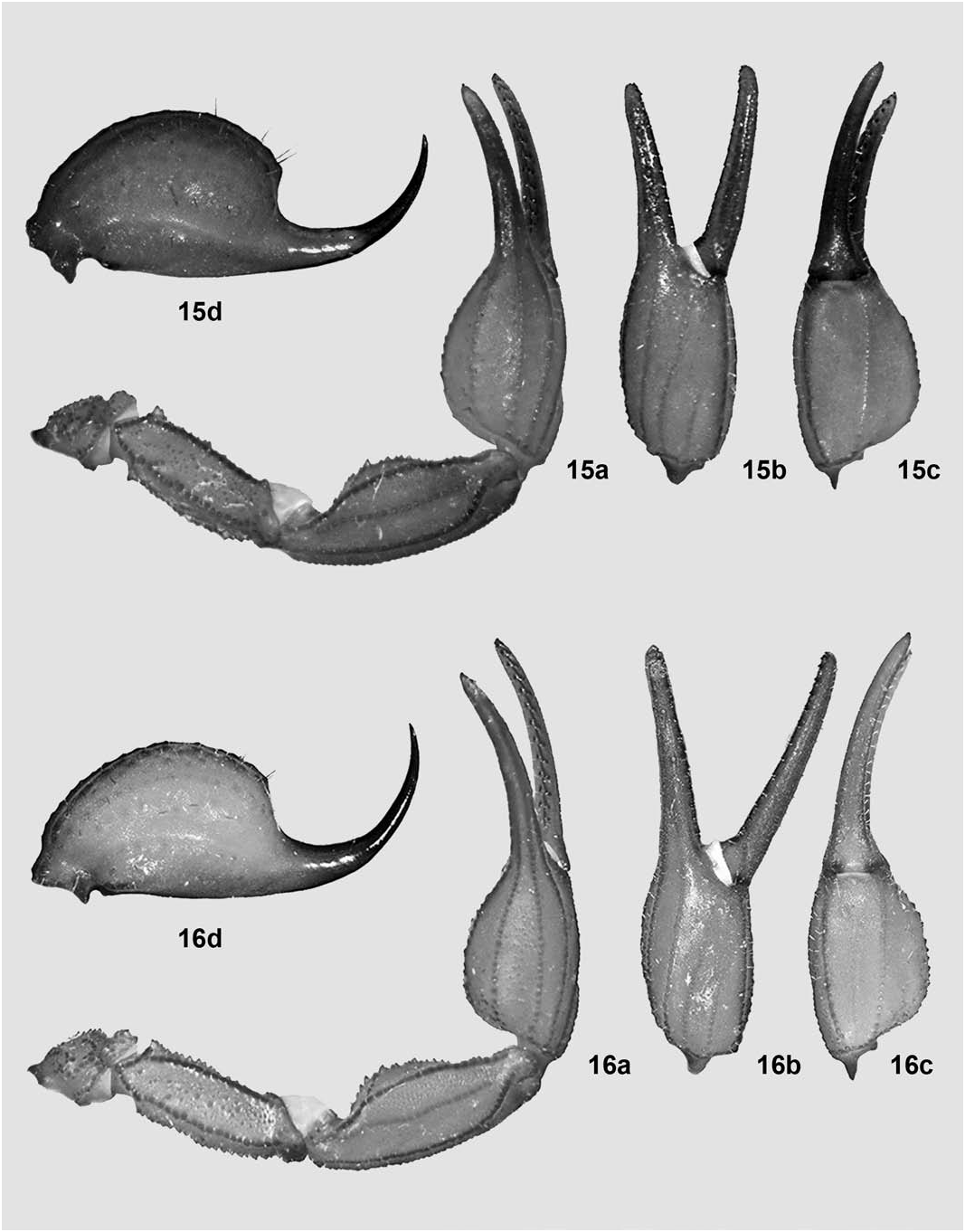
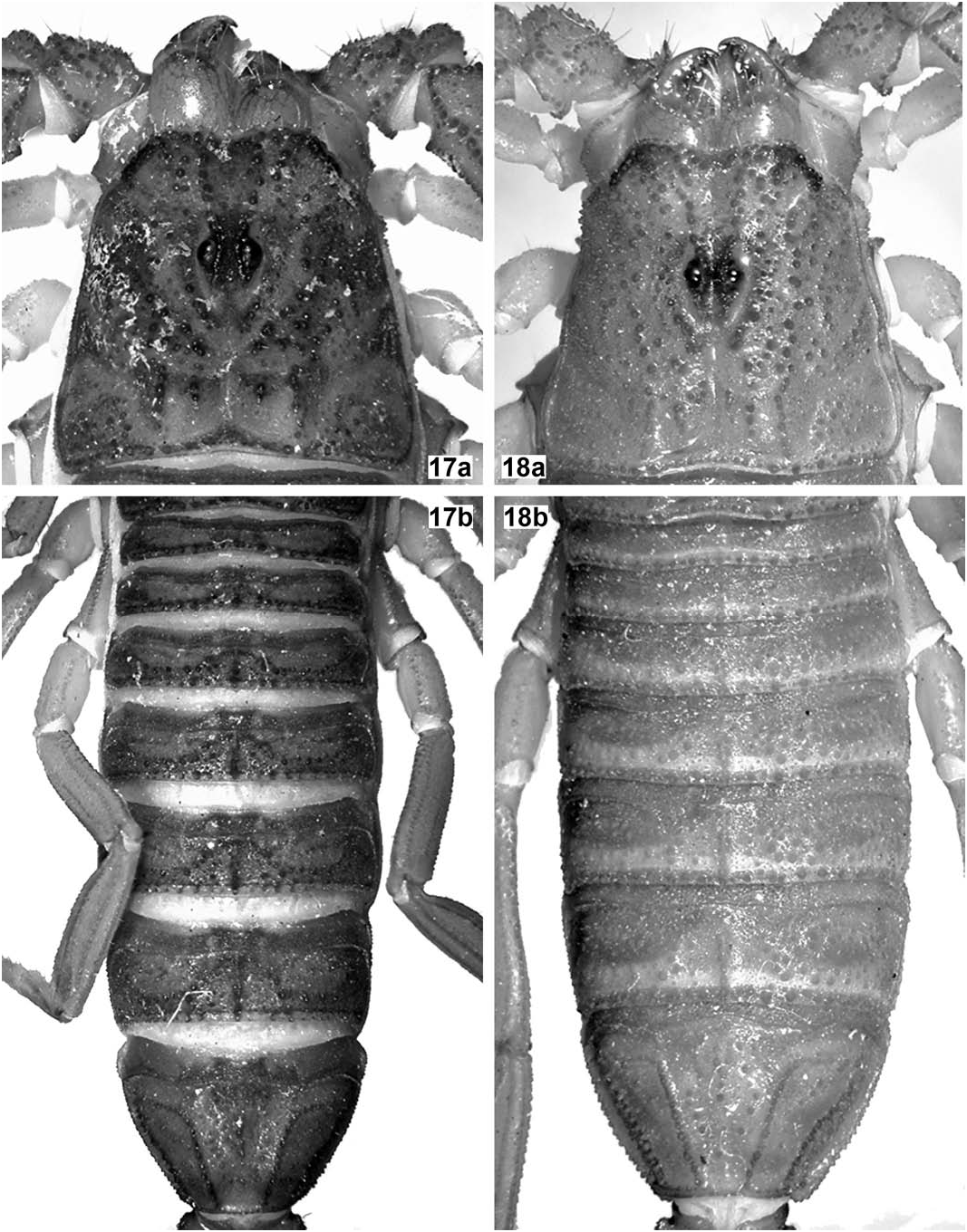
( Figures 2 View Figure 2 , 14 View Figures 13– 14 , 16 View Figures 15–16 , 18 View Figures 17–18 , 20 View Figures 19–20 , 22 View Figures 21–22 , 35 View Figures 34–35 , 39 View Figures 39–40 , Tables 2, 4 View Table 4 )
http: //zoobank. org/urn: lsid: zoobank. org: act: AC88BB73- DE45-4045-890D-74C4D6E2A98B
TYPE LOCALITY AND TYPE DEPOSITORY. Cuba, Matanzas Province, Ciénaga de Zapata Municipality, Los Hondones , 2–3 m a. s. l., RTO .
TYPE SPECIMENS. Cuba, Matanzas Province, Ciénaga de Zapata Municipality, Los Hondones , 2–3 m a. s. l., 22–25/ January/2013, leg. R. Teruel & T. M. Rodríguez, 1♀ (holotype), RTO; Matanzas Province, Ciénaga de Zapata Municipality, Playa Larga , 1 m a. s. l., 20 June 2011, leg. L. A. Casanella, F. Cala, 1♀ juv. (paratype), RTO; 3.5 km southeast of Soplillar , 3 m a. s. l., 25 January 2013, leg. R. Teruel & T. M. Rodríguez, 2♀ (paratypes), RTO; Los Hondones , 2–3 m a. s. l., June 1985, leg. J. Novo & L. V. Moreno, 1♀ juv. (paratype), IES, 1 February 2001, leg. R. Teruel, 2♀1♀ juv. (paratypes), RTO, 28 November–4 December 2012, leg. T. M. Rodríguez, 5♀1♀ juv. (paratypes), RTO, 22–25/January 2013, leg. R. Teruel & T. M. Rodríguez, 12♀9♀ juvs. (paratypes), RTO, 16 August 2014, leg. T. M. Rodríguez, 4♀2♀ juvs. (paratypes), RTO .
ETYMOLOGY. The selected epithet is a Latinized patronym honoring two good friends of us: the Cañizares brothers (Maikel and Maydiel), who greatly helped us during our repeated collecting trips to the type locality.
DIAGNOSIS (females only). Adult size medium for the genus (25–29 mm). Adult coloration: base yellowish brown, slightly darker on metasomal segments IV–V and slightly paler on legs, venter and telson; dark patterns essentially absent, except for faint brown reticulation in chelicera manus and faint infuscation in carapace (along anterior margin and around eyes) and metasomal segments II–IV (on ventral surface). Juvenile coloration: base pale orange to yellowish brown, slightly paler on pedipalps, legs, venter and telson; dark patterns essentially absent, except for faint gray reticulation in chelicera manus and faint infuscation in carapace (along anterior margin and around eyes) and metasomal segments II–V (mostly on posterior part and around carinae); tergites with posterior margins translucent whitish; pedipalps fingers not infuscate. Pedipalps remarkably elongate for the genus (length/width ratio of femur, patella and chela: 2.70–2.80, 2.54–2.61 and 3.65–3.74); chela with manus elongate-oval (length/width ratio: 1.53–1.61) and with fingers long (movable finger/manus length ratio: 1.31–1.39), fixed/movable fingers with 11–12/11–12 principal rows of denticles (mode 12/12). Tegument of carapace, tergites, sternites, metasoma and telson with a satin sheen. Carapace and tergites with many coarse granules scattered. Pectines with 11–14 teeth (mode 12–13); basal middle lamella greatly enlarged and oval. Sternite V with median smooth medium-sized, as long as wide and cordiform; lateral smooth patches small, oblique and slightly bulky. Metasoma slightly elongate (length/width ratio of segments I–V: 1.09–1.16, 1.40–1.47, 1.52–1.58, 1.71–1.88 and 2.26– 2.32), with 10/8/8/8/7 complete to almost complete, coarsely denticulate to serrato-denticulate carinae; lateral inframedians on II obsolete on basal half; intercarinal tegument of segment V with many large, conical granules. Telson with vesicle less globose than standard for the genus; subaculear tubercle small.
DESCRIPTION (♀ holotype; Figs. 2 View Figure 2 , 14 View Figures 13– 14 , 16 View Figures 15–16 , 18 View Figures 17–18 , 20 View Figures 19–20 , 22 View Figures 21–22 , 35a View Figures 34–35 ; Tabs. 2, 4 View Table 4 ).
Coloration ( Figs. 14 View Figures 13– 14 , 35a View Figures 34–35 ). Base light yellowish brown, slightly darker on metasomal segments IV–V (medium brown) and slightly paler on legs, venter and telson. Chelicerae manus very sparsely reticulate with brown (more densely distally); fingers faintly and irregularly infuscate. Pedipalps immaculate, with carinae and fingers slightly darker due to heavier sclerotization. Carapace largely immaculate, with faint symmetrical infuscation along anterior margin and around all three eye groups; eyes and ocular tubercles black. Tergites immaculate, with carinae of VII slightly darker due to heavier sclerotization. Pectines immaculate pale yellowish brown, with basal portion and basal plate progressively darker due to heavier sclerotization. Sternites immaculate; V with median and lateral smooth patches translucent. Legs immaculate, with carinae slightly darker due to heavier sclerotization. Metasoma with segments IV–V slightly darker (medium brown), with carinae darker due to heavier sclerotization and without any annulated pattern; all surfaces immaculate, except for ventral surface with a pair of very faint, elongate brownish spots between ventral submedian and ventrolateral carinae over distal half of segments II–IV. Telson immaculate, with distal half of aculeus dark reddish brown.
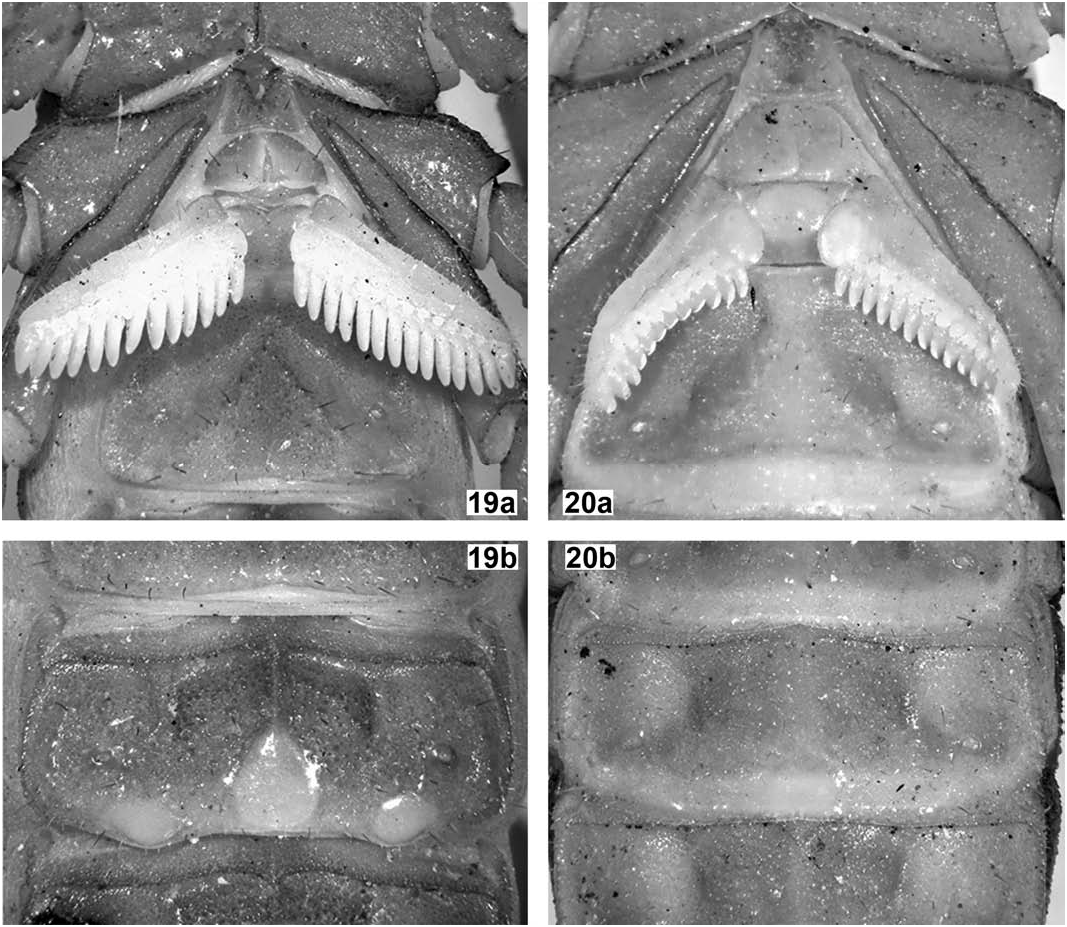
Chelicerae ( Fig. 18a View Figures 17–18 ). With dentition typical for the genus, teeth standard-sized and sharp. Tegument glossy but with minute granulation scattered, dorsodistal portion of manus with coarse, glossy granules irregularly arranged transversally around dorsodistal depression. Setation very dense ventrally, but essentially lacking dorsally, except for five rigid macrosetae around dorsodistal depression of manus.
Pedipalps ( Fig. 16a–c View Figures 15–16 ). Remarkably elongate for the genus, i.e., moderately longer and with chelae slenderer than standard. Femur essentially straight (very slightly bent inwards distally), with all carinae very strong, coarsely denticulate to serrate; intercarinal tegument with a satin sheen, very densely and irregularly granulose; internal (i) trichobothria surrounding a large, irregularly triangular spur. Patella straight, with all carinae strong, coarsely granulose to serrato-crenulate; intercarinal tegument with a satin sheen, very finely and densely granulose, internally with many conical denticles. Chela with manus elongate-oval (1.53 times longer than wide), wider than patella (ratio 1.19) and with the basal half slightly widest, all carinae strong, coarsely serrate to serratocrenulate, intercarinal tegument with a satin sheen, very finely and densely granulose on all surfaces; fingers long for the genus (movable finger 1.39 times longer than underhand) and evenly curved, fixed finger with 12/12 principal rows of denticles, movable finger with 12/12 plus an apical subrow of four denticles and a large internal accessory denticle (large terminal denticle not included), basal lobe/notch combination absent.
Carapace ( Fig. 18a View Figures 17–18 ). Wider than long (length/width ratio = 0.93). Anterior margin widely V-shaped, with two pairs of dark macrosetae. Carinae: anterior medians almost parallel (very slightly divergent anteriorly) and formed by very coarse, isolated granules and irregularly fused to the superciliaries, which are composed of much smaller and closer granules; posterior medians almost parallel (very slightly divergent posteriorly) and formed by coarse, isolated granules and irregularly fused to the central laterals and lateral oculars which are equally formed by coarse, isolated granules; other carinae absent or indistinct from general granulation. Furrows: anterior marginal, anterior median, median ocular, lateral oculars, central median, posterior median and posterior marginal all fused, wide and moderately deep; posterior laterals very long, oblique, wide and moderately deep, almost fused to posterior marginal; other furrows absent or indistinct. Tegument with a satin sheen, very densely and irregularly but finely granulose, with many coarse, rough granules scattered. Median eyes relatively large and separated by about one ocular diameter; lateral eyes noticeably smaller and largely concealed below anterolateral margin.
Sternum ( Fig. 20a View Figures 19–20 ). Standard for the genus, although more pentagonal in shape, with two pairs of dark macrosetae. Tegument minutely and densely granulose.
Genital operculum ( Fig. 20a View Figures 19–20 ). Standard for the genus, with three pairs of dark macrosetae. Valves roundly subtriangular and not separated medially; tegument coriaceous. Genital papillae absent.
Pectines ( Fig. 20a View Figures 19–20 ). Smaller than standard for the genus (barely reaching sternite III spiracles). Tooth count 13/12, teeth very short, straight and not swollen. Fulcra large and bulky, paraboloid to round. Basal middle lamella greatly enlarged and oval in shape. Pre-pectinal plate vestigial, poorly sclerotized, very wide and short; tegument smooth. Basal plate normally sclerotized, about as long as wide; anterior margin with only a subtle indication of anteromedian notch, posterior margin paraboloid; tegument smooth.
Legs. Slenderer than standard for the genus, with all carinae strong, finely denticulate to granulose. Intercarinal tegument finely and very densely granulose, with abundant slightly coarser granules scattered mostly over external surface, even defining an additional carina (externomedian) on lateral surface of femur of all legs. Claws short and strongly curved. Mesosoma ( Figs. 18b View Figures 17–18 , 20 View Figures 19–20 ). Tergites with a satin sheen, very densely and irregularly but finely granulose, with many coarse, rough granules scattered; carinae: median longitudinal strong, serrato-crenulate to serrate and progressively longer from I– VII, submedians absent on I–VI, complete, straight and strongly serrate on VII, laterals absent on I–VI, complete, straight and strongly serrate on VII. Sternites with a satin sheen, very densely and irregularly but finely granulose, with some dark macrosetae scattered all over; lateral margins of all sternites subserrate to subgranulose; posterior margin of all sternites essentially smooth, on III–IV widely bilobed, on V vestigially and widely bilobed (lacking any protuberance of lateral smooth patches), on VI essentially straight, on VII shallowly concave; carinae: III–V acarinate, VI-VII with paired laterals and submedians which are long, straight and finely crenulate; spiracles small, narrowly oval to short-oval; sternite V with median smooth patch medium-sized, cordiform (as long as wide), moderately bulky and glossy, not protruding from posterior margin; lateral smooth patches small, oblique, oval, slightly bulky and glossy, not protruding from posterior margin.
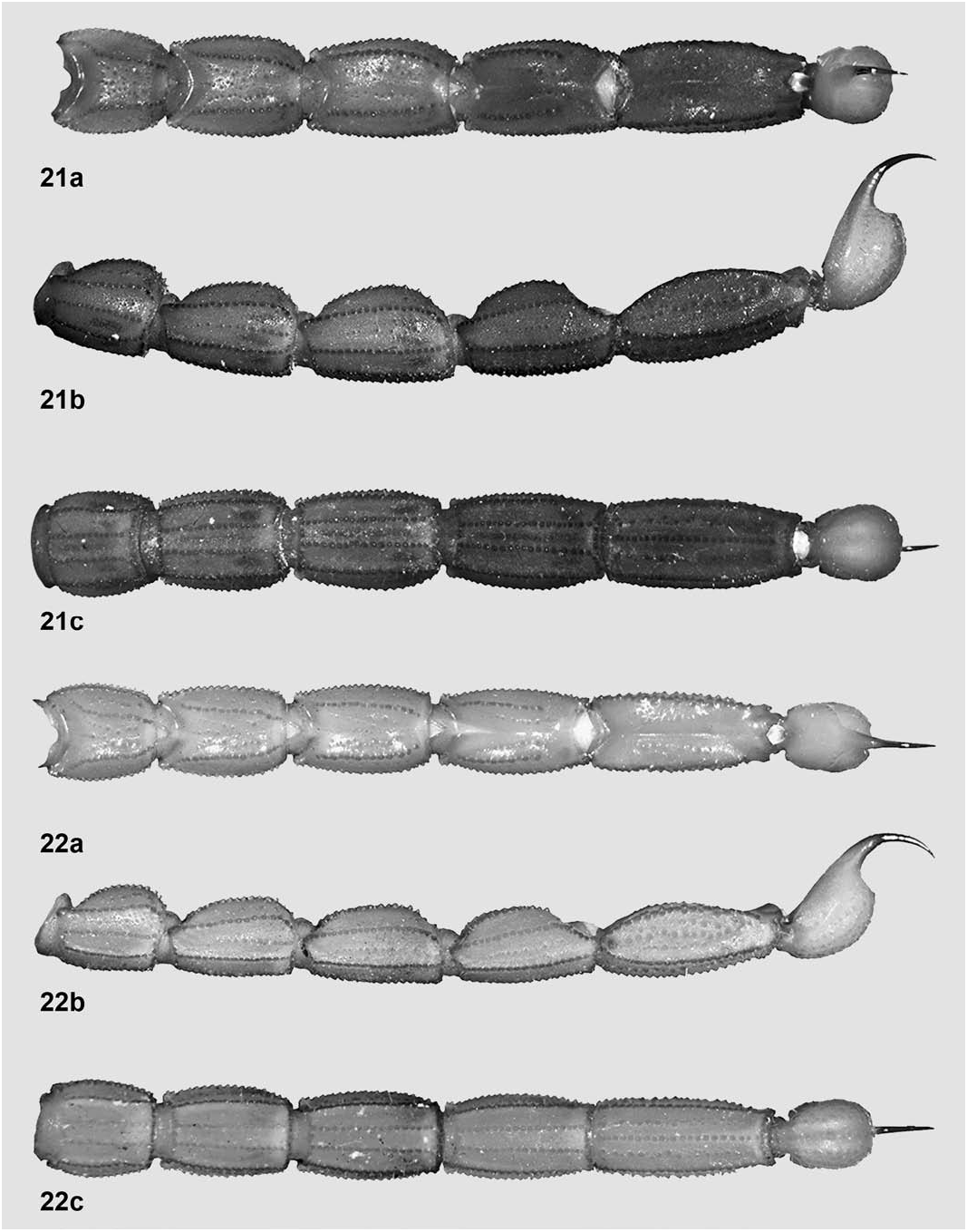
Metasoma ( Fig. 22 View Figures 21–22 ). Length standard for the genus (3.97 times longer than carapace) and slightly narrower distally. Segment I with ten complete to almost complete carinae, II–IV with eight, V with seven: dorsal laterals very strong, coarsely denticulate to serrato-denticulate on I–IV, absent on V but subtly indicated by an incomplete row of conical granules; lateral supramedians very strong, coarsely denticulate to serrato-denticulate on I–V; lateral inframedians very strong, coarsely serrato-denticulate on I, present only on distal half of II, absent on III–V (but indicated in III–IV by a few coarser, irregularly arranged granules and on V by a well-defined row of very coarse granules that almost define true carinae); ventral laterals very strong, coarsely serrate to serrato-denticulate on I–V; ventral submedians very strong, coarsely serrate to serrato-denticulate on I–IV, indicated by an incomplete but raised row of coarse granules on more than basal half of V; ventral median absent on I–IV, very strong, coarsely denticulate to serrato-denticulate on V. Intercarinal tegument with a satin sheen, very finely and densely granulose, with some medium-sized granules scattered on dorsal surface of all segments and with many large, conical granules on lateral surfaces of V; dorsal furrow complete, narrow and moderately deep on all segments; setation sparse, with two pairs of macrosetae on essentially all carinae.
Telson ( Fig. 16d View Figures 15–16 ). Vesicle less globose than standard for the genus (1.59 times longer than wide, exactly as wide as deep) and with several dark and whitish setae scattered; tegument coriaceous but with a satin sheen, with many medium-sized granules scattered ventrally and ventrolaterally; ventral median carina weak but coarsely subgranulose, continued into the subaculear tubercle that is obsolete. Aculeus standardsized, sharp, shorter than vesicle and moderately curved.
MALE. Unknown. This is apparently a female-only species that reproduces asexually by thelytokous parthenogenesis (see below, in Comments section).
VARIATION. The 40 paratypes are remarkably homogeneous for most morphological characters such as coloration, tegument sculpture, carination, setation and morphometric ratios, where variation was only minor and mostly due to eval differences, e.g., older adults have slightly darker coloration and setation, granulation and claws worn-out to different degrees. The only noteworthy exceptions are discussed as follows.
Adult size ( Tab. 2) varied from 24.9–29.0 mm. The maximum gap observed between two specimens was 4.07 mm (a 14% absolute difference), which suggests the existence of at least two distinct size-classes; on the contrary, within each putative size class specimens differed by not more than 1.2 mm (4%). Number of principal rows of denticles varied from 11–12 on both fixed and movable fingers, with a clearly defined mode of 12 in each case. Fixed finger was slightly more variable, with five fingers out of 66 (7.6%) having 11 rows; on movable finger, only two fingers out of 65 (3.1%) had 11 rows. All cases of fixed and movable fingers with 11 rows were singlefinger occurrences (most frequently on right pedipalp), i.e., the specimen had the three other fingers with 12 rows. An adult female paratopotype had right movable finger with seven rows only, but it was a teratology: about half of the denticles were irregularly aligned into a single, poorly defined row.
Pectinal tooth counts ( Tab. 4 View Table 4 ) varied from 11–14, with a clearly defined mode of 12–13. Occurrence of symmetric counts between left/right pectines was essentially as common as one-tooth asymmetry (20 vs. 16 individuals, respectively, two specimens excluded from analysis due to having one pecten either missing or too damaged for reliable counting); the only two naturally deviant specimens (i.e., not showing any evidences of teratology), had 11/13 and 14/12.
AFFINITIES (adult females only). This species is very easy to distinguish at first sight from the three other described species of the genus where this sex is known, by two unique characters: the metasomal segment V with intercarinal tegument very coarsely denticulate (especially on lateral surfaces) and the pedipalp chela remarkably more elongate and slender (3.65– 3.83 times longer than wide).
In the remaining species, metasomal segment V always has much weaker and sparser intercarinal granulation. Besides, pedipalp chela length/width ratio is much lower and compares as follows, enhanced by additional diagnostic features: T. mulata sp. n.: 1) size larger (32–34 mm); 2) coloration much darker (basically olivaceous brown) and very densely spotted all over with blackish brown; 3) pedipalp chela shorter and more robust (length/width ratio: 3.24–3.30); 4) tegument of carapace, tergites, sternites, metasoma and telson matt; 5) pectines with slightly but consistently higher tooth counts (12–15) and with basal middle lamella slightly smaller and more oval in shape; 6) sternite V with median smooth patch markedly narrower; 7) metasoma more robust, especially segment V (length/width ratio: 2.16–2.17); 8) telson vesicle more globose.
T. inaequalis : 1) size much larger (33–36 mm); 2) pedipalp chela shorter and more robust (length/width ratio: 3.43–3.51); 3) carapace and tergites with granulation finer; 4) pectines larger, with higher tooth counts (14–16) and with basal middle lamella slightly smaller and more oval in shape; 5) sternite V with median smooth patch smaller, longer than wide and narrowly cordiform in shape; 6) metasoma with carinae weaker and finer; 7) telson vesicle more globose.
T. inexpectata : 1) coloration basically light reddish brown, with carapace darker and pedipalp fingers deeply infuscate; 2) pedipalp chela shorter and more robust (length/width ratio: 3.31–3.48); 3) sternite V with median smooth patch shorter and wider than long; 4) metasoma markedly more robust (length/width ratio of segments I–V: 1.01–1.08, 1.31–1.35, 1.49–1.53, 1.63–1.70 and 2.01–2.06); 5) telson vesicle more globose.
The females of T. pumila sp. n. are still unknown, but it can be distinguished from T. canizaresorum sp. n. based on a nonsexually dimorphic character: the coloration remarkably darker (base orange-brown) and symmetrically patterned with dark brown on pedipalps (femur and patella), carapace, tergites and metasoma. Moreover, T. pumila sp. n. and T. canizaresorum sp. n. are two of the most widely allopatric members of Tityopsis , by occurring near the opposite longitudinal tips of the genus range ( Fig. 2 View Figure 2 ).
ECOLOGICAL NOTES. This species lives exclusively in mesophylous semicaducifolious forest on karstic soil ( Fig. 39 View Figures 39–40 ). Almost all specimens were collected during diurnal searches, under rocks of various sizes semi-buried in the leaf litter, usually hanging to the underside of the rock, as well as directly on the soil. In the type locality, T. canizaresorum sp. n. is more common inside the small collapsed depressions (dolines) that are locally called “hondones” and give the place its name.
Repeated nocturnal searches by the authors with UV light at Los Hondones, Playa Larga and other sites scattered all over Zapata Swamp, were completely unsuccessful to yield any specimens of this elusive species. Nevertheless, two juveniles were unintentionally found at night inside human-inhabited rooms: one walking on the floor at the type locality at about 22:30 hrs, the other on a hotel bed at Playa Larga at about 2:15 hrs. The latter stung one of the collectors in his back while sleeping and caused him excruciating pain for about two hours (Luis A. Casanella, personal communication).
In the three known localities, T. canizaresorum sp. n. lives syntopically with the buthids C. guanensis and H. junceus (which are also commonly found on the vegetation, especially the former), as well as with the diplocentrid Heteronebo morenoi ( Armas, 1973) , which is largely restricted to the seashore.
All collecting events listed above included pregnant females, thus, this species reproduces all year round. Five captive females gave birth to litters of 6–17 newborn, which underwent the first ecdysis 4–6 days later.
COMMENTS. Although not proved yet, there is strong factual evidence that T. canizaresorum sp. n. reproduces by obligate thelytokous parthenogenesis. Most adult females were collected pregnant, but not a single male is present in the entire sample (41 specimens, 26 of them adults) and the five females that gave birth in captivity, produced all-female broods.
We tried to get a direct, positive confirmation of parthenogenesis from all juveniles collected on 22–25 January 2013, by rearing them in captive isolation to maturity and then expect for any virgin parturition. They readily accepted as prey small beetle larvae ( T. molitor ) and cockroach nymphs ( B. germanica and P. americana ), but unfortunately a month later there was a massive, long-term anti-dengue fumigation campaign all over the city and despite all efforts, the entire breeding group was killed by envenomation, either directly or via contaminated prey.
To overcome this fatality, we turned to check the sex ratio in all our 179 samples of Tityopsis spp. from populations where both sexes have been found, following Francke (2007). We found that despite the species, at least one male (either adult or juvenile) was present in: 1) all populations with a minimum cumulative sample size of five specimens; 2) all individual samples composed of five or more specimens; 3) 84% of the individual samples composed of two to four specimens. There is no need for a statistical analysis to discard sampling error as the potential cause of the observed absolute absence of males in T. canizaresorum sp. n. Last, collecting events of this species span all along the year (January, February, June, August and November) and represent both dry and rainy seasons, thus, environmental sex determination and male birth/survival seasonality can also be safely disregarded as alternate hypotheses.
DISTRIBUTION ( Fig. 2 View Figure 2 ). This species is known only from three nearby localities enclaved in the karstic limestone coastal plain of southern Zapata Swamp. This coastal stripe is completely isolated from inland by extensive marshlands and permanently flooded areas.
The presence of T. canizaresorum sp. n. in adjacent areas of Cienfuegos Province (e.g., Aguada de Pasajeros and Abreus Municipalities) seems likely, as these areas are geographically very close to those where it is confirmed to occur (less than 25 km air-distance at the nearest point) and have the same types of vegetation and soil. A few scattered attempts to find it there by the present authors and some collaborators were all unsuccessful, but these searches were not intensive nor extensive enough to be conclusive.


| R |
Departamento de Geologia, Universidad de Chile |
| T |
Tavera, Department of Geology and Geophysics |
| V |
Royal British Columbia Museum - Herbarium |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.