
Vrijenhoekia balaenophila, Pleijel & Rouse & Ruta & Wiklund & Nygren, 2008
|
publication ID |
https://doi.org/10.1111/j.1096-3642.2007.00360.x |
|
persistent identifier |
https://treatment.plazi.org/id/03E287CA-1709-FFC2-FF21-16AB299C6C00 |
|
treatment provided by |
Felipe |
|
scientific name |
Vrijenhoekia balaenophila |
| status |
|
RELATIONSHIPS OF VRIJENHOEKIA BALAENOPHILA View in CoL
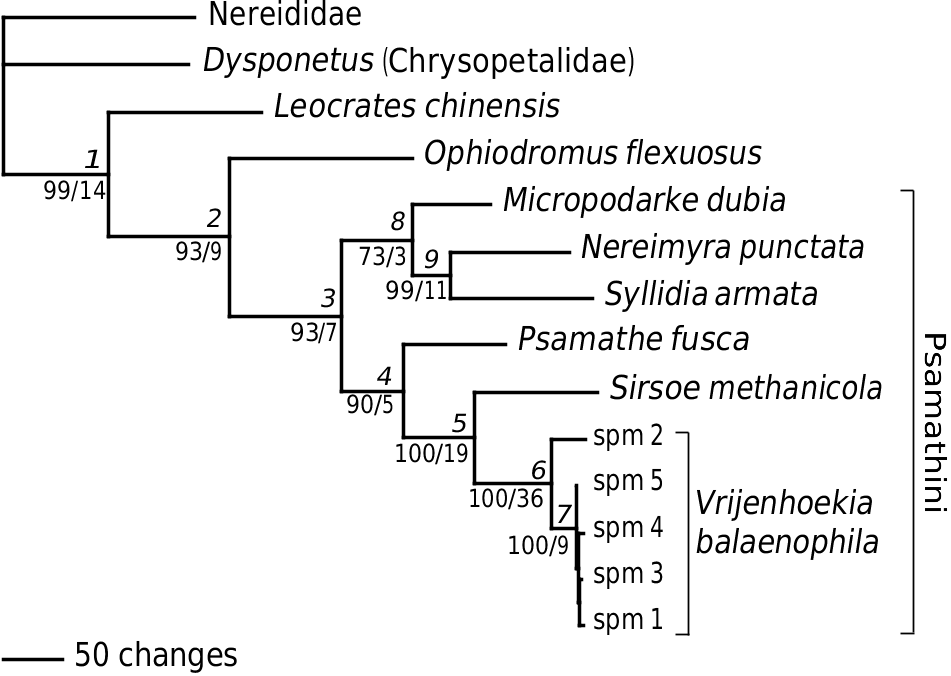
Vrijenhoekia balaenophila is clearly a member of Hesionidae , as seen by, for example, the presence of well-differentiated, cylindrical ventral cirrophores on the anterior segments ( Fig. 3D), but absence or much less well differentiated cirrophores on ‘normal’ segments, a feature that is uniqe within hesionids. Preliminary parsimony analyses based on molecular and morphological data (not included here) of Vrijenhoekia balaenophila together with a large selection of other hesionids provide strong support for a position within Psamathini . This assignment, however, relies heavily on molecular data, and the morphological evidence for both Psamathini and V. balaenophila is weak. Pleijel (1998) listed three apomorphies for Psamathini , including absence of facial tubercles, pattern of dorsal cirri alternation and prolonged teeth of chaetal blades. Of these, a facial tubercle reappears in some members, and the prolonged teeth are reduced in several taxa, including Vrijenhoekia .
The combined analysis of molecular and morphological data yielded two equally parsimonious trees with a tree length of 1718, a consistency index of 0.654 and a retention index of 0.5190. The difference between the two equally parsimonious trees is restricted to the Vrijenhoekia clade. Of the five specimens included, one (spm 2 in Fig. 5 View Figure 5 ) is separated from the others by a longer branch length; however, re-examination of this specimen gives no support to the possibility that it belongs to a different species. It may indicate multiple colonizations by V. balaenophila of this whale carcass, but this requires further data and different analytical approaches.
The jackknife analysis provides > 50% support for ten nodes ( Fig. 5 View Figure 5 ), eight of which have a > 90% support. The separate analyses are largely in concordance with the combined result. All separate analyses, as well as the combined one, provide support for a sister relationship between Sirsoe and Vrijenhoekia as well as support for a clade of the five specimens of Vrijenhoekia (nodes 5 and 6 in Fig. 5 View Figure 5 and Table 4). There are only two cases in which there is jacknife support > 50% for conflicting resolutions in the separate data sets compared with the combined result. First, a sister group relationship between Micropodarke and Psamathe has 57% supported in 16S, and 74% support in the morphological data set, conflicting *Node in conflict with jackknife support > 50% (value shown in parentheses).
A dash indicates support for this node or alternative resolution < 50%.
values between COI and the other partitions following removal of the third codon positions (see text) with nodes 4 and 9 in the combined analysis ( Fig. 5 View Figure 5 , Table 4). Secondly, a clade with Leocrates chinensis and Ophiodromus has 79% support by morphology, conflicting with node 2 in the combined analysis ( Fig. 5 View Figure 5 , Table 4). Relating to the same conflicts, the PBS analysis shows negative Bremer values for nodes 2 and 4 for 16S and morphology. The largest negative PBS values are found in the COI data set for node 3 (all hesionids except Ophiodromus and Leocrates ) and for node 10 ( Nereimyra and Syllidia ) ( Fig. 5 View Figure 5 , Table 4), but there is no alternative topology supported with a jacknife value > 50%. These negative PBS values are also reflected in the ILD test, which shows the COI data set to be incongruent with all the three other partitions ( Table 5). Saturation in COI, associated with long branch attraction (e.g. Bergsten, 2005), would be a possible cause, and in order to examine this we removed the third codon positions of COI and re-ran the ILD test with the other partitions ( Table 5). However, this yielded no change in the significance of the ILD values and we therefore consider saturation to be an unlikely cause. We find this incongruence noteworthy but cannot provide any explanations, and see no justifications for data removal.
The resolution of the phylogenetic relationships within Psamathini has varied among previous analyses ( Pleijel, 1998; Pleijel & Rouse, 2000; Ruta & Pleijel, 2006; Ruta et al., 2006). Here, two clades are well supported, ( Nereimyra , Syllidia ) and ( Sirsoe , Vrijenhoekia ), of which the first is present in all previous analyses as well. The sister group to ( Sirsoe , Vrijenhoekia ) is here Psamathe , although the support is not high (jackknife 90, Bremer 5), and previous studies have suggested Micropodarke or ( Nereimyra , Syllidia ) as sisters to this clade, in place of Psamathe .
Sirsoe currently includes S. grasslei and S. methanicola , both of which, in contrast to Vrijenhoekia , have a median antenna, no glandular lip pads and well-developed neuropodial lobes on segment 3. From a phylogenetic point of view the choice between including this new hesionid in Sirsoe or to erect a new genus is largely arbitrary. However, for practical reasons we have here opted for the second alternative, simply because the two taxa differ in a number of morphological characters and a diagnosis would include many polymorphisms.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.