
Plesionika ensis (A. Milne Edwards, 1881 )
|
publication ID |
https://doi.org/10.5281/zenodo.188076 |
|
DOI |
https://doi.org/10.5281/zenodo.6216263 |
|
persistent identifier |
https://treatment.plazi.org/id/265887F1-FD03-FFE7-FF2A-E54E484096FD |
|
treatment provided by |
Plazi |
|
scientific name |
Plesionika ensis (A. Milne Edwards, 1881 ) |
| status |
|
Plesionika ensis (A. Milne Edwards, 1881) View in CoL
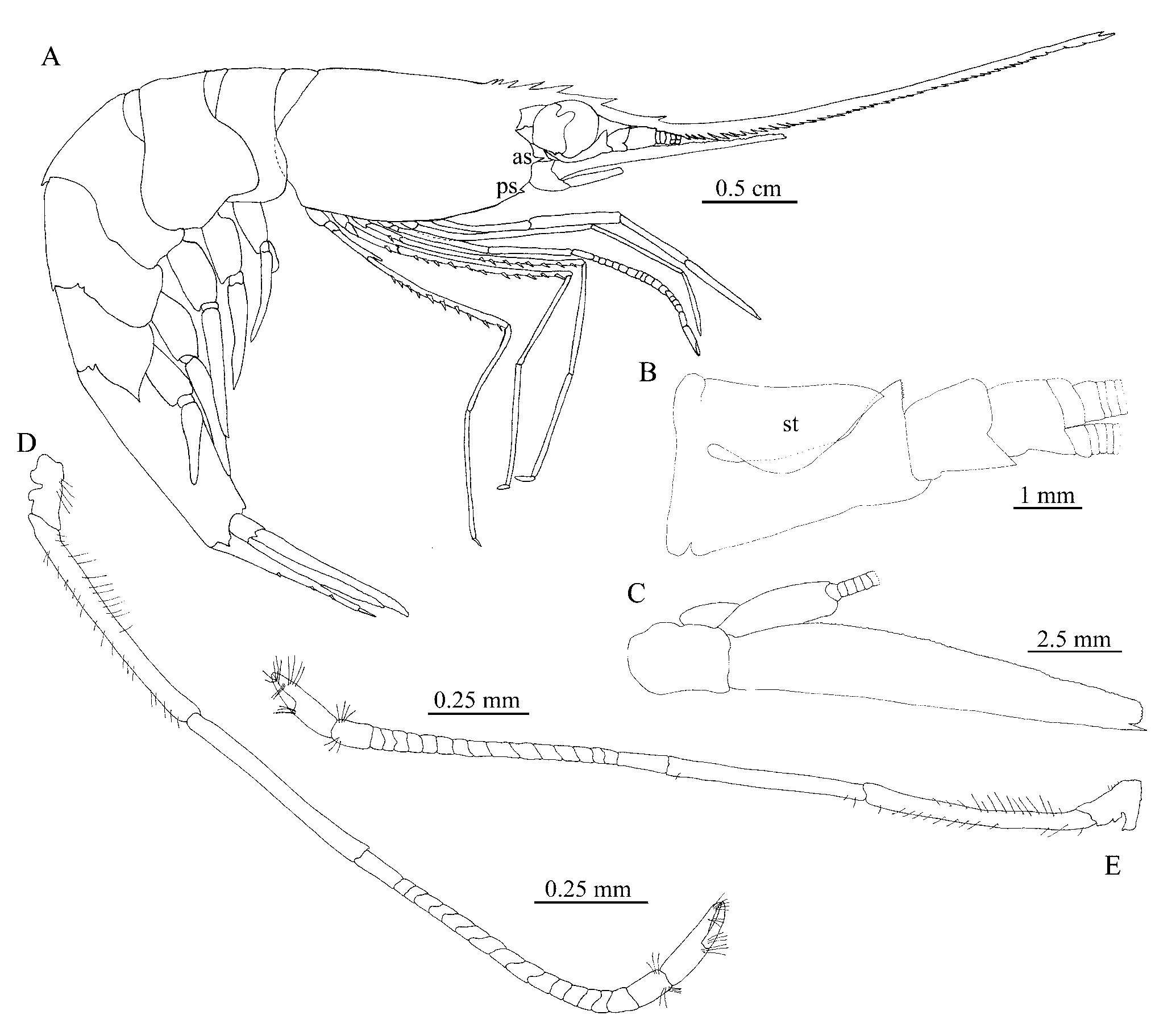
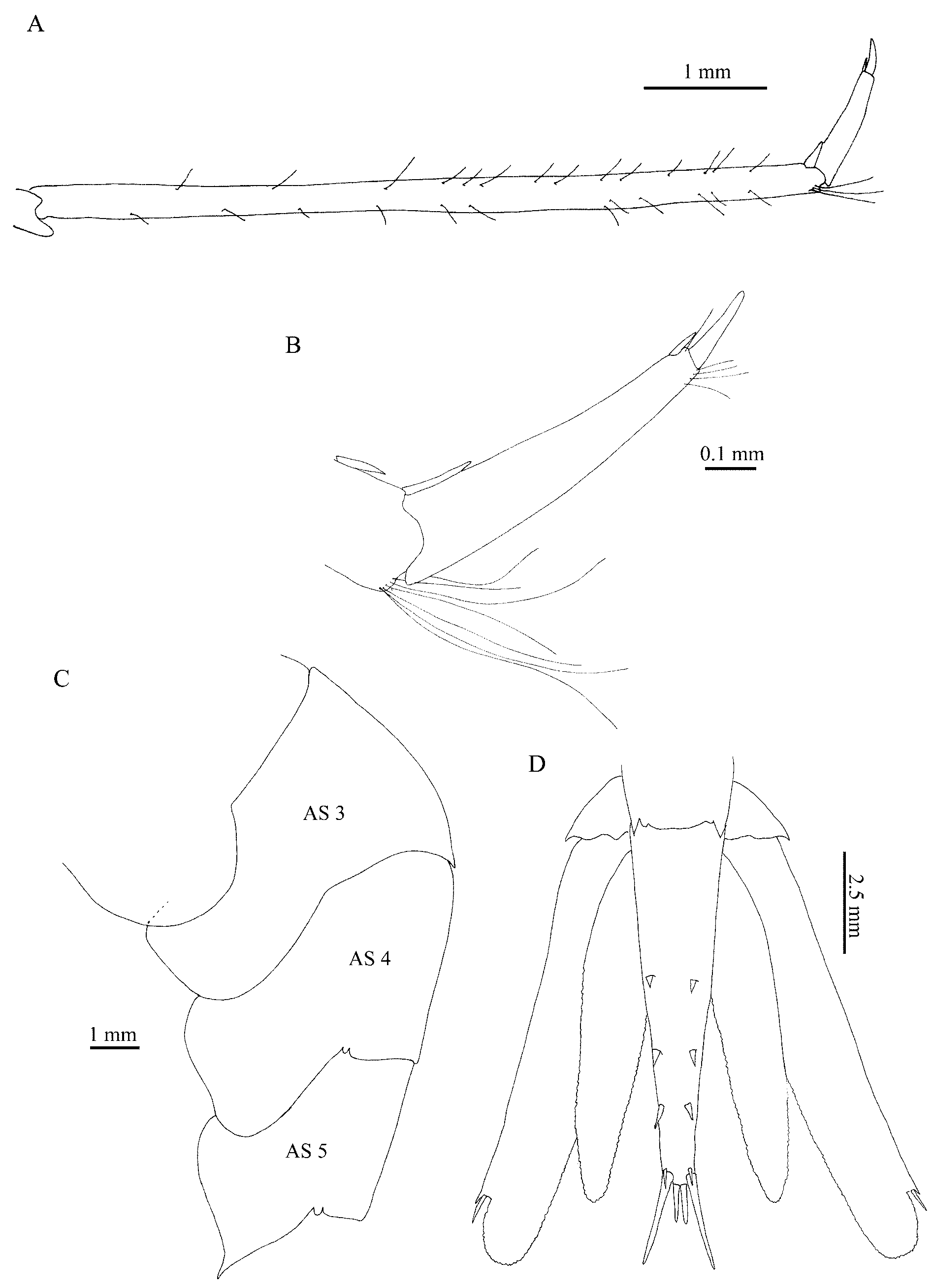
( Figs. 3 View FIGURE 3 , 4 View FIGURE 4 )
Acanthephyra ensis A. Milne Edwards, 1881: 14 ; Young, 1900: 476. Pandalus ensis View in CoL – A. Milne Edwards, 1883: pl. 18; Faxon, 1896: 161; Alcock, 1901: 96; Coutiére, 1905: 675; Rathbun, 1906: 914.
Plesionika uniproducta Bate, 1888: 641 View in CoL , pl. 113, fig. 1 (part); Moreira, 1901: 8 (part). Plesionika semilaevis Bate, 1888: 664 View in CoL (part).
Plesionika ensis View in CoL – De Man, 1920: 106; Holthuis, 1951: 55, fig. 10; Crosnier & Forest, 1968: 1138; 1973: 209, figs. 63c, 64f; Pequegnat, 1970: 94; Omori, 1971: 241; Cabral et al., 2000: 246; Cruz & Fransen, 2004: 141; Viana et al., 2007: 36.
Material examined: REVIZEE: E- 0 500, 13o22’S, 38o40’W, 394 m, 1 male ( 12 mm), 1 ovigerous female ( 14 mm), MNRJ 14668; E-0513, 15o53´S, 38o02´W, 489 m, 4 males ( 13–15 mm), 1 female ( 13 mm), 3 ovigerous females ( 13–14.5 mm), MNRJ 14658; E-0518, 13o21’S, 38o38’W, 518 m, 1 male ( 21 mm), 1 female ( 17 mm), MNRJ 14660; D- 0 464, 21o48´S, 40o01´W, 618 m, 1 male ( 13.5 mm); MNRJ 13735.
Description: Rostrum long, curved downwards in proximal region, and upwards beyond antennular peduncle; far overreaching scaphocerite, 2.3–2.6 (avg. 2.4, n=6) times as long as carapace; ventral margin with 40–51 (avg. 43, n=6) teeth closely disposed, dorsal margin with one to three (avg. 2, n=11) large teeth well spaced, reaching antennular peduncle end, one distal tooth and two to three post-rostral teeth the proximal one variably movable; eye large, spherical, with ocellus; strong antennal and small pterygostomian spines present; carapace smooth ( Fig. 3 View FIGURE 3 A). Stylocerite straight at base and tapering only near the triangular apex, not overreaching the distal margin of first antennular peduncle article ( Fig. 3 View FIGURE 3 B). Scaphocerite 0.94–1.05 (avg. 0.99, n=12) as long as carapace, with blunt apex, distal tooth strongly overreaching blade ( Fig. 3 View FIGURE 3 C). Maxilliped III with epipod, penultimate segment 0.76–0.93 (avg. 0.84, n=13) times longer than terminal segment. Pereopods 1–4 with epipod, pereopod 5 without epipod. Pereopod 2 chelate, equal in size ( Fig. 3 View FIGURE 3 D, E), carpus with 15–28 (avg. 17, n=12) articles. Pereopod 3 overreaching scaphocerite with propod distal third; propod 0.45–0.55 (avg. 0.48, n=10) times as long as carapace; 4.3–7.6 (avg. 5.8, n=11) times longer than dactyl ( Fig. 4 View FIGURE 4 A), dactyl with two stout setae ( Fig. 4 View FIGURE 4 B). Dorsal surface of abdominal somite 3 with a straight spine; pleura of somite 3 rounded, pleura of somite 4 triangular but not acute, pleura of somite 5 triangular and acute ( Fig. 4 View FIGURE 4 C). Telson 0.72–0.92 (avg. 0.77, n=13) times as long as abdominal somite 6; not sulcate in dorsal midline, with three pairs of dorsolateral stout setae, and three pairs of stout distal setae ( Fig. 4 View FIGURE 4 D).
Distribution: Western Atlantic: Florida, Gulf of Mexico, Antilles, Brazil (Maranhão, Paraíba, Alagoas, Bahia, Rio de Janeiro). Eastern Atlantic: Senegal, Gabon, Congo, Angola. Indian and Pacific: Andaman Sea, Arabian Sea, Hawaii, Fiji. Adults in Atlantic founded in depths from 230 to 732 m, in Indo-West-Pacific in depths from 101 to 1251 m, the shallowest record is in Hawaiian waters at 55 m (modified from De Man, 1920; Crosnier & Forest, 1973; Cabral et al., 2000).
Remarks: Working with material from Brazilian waters ( 9o05`N / 34o50`W, Alagoas), collected by the Challenger Expedition, Bate (1888) described Plesionika uniproducta based on two specimens, one male and one female. According to Crosnier & Forest (1973), the female is actually P. e n s i s and the male is actually Plesionika martia (A. Milne Edwards, 1883) . In this way, Bate (1988) was the first to record P. e n s i s and P. martia to Brazilian waters, both identified as P. uniproducta .
According to Chan & Crosnier (1997) Plesionika ensis is in a group together with Plesionika reflexa Chace, 1985 . These species are very similar as pointed by Chace (1985), Crosnier (1986), Chan & Crosnier (1997) and Fransen (2006). Chace (1985) compares the type series of P. reflexa with specimens identified as P. e n s i s from various parts of the world and gives several characters that might prove to be of specific significance. The proportional length of dactyl and propod of pereopod 3 being 0.17–0.25 in the 14 specimens of P. e n s i s from western Atlantic and 0.30–0.46 in P. reflexa from South China Sea and Philippines; in the Brazilian material this proportion is 0.13–0.23, with the average of 0.18 (n=11) fitting with the range of western Atlantic P. e n s i s. Populations of P. e n s i s from East Atlantic, however, have a similar ratio (0.26–0.40), as P. re f l e x a material cited above ( Chace, 1985; Fransen, 2006). In Hawaiian material of P. e n s i s there are two forms, one with a very short dactyl (0.12–0.16) and other with a range intermediate (0.21–0.30) between that of the western and Eastern Atlantic populations ( Chace, 1985).
Other feature cited by Chace (1985) as useful to distinguish both species is the dorsal spine on abdominal somite 3 with a tendency to recurve upwards in P. reflexa , whereas no such inclination has been noticed in typical form of P. e n s i s. Chan & Crosnier (1997) mentioned that the absence of any specimen with a recurved dorsal spine in the Atlantic is sufficient reason to not synonymize P. reflexa with P. e n s i s. In the 13 Brazilian specimens examined herein the dorsal spine on abdominal somite 3 is straight ( Fig. 4 View FIGURE 4 C).
The scaphocerite distal tooth also may be of taxonomic importance, in P. reflexa it slightly overreaches blade, while in P. e n s i s it strongly overreaches blade ( Chace, 1985; Fransen, 2006), as was observed in Brazilian material ( Fig. 3 View FIGURE 3 C). Chan & Crosnier also pointed that the western Atlantic population of P. e n s i s presents a rostrum 2.2–2.8 (avg. 2.5) times carapace length, with 35–48 ventral teeth (avg. 42, n=6). The material herein examined fits in this range, with rostrum 2.3–2.6 (avg. 2.4, n=6) times as long as carapace and ventral margin with 40–51 teeth (avg. 43, n=6).
Chan & Crosnier (1997) comment that many more species may be also present in the material now identified as P. e n s i s and P. reflexa (as occur with the great diversity of species contained in the “ P. m a r t i a (A. Milne Edwards, 1883)” ( Chace, 1985) and “ P. narval (Fabricius, 1787) ” ( Chan & Crosnier, 1991) groups). Or the problem of these two species may be similar to the case of P. e d w a rd s i i and P. crosnieri , in which both species are distributed in the Indo-Pacific but only one occurs in the Atlantic ( Chan & Yu, 1991; Chan & Crosnier, 1997).
| MNRJ |
Museu Nacional/Universidade Federal de Rio de Janeiro |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
InfraOrder |
Caridea |
|
Family |
|
|
Genus |
Plesionika ensis (A. Milne Edwards, 1881 )
| Cardoso, Irene 2009 |
Plesionika ensis
| Viana 2007: 36 |
| Cruz 2004: 141 |
| Cabral 2000: 246 |
| Omori 1971: 241 |
| Pequegnat 1970: 94 |
| Crosnier 1968: 1138 |
| Holthuis 1951: 55 |
| De 1920: 106 |
Plesionika uniproducta
| Moreira 1901: 8 |
| Bate 1888: 641 |
| Bate 1888: 664 |
Acanthephyra ensis A. Milne Edwards, 1881 : 14
| Rathbun 1906: 914 |
| Coutiere 1905: 675 |
| Alcock 1901: 96 |
| Young 1900: 476 |
| Faxon 1896: 161 |
| Milne 1881: 14 |