
Tychobythinus inopinatus Sabella, Costanzo and Nicolosi, 2020
|
publication ID |
https://doi.org/10.11646/zootaxa.4779.2.8 |
|
publication LSID |
lsid:zoobank.org:pub:F187FD14-DDE5-4FD5-AD5C-554FB22579D3 |
|
DOI |
https://doi.org/10.5281/zenodo.3851211 |
|
persistent identifier |
https://treatment.plazi.org/id/40039BFF-19D6-4966-B466-9D31DA9FEC5F |
|
taxon LSID |
lsid:zoobank.org:act:40039BFF-19D6-4966-B466-9D31DA9FEC5F |
|
treatment provided by |
Plazi |
|
scientific name |
Tychobythinus inopinatus Sabella, Costanzo and Nicolosi |
| status |
sp. nov. |
Tychobythinus inopinatus Sabella, Costanzo and Nicolosi View in CoL , sp. nov.
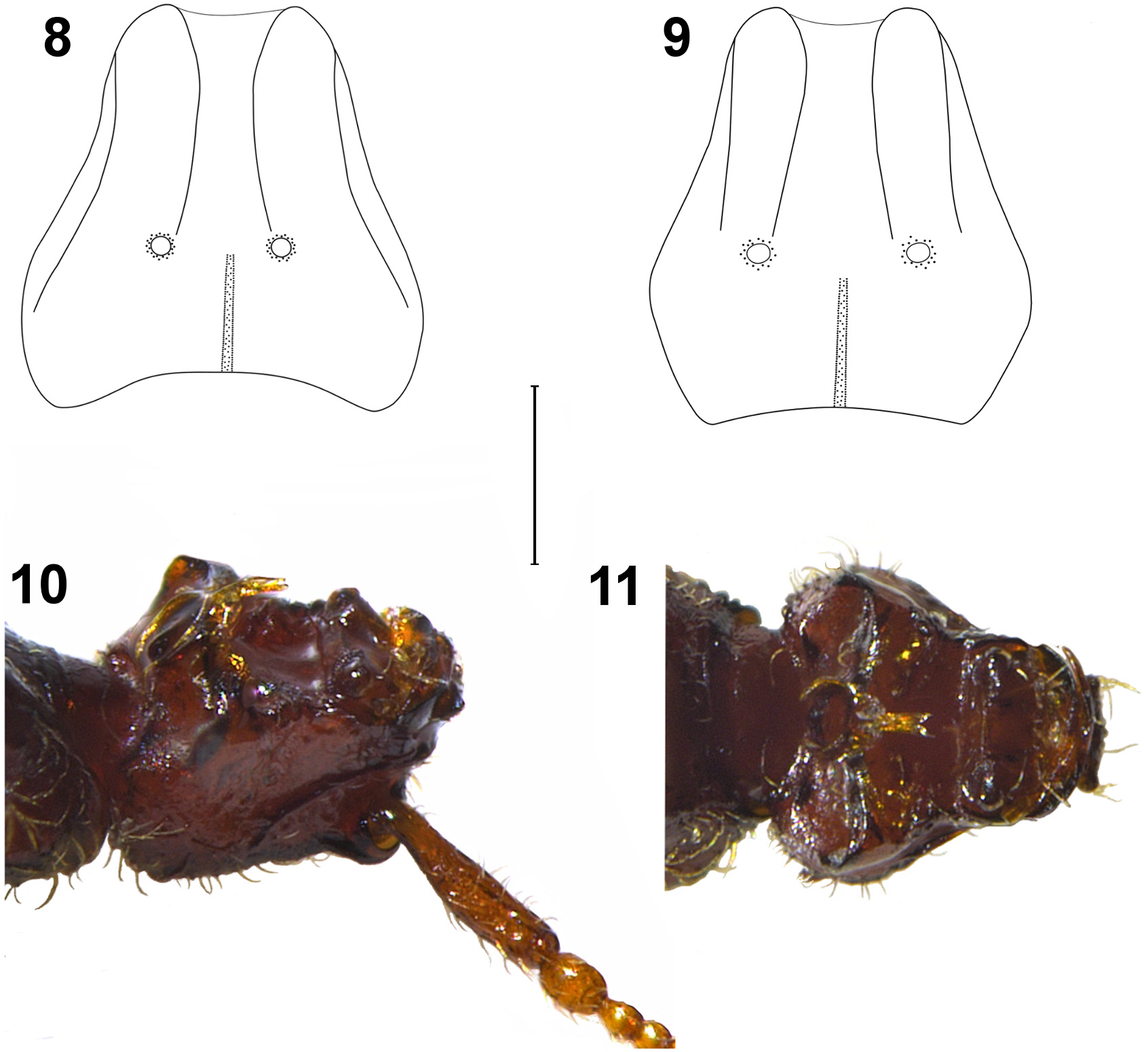
( Figs. 6–16 View FIGURES 6–7 View FIGURES 8–11 View FIGURES 12–16 )
urn:lsid:zoobank.org:act:
Type material. Holotype: ITALY: Sicily Region: Syracuse, Monello Cave , 22.X.2019, trap 17, 1 ♂, G. Nicolosi leg. ( DBUC) . Paratypes (all 17 specimens in DBUC): ITALY: Sicily Region : 1 ♀, same data of holotype ; 1 ♀, same data of holotype, but trap 8 ; 1 ♀, same data of holotype, but trap 10 ; 1 ♂, same data of holotype, but trap 12 ; 1 ♀, same data of holotype, but trap 16 ; 1 ♀, same locality, 31. VII.2019, trap 10, G. Nicolosi leg. ; 2 ♀♀, same data, but trap 12 ; 1 ♀, same locality, 22.VIII.2019, trap 12, G. Nicolosi leg. ; 1 ♀, same locality, 05.IX.2019, trap 12, G. Nicolosi leg. ; 1 ♀, same locality, 19.IX.2019, trap 17, G. Nicolosi leg. ; 1 ♀, same locality, 07.X.2019, trap 10, G. Nicolosi leg. ; 1 ♀, same data, but trap 17 ; 1 ♀, same locality, 07.XI.2019, trap 10, G. Nicolosi leg. ; 3 ♀♀, same locality, 05.XII.2019, trap 10, G. Nicolosi leg.
Description. Male ( Fig. 6 View FIGURES 6–7 ): Length 1.30–1.35 mm, apterous and anophthalmous. Pale brown or brown with lighter antennae, palpi, and legs. Pubescence consists of long and flattened setae (length: 0.07–0.08 mm) on head, pronotum, elytra, and abdomen, other suberect shorter setae (length: 0.03–0.04 mm) on antennae, and legs, and thin fluff on apical segment of maxillary palpi.
Head ( Fig 8 View FIGURES 8–11 ) wider ( 0.30 mm) than long ( 0.24 mm) narrower than pronotum. Frontal lobe wider ( 0.175 mm) than long ( 0.08 mm) with subparallel, protruding and sharp sides; antennal tubercle protruding. Frons between antennal tubercles with large median sulcus reaching anterior edge of vertexal foveae. Median clypeal carina welldefined, equally visible in dorsal as well as lateral view, extending to ocular region. Tempora rounded, convex occipital region traversed by median longitudinal sulcus, extending to posterior edge of vertexal foveae, these last wide and well-impressed. Gular region ( Figs 10–11 View FIGURES 8–11 ) behind labium with deep and broad impression margined pos- teriorly by transverse ridge projecting and laterally recurved backwards on each side of head. This ridge with long and acute median process, process projecting ventrally and ending with a bifid tuft of setae on the tips. Base of median process enlarged posteriorly in an oval impression, at base with two long and sturdy median bristles. Antennae 0.65–0.66 mm long, scape more than 4 times longer ( 0.19–0.20 mm) than wide ( 0.045 mm), narrowed and flattened with sharp medial margin on basal third, wider at middle. Pedicel ovoid, slightly asymmetric, about 1.5 times longer than wide, about as wide as scape, and wider than funicular segments. Antennomere III distinctly longer than wide and narrowed at base; antennomeres IV and V about as long as wide, antennomeres VI–VII wider than long, antennomeres VI shorter and closer than VII; antennomeres VIII wider than long, and wider than VII. Antennal club consisting of last three antennomeres which are widening progressively from IX to XI. Antennomeres IX and X distinctly wider than long, antennomere XI distinctly longer than wide and twice as long as combined length of antennomeres IX and X. Maxillary palpi ( Fig. 13 View FIGURES 12–16 ) with palpomere IIs elongate and gradually expanded from base to apex, their surface covered by 8-10 tubercles. Palpomeres III slightly longer than wide, surface with 4–6 tubercles; last palpomere about 5 times as long ( 0.29 mm) as wide ( 0.055 mm), widest at basal third, lateral margin distinctly curved and sinuate at middle.
Pronotum wider ( 0.31 mm) than long ( 0.295 mm), widest near middle, anteriorly narrowed with convergent sides, posteriorly very slightly narrowed with rounded and subparallel sides very flattened laterally. Dorsal surface shiny with some sparse and faint punctures. Two well-impressed antebasal lateral foveae linked by wide antebasal sulcus. Tegument between pronotal posterior margin and antebasal sulcus rough, makeing it difficult to see small median antebasal fovea. Metaventrite distinctly raised at middle, its surface with dense and large punctures, with median sulcus beginning from its posterior margin and extending to just posterior to mesocoxal cavities. Base of mesocoxal cavities with pubescent lateral mesosternal fovea on each side.
Elytra distinctly wider ( 0.57 mm) than long ( 0.52 mm), convex, sides slightly rounded from base to the apex, widest near middle. Humeral calli strongly reduced. Dorsal surface shiny with only some superficial punctures. Each elytron with two basal foveae, subhumeral fovea well-defined. Both marginal and sutural striae reaching to about elytral apices, discal striae lacking.
Abdomen normally shaped without species-specific characters.
Legs relatively long and thin. Protrochanters with 1–2 tubercles on ventral surface, protibiae slightly flattened and sinuate at distal third, protarsomere II slightly dilated. Meso- and metatrochanters simple; femora simple, mesotibiae slightly enlarged and sinuate for distal third, length of metatibiae: 0.45–0.46 mm, slightly enlarged and sinuate for distal third.
Aedeagus ( Fig. 14 View FIGURES 12–16 ) 0.325–0.33 mm long, ovoid with relatively short parameres, parameres convergent and with narrow apex, each bearing two subapical setae. Internal sac ( Figs 14–15 View FIGURES 12–16 ) with two long apophyses converging to apices with numerous spines.
Female ( Fig. 7 View FIGURES 6–7 ): Similar to male, length 1.30–1.35 mm, head ( Fig. 9 View FIGURES 8–11 ) slightly wider ( 0.25–0.26 mm) than long ( 0.23–0.24 mm), gular region unmodified, slightly convex. Antennae ( Fig. 12 View FIGURES 12–16 ) a little shorter (length: 0.63-0.64 mm) and with sligthly thinner antennomeres than in male; surface of palpomere II covered by 10–14 tubercles, anterior sides of pronotum narrower than in male. Metaventrite lacking median impression; legs with protibiae, tarsomere II of protarsi, and metatibiae unmodified. Telisternite as in Fig. 16 View FIGURES 12–16 .
Discussion. Tychobythinus inopinatus sp. nov. differs from all other congeneric species by its aedeagal and exoskeletal features. It shares affinities with Tychobythinus villasmundi Sabella, Amore, Nicolosi, 2019 , however it differs from the latter in numerous characters: antennal scape more than 4 times longer than wide (less than 4 times longer than wide for T. villasmundi ); last palpomere about 5 times longer than wide, with lateral margin distinctly curved and sinuate at middle (about 4 times longer than wide with lateral margin slightly curved and sinuate at middle for T. villasmundi ), different shape of gular ridge of male (cfr. Figs 10–11 View FIGURES 8–11 with Figs 8–9 View FIGURES 8–11 in Sabella et al. 2019), aedeagus with different apical morphology of the parameres, which are not narrowed in T. inopinatus sp. nov., and are distinctly narrowed in T. villasmundi (cfr. Fig. 14 View FIGURES 12–16 with Fig. 12 View FIGURES 12–16 in Sabella et al. 2019), and differences of the internal sac (cfr. Figs 14–15 View FIGURES 12–16 with Fig. 12 View FIGURES 12–16 in Sabella et al. 2019).
Like T. villasmundi , T. inopinatus sp. nov. shares affinities to the congeneric species from North Africa which belong to two distinct groups ( Sabella et al. 2014). However, they do not represent, with certainty, two homogeneous phyletic lineages because their external morphology is closely related to their different levels of adaptation to endogean life.
The aedeagus of T. inopinatus sp. nov. is comparable to the species belonging to the Tychobythinus algiricus group ( Sabella et al. 2014; Chiasmatobythus sensu Jeannel 1956 ), due to their troglobitic adaptations. T. inopinatus sp. nov. is externally close to the species belonging to the Tychobythinus theryi group ( Sabella et al. 2013; Anopsibythus sensu Jeannel 1956 ).
Among the material examined we also found a female collected in Monello Cave, 06.VI.2019, trap 13, G. Nicolosi leg. ( DBUC), with only 10 antennomeres, and with antennomere VI distinctly longer than wide, which is probably derived from the fusion of antennomers VI and VII. This specimen has been excluded from the type series .
Finally, it seems interesting to underline that the previous research conducted inside Monello Cave in 1991 ( Caruso 1995), which used the same collection techniques, did not produce any specimens of the new species. On the contrary, our investigation that began on 11 January 2019 and is still in progress allowed us to collect, in addition to the species relieved in the previous study, 18 specimens ( 2 males and 16 females) of the new species of Tychobythinus in various areas of the cave, but only during the period from August to December 2019.
This fact could be explained considering that the ecological conditions inside the cave have improved signifi- cantly in the last thirty years, probably in relation to the implemented strict conservation measures, but the factors driving this change certainly deserve further study (with a careful comparison of the environmental data emerged from the study conducted in 1991 and in 2019), which however goes beyond the purpose of this article.
A simpler explanation can be provided by the different sampling periods for the two research programs: research in 1991 covered the period January–July, while our research program, still in progress, covered the entire year of 2019, with collection of the new species happening between August and December, a period not covered by the previous research program.


| VI |
Mykotektet, National Veterinary Institute |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Pselaphinae |
|
Genus |