
Molinodus suarezi Muizon & Marshall, 1987
|
publication ID |
https://doi.org/ 10.5252/geodiversitas2019v41a25 |
|
publication LSID |
urn:lsid:zoobank.org:pub:4EF01B8A-BA1D-4F14-9432-CA8754D765D2 |
|
DOI |
https://doi.org/10.5281/zenodo.3704858 |
|
persistent identifier |
https://treatment.plazi.org/id/291A87B5-FFE2-1E08-FCC5-F0D41D3BFB1C |
|
treatment provided by |
Valdenar |
|
scientific name |
Molinodus suarezi Muizon & Marshall, 1987 |
| status |
|
Molinodus suarezi Muizon & Marshall, 1987
EMENDED DIAGNOSIS. — Size similar to that of Promioclaenus; dental formula I?/3, C?/1, P?/4, M3/3; P3 triangular, with a strong parastyle anterior to paracone; pre- and postparacristae well-developed; protocone is a well-developed cusp and distincly individualized from the paracone but slender with slight pre- and postcingula; P4 more massive, shorter, and wider than P3, with small conules; protocone only slightly larger than on P3 but more massive; wellmarked pre- and postcingula, and strong labial cingulum; M1 triangular and almost symmetrical in relation to its transverse axis; protocone transversely compressed with an oval-shaped apical wear facet; metacone only slightly lingual to the paracone; welldeveloped cingula (pre-, post- and labial); styles slightly to not projected labially; no hypocone; M2 subquadrangular and strongly asymmetrical, with oblique labial edge; well-developed pre- and postcingula, not reaching the para- and metastyles (but close to them); labial cingulum very strong; protocone large, bulbous, and mesiodistally elongated with incipient duplication; conules large; well-developed para- and metacingula reaching para- and metastyles; paracone higher and more voluminous than metacone; metacone much more lingual than paracone; centrocrista straight; para- and metastyles almost aligned with para- and metacones but parastyle still slightly shifted labially; no well-individualized hypocone; M3 much wider than long; strongly bent posteriorly; labial edge strongly oblique; little reduced (i.e. only slightly shorter than M2); metacone and metaconule reduced; i1 and i2 distinctly larger than i3; i2 staggered; lower canine short and robust; p1 single-cusped, single-rooted, and procumbent; p2 triangular in lateral view; transversely flattened; small posterior cusp; p4, much larger; variable in shape from triangular to quadrate, with a large metaconid appressed against protoconid; anterior crest of protoconid possessing a tiny paraconid; large talonid cusp; lower molars with bulbous cusps; trigonid and talonid basin reduced; strong pre- and small postcingulids (on m1 and m3 only); paraconid clearly smaller than and appressed against metaconid; paracristid transverse and arched distally; metaconid distolingual to and slightly smaller than protoconid; cristid obliqua variable in size and reaching labial edge of metaconid; talonid basin small and open lingually (more an oblique groove than a basin); hypoconid large, inflated, circular, and only slightly smaller than protoconid; entoconid and hypoconulid almost completely fused forming a distolingual oblique crest; talonid of m3 larger than on m1-2 with hypoconulid as large as hypoconid. On the dentary presence of a well-developed coronoid crest separated from the labial edge of m3 by a coronoid fossa.
Among Tiupampa “condylarths”, Molinodus more resembles Simoclaenus than the other taxa. However, Molinodus differs from Simoclaenus in its smaller size, its cheek teeth more elongated mesiodistally, its upper molar less transverse, its M2 more asymmetrical with an mesiolabially projected parastyle, its procumbent p1, and its longer rostrum.
Molinodus differs from Promioclaenus in its molars, which are more bulbous with apices of the cusps more approximated, in the longer trigonid of the lower molars with a paraconid less appressed against the metaconid, in the presence of a generally distinct labial cingulum, in the non-reduced m3, in the more transverse and less bulbous upper molars, in the thinner postcingulum, in the strong asymmetry of the M2 with an mesiolabially projecting parastyle, and in the unreduced M3.
HYPODIGM. — As in Muizon & Cifelli (2000) with the additional following specimens: MHNC 13883, a partial left mandible with alveoli of incisors, canine, p1, roots of p2, alveoli of p3 and alveolus of anterior root of p4; MHNC 13867, a partial left mandible with p3-m3; MHNC 13870, a partial left maxilla with M1-M3, with M1 and M2 missing the labial edges of para- and metacones.
DESCRIPTION
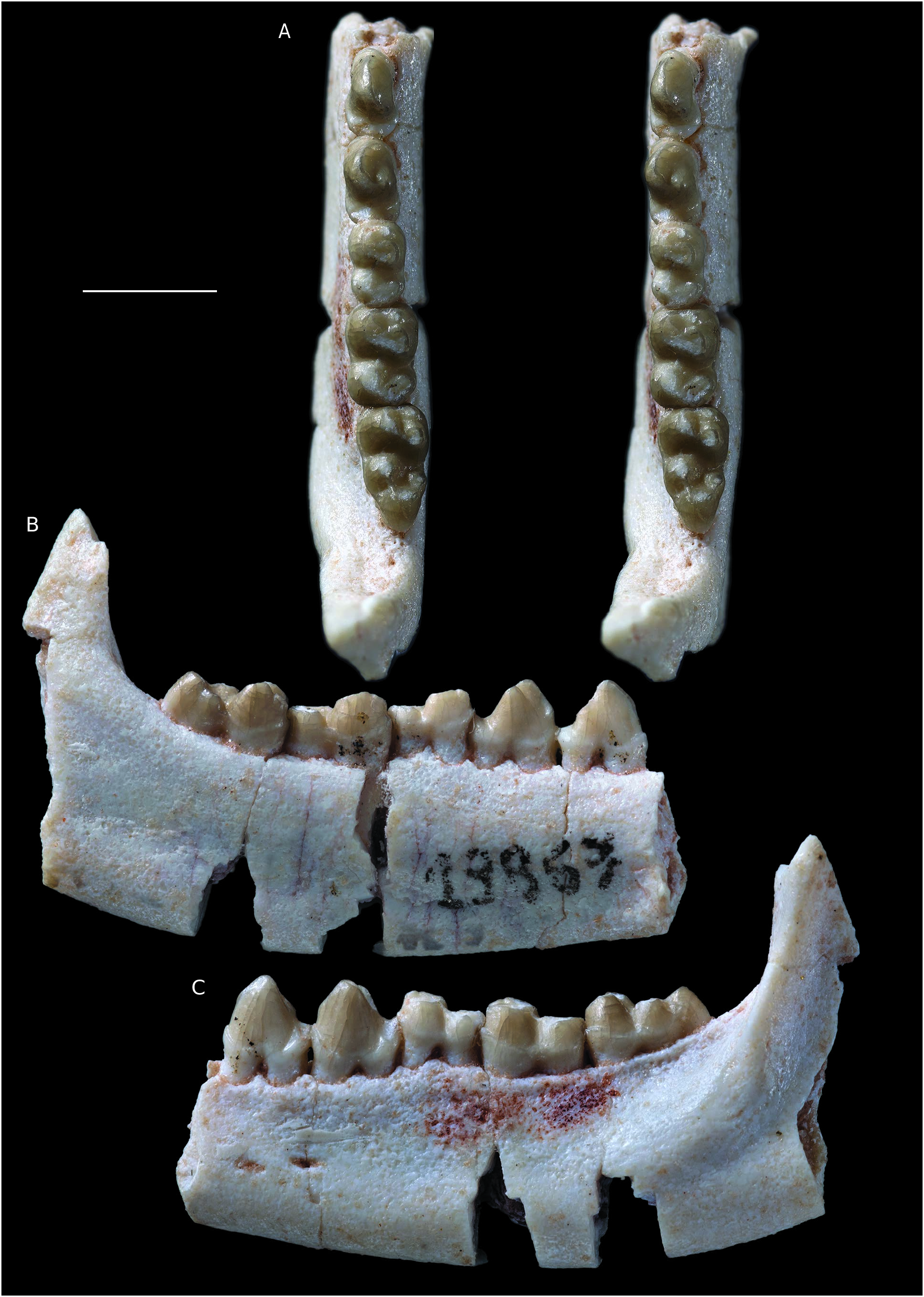
An anterior fragment of left mandible of Molinodus suarezi ( Fig. 1 View FIG ) bears the labial edge of the alveoli of the three incisors, the canine, the p1, the roots of p2, the alveoli of p3, and the anterior alveolus of p4 (MHNC 13883). On the medial aspect of the dentary a large symphyseal surface for the intermandibular suture is present. This surface is in one plane with sharp edges, but it is rough and bears numerous ridges and grooves probably interlocking with the symmetrical surface on the other mandible. The two symphyseal surfaces were therefore tightly attached one to the other, but the symphysis was clearly unfused on this specimen. Although ligamentous, it is likely that the symphysis was quite rigid and allowed little intermandibular movements. Because the roots of p3 extends as far as the ventral region of the dentary leaving no space of a potential unerupted tooth germ, we regard that the definitive p3 was erupted. Because the preserved teeth (c and i1) are absolutely unworn, this specimen is interpreted as belonging to a young adult. It is, therefore, not impossible that ontogenetically older individuals may have had a fused symphysis.
On the labial side of the dentary a large anterior mental foramen is present ventral to the embrasure between p1 and p2.
Although the incisors are missing, the anteriormost region of the dentary is preserved with the labial edge of the incisors alveoli. The dentary is broken vertically in this region and the lingual portion of the alveoli is missing; therefore, a vertical section of the incisors alveoli can be observed. Based on the size of the roots, i1 was apparently the largest of the three incisors or was, at best, similar in size to i2. The i3 is clearly the smallest of the three incisors. Interestingly, the alveolus of i2 presents a distinct lingual shift of its root. As a consequence, the vertical section the alveolus is clearly triangular whereas that of the other alveoli are cylindrical with parallel edges ( Fig. 1B, E View FIG ). This condition of the i2 of Molinodus , is identical to the staggered second incisor, (the i3 according to Hershkovitz 1982, 1995), of many metatherians (e.g., stagodontids, pucadelphyids, sparassodonts, didelphids, microbiotheres, peramelids, thylacinids, dasyurids). The triangular section of the alveolus of the staggered incisor is clearly observed on the figure 5 of Hershkovitz (1982). It is not the first time that a staggered second incisor is described in a eutherian since Hershkovitz (1982: 197) mentions this condition in several extant Carnivora. Among the Tiupampa eutherians, it is absent in Alcidedorbignya inopinata ( Muizon et al. 2015), and there is no indication that it was present in Tiuclaenus minutus and Pucanodus gagneri, the only other Tiupampa eutherians that preserve the anterior portion of the dentary. It may be the first time that this trait is observed in a fossil eutherian, but this is probably due to the fact that the anterior portion of the dentary is rarely preserved in fossil mammals.
In dorsal view, the three lower incisors are set in a slightly oblique row, relative to the symphyseal plane ( Fig. 1C View FIG ). In other words, i1 is anteromedial to i2, and i3 is posterolateral to i2. The lower incisor tooth row was, therefore, more or less parabolic or V-shaped. The last incisor, i3, was closely appressed against the mesial edge of the canine as indicated by the posi- tion of the i3 alveolus. The canine is a moderately developed tooth but it is robust. The height of the crown (4.60 mm) is approximately twice its mesio-distal length (2.44 mm) at base.
The canine is transversely compressed, being distinctly narrower (1.75 mm) at its base than long (2.44 mm). The posterior curvature of the canine is weak and its posterior edge is only very slightly concave. Its mesial edge is distinctly convex. On the mesiolingual edge of the tooth a blunt ridge extends mesioventrally on the dorsal half of the crown and distoventrally on the ventral half ( Fig. 1B View FIG ). In dorsal view, the labial side of the canine is strongly convex, whereas it is almost flat to slightly concave on its lingual side. In labial view the main axis of the crown is moderately oblique in relation to the alveolar plane, with which it forms an angle of approximately 110°.
Posterior to the canine is a small p1 separated from the former by a small but distinct diastema. The p1 is single-rooted. It is slightly procumbent and asymmetrical in labial view. The crown of the tooth is markedly compressed transversely. It bears a pronounced mesial crest, which is slightly convex. The distal edge of the tooth is straight and bears a small cuspule at its base ( Fig. 1D View FIG ). There is no cingulum at the base of the crown. The lingual aspect of the crown in markedly concave and its labial edge is convex. As a consequence, in distal view the crown is slightly bent lingually.
The roots of p2 are preserved in the dentary of MHNC 13883. The very short diastema, which separates p2 from p1, is approximately twice shorter than the diastema anterior to p1. A similar condition is observed on MHNC 1243, a mandible fragment, which preserves the root of c, the alveolus of p1 and p2 (see Muizon & Cifelli 2000). Interestingly, on MHNC 13883 the roots of p2 are set obliquely in the dentary and the anterior root is mesiolabial to the posterior one. The mesiodistal axis of the tooth is approximately at an angle of 30° with the anteroposterior axis of the tooth row (from c to p4) ( Fig. 1D View FIG ). Because this condition is not present in MHNC 1243, it could represent a pathological condition or an individual variation in MHNC 13883. Posterior to p2, the alveoli of p3 indicate that the tooth was implanted parallel to the axis of the tooth row. The p2 and p3 were not separated by a diastema and were closely appressed one against the other.
A relatively complete left mandible of Molinodus suarezi (MHNC 13867) bears p3-4 and m1-3 ( Fig. 2 View FIG ). The most interesting element of this specimen lies in its premolars. The teeth are relatively narrow transversely being c. 30% longer than wide. They are roughly triangular in lateral view and are approximately as high as long. The p3 is slightly more slender than the p4 but both teeth are built on a similar pattern. The mesial edge of p3 bears a sharp crest with a marked angulation on the dorsal third of the crown. The largest cusp of the tooth, the protoconid, is transversely narrow and its lingual edge is slightly concave lingually, in its dorsal region. More ventrally it is inflated and convex. The distolingual angle of the protoconid bears a strong inflation, which reaches the dorsal half of the distal edge of the tooth in lingual view. This structure corresponds to the development of a small metaconid. At the mesial base of the tooth is a small cingulum, which is continuous from the distolabial to distolingual edges of the mesial root. Distally a well-developed talonid cusp forms the base of the crown and extends transversely in a robust and rounded cingulum from the distolabial to the distolingual angle of the tooth. In occlusal view, p3 is distinctly wider distally than mesially.
The p4 is slightly longer than the p3. Its mesial edge bears a sharp crest that shifts lingually at mid-height of the crown. There, it forms a small tubercle interpreted as an incipient paraconid. The protoconid is strongly convex labially and flat lingually. On its distolingual edge is a large metaconid. In dorsal view, it is slightly smaller than the protoconid and its apex is distinctly distal to that of the protoconid. On the mesial edge of the protoconid, a sharp and well-developed paracristid connects to the paraconid mesially. Posterior to the trigonid, the talonid does not bear differentiated cusps but forms a thick distobasal ridge, which extends from the mesiolingual to mesiolabial aspects of the distal root. The distal edge of the tooth is slightly wider than the mesial one.
The condition of the p4 of MHNC 13867 distinctly departs from the morphology of the isolated p4 (MHNC 1244) referred to Molinodus suarezi by Muizon & Cifelli (2000: fig. 3M, N). On MHNC 1244 the metaconid is almost totally lingual to the protoconid and not shifted distally. Furthermore, the crown of the tooth is markedly more massive, strongly compressed mesiodistally, and more inflated transversely. As a whole, MHNC 1244 more resembles the p4 of Simoclaenus sylvaticus in all its features but is clearly much smaller. When compared to the size of the roots observed on the holotype of S. sylvaticus , it is even too small to be a p3 of and it is also slightly too small to be a p2. In other respects, the large metaconid and the well-developed paraconid and paracristid of MHNC 1244 are more compatible with the morphology generally observed on a p4. Therefore, we think that MHNC 1244 is actually a p4 and that, either, the morphology of the p4 of M. suarezi is highly variable or MNHC 1244 represents a new taxon at Tiupampa. Given the size difference it is unlikely that it could be referred to S. sylvaticus , except if this species is sexually dimorphic, which is unknown yet. However, given the lack of a representative sample of M. suarezi and S. sylvaticus , this uncertainty cannot be resolved so far.
The three molars have the characteristic features of Molinodus suarezi as described by Muizon & Cifelli (2000). The trigonid bears a well-developed paraconid but distinctly smaller than the metaconid and placed mesial to it or slightly labial (on m1). The paraconid is closely appressed against the metaconid. The latter is large, almost as large as the protoconid and located distolingual to it. As a consequence, the protocristid is obliquely oriented (mesiolabially-distolingually) as compared to the axis of the tooth row. On m1 and m2 the talonid is slightly narrower than the trigonid. The cristid obliqua extends from the large hypoconid to the central region of the protocristid. Lingually, the hypoconulid and the entoconid are connate and the talonid basin is in fact an obliquely oriented groove that opens mesiolingually as is observed in all kollpaniines. On m3 the hypoconulid is much larger than on the other molars and is as large as the hypoconid.
The small size of the m1 relatively to the other molars is noteworthy. This feature is present in all kollpaniines but it is especially pronounced in the holotype (YPFB Pal 6112) of M. suarezi ( Muizon & Cifelli 2000: fig. 3A) and MHNC 13867 ( Table 1 View TABLE ).
On the dentary of MHNC 13867, the posteriormost mental foramen is located below the anterior root of p4. The corpus mandibularis of MHNC 13867 and 13871 is relatively massive especially in the latter. On both specimens, most of the ramus mandibularis is missing, except the anterior portion of the coronoid process. In the region of the retromolar space, the anterior basis of the coronoid process is very wide and bears a conspicuous labially opening coronoid fossa. The fossa is much deeper on MHNC 13871 than on 13867. Lateral to the fossa, the coronoid crest is salient and extends ventrally on the lateral aspect of the dentary ( Fig.3 View FIG ). This condition indicates a powerful masseter muscle, which inserts in the masseteric fossa and on the coronoid crest and is congruent with the stoutness of the corpus mandibularis. A robust coronoid crest is observed on the other specimens of M. suarezi (YPFB Pal 6114, MHNC 8269) and on the large kollpaniines ( Simoclaenus and Andinodus). It is absent or barely present in Tiuclaenus and Pucanodus , which are the smallest kollpaniine taxa (see Muizon & Cifelli 2000).
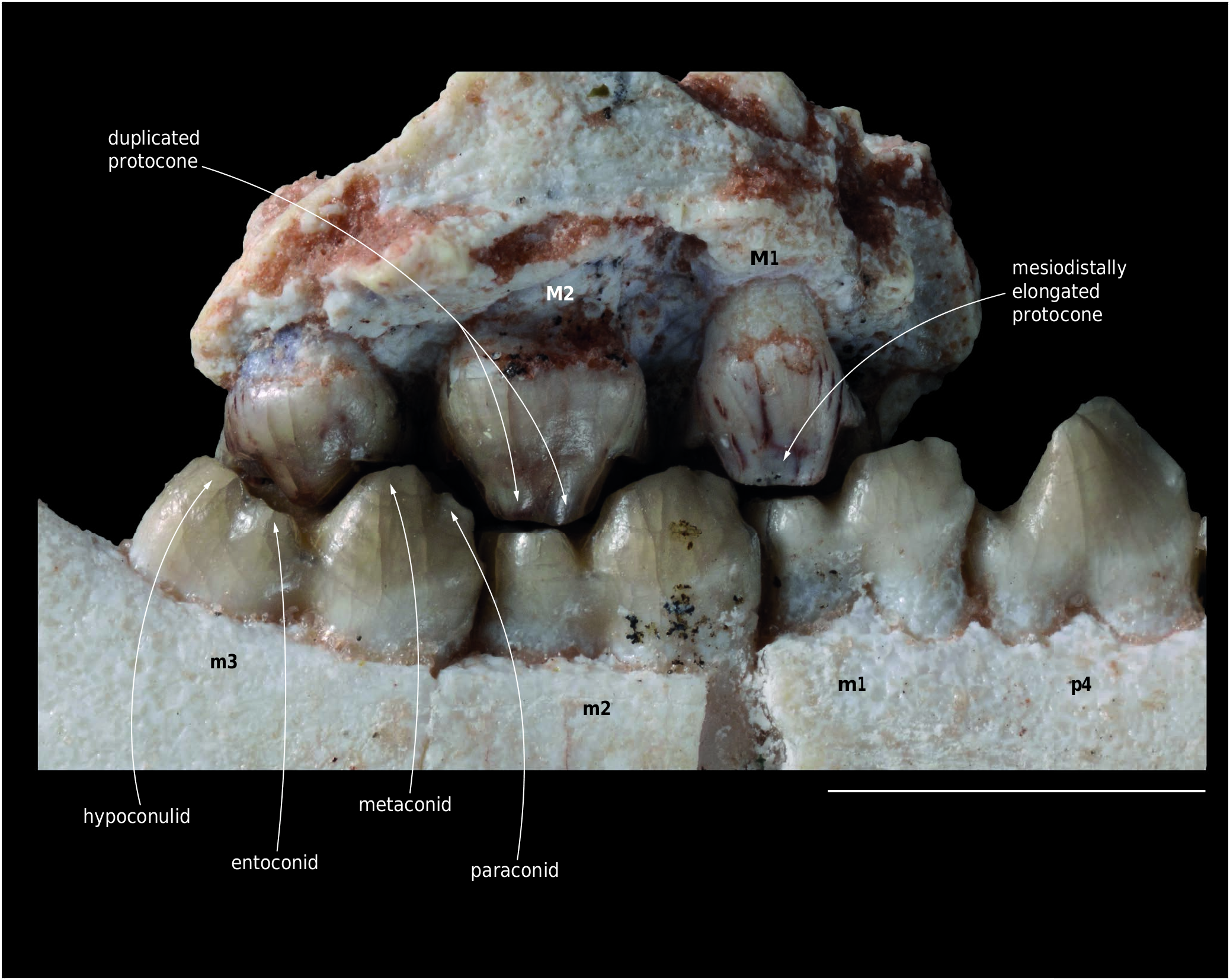
A partial left maxilla referred to Molinodus suarezi (MHNC 13870) bears the three molars and is the first specimen discovered with an M3 associated to the anterior molars ( Figs 4 View FIG ; 5 View FIG ; 6 View FIG ). The M3 referred by Muizon & Cifelli (2000: fig. 2, E) to M. suarezi is an isolated specimen and its referral had yet to be confirmed. M1 and M2 of MHNC 13870 are incomplete and are missing most of the labial edges of the para- and metacones and all the stylar regions and labial cingulum. M3 of MHNC 13870 is complete.
M1 and M2 of MHNC 13870 preserve the lingual half of the tooth. The large protocone bears large conules which are similar to those already described in Molinodus . The mesial and distal edges of M1-2 present well-developed pre- and postcingulae, which extended labially until the stylar area of the teeth. The most interesting cusp of this specimen is the protocone. On M1-2 as is observed on the molars of M. suarezi described by Muizon & Cifelli (2000), the protocone is mesiodistally elongated and its wear facet is markedly ovale-shaped. On the M2 of MHNC 13870, the wear facet is constricted and the protocone bears vertical grooves, which reach the apex of the cusp. As a consequence, the latter is distinctly bilobate ( Fig. 5 View FIG ). The posterior lobe of the protocone of MHNC 13870 receives the postprotocrista, which links it to the metaconule. This bilobate condition is even more pronounced than in MHNC 8280 ( Muizon & Cifelli 2000: fig. 2A) as it concerns more than the apex of the cusp and extends dorsally on the protocone. It indicates that the duplication of the apical region of the protocone was almost certainly observable on the dentine at the enamel-dentine joint (see Anemone et al. 2012).
The M3 of MHNC 13870 is remarkably similar to the isolated tooth referred to Molinodus suarezi by Muizon & Cifelli (2000) and does not require further description.




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |