
Cocalodinae
|
publication ID |
https://doi.org/ 10.5281/zenodo.186069 |
|
DOI |
https://doi.org/10.5281/zenodo.6226943 |
|
persistent identifier |
https://treatment.plazi.org/id/335D87D7-5E08-1F11-FF11-5818266EAF0D |
|
treatment provided by |
Plazi |
|
scientific name |
Cocalodinae |
| status |
|
Subfamily Cocalodinae Simon
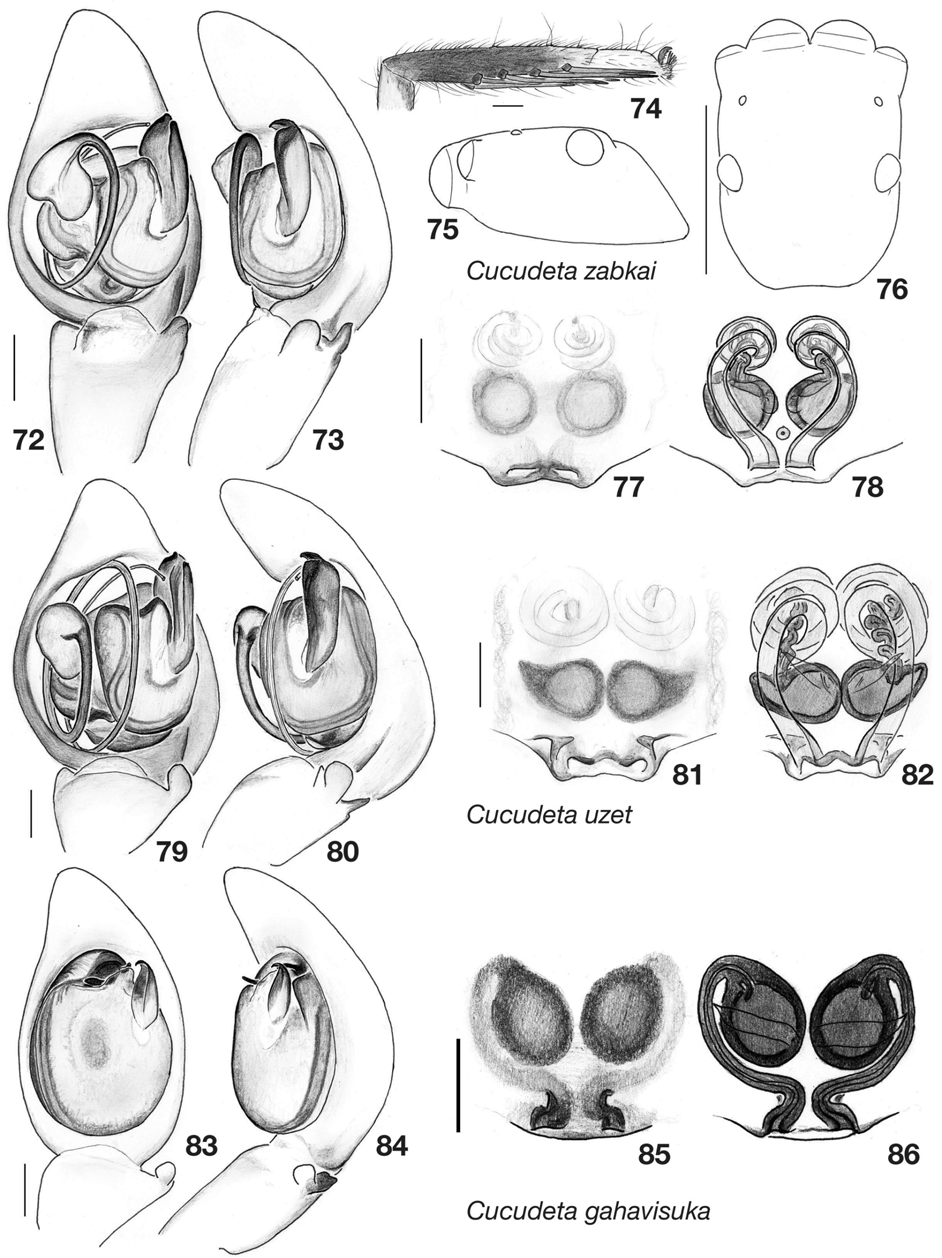
A median apophysis on the male palpus is a striking rarity in salticid spiders. When present, it is a sclerite just clockwise from the base of the embolus (in a left palp, ventral view), surrounded by hematodocha, and at least partially surrounded by the tegulum ( Wanless 1982, 1985; Maddison 2006; Figs 26 View FIGURES 20 – 30 , 57 View FIGURES 51 – 61 , 73 View FIGURES 72 – 86 ). The sperm duct typically approaches it before turning counter-clockwise into the embolus. Wanless (1982) assumed the median apophysis was derived within salticids and thus was evidence that Cocalodes , Allococalodes , and the African Holcolaetis Simon and Sonoita Peckham and Peckham form a monophyletic group. However, the median apophysis is probably ancestral for salticids, based on its widespread presence throughout araneomorph spiders ( Coddington 1990), in particular among families related to salticids (e.g. Bosselaers & Jocqué 2002; Ramírez 2003; Silva 2003; Benjamin et al. 2008), and based on its distribution among basal salticids in recent molecular phylogenies ( Maddison & Needham 2006, Maddison et al. 2007). If so, then the presence of a median apophysis does not indicate to what clade a salticid belongs, but rather to what clades it doesn't belong — a salticid with a median apophysis is a member of neither the Salticoida nor the Spartaeinae , both of which have lost it. [The sclerite reported as a "median apophysis" from the salticoid Tarne Simon by Szûts and Rollard (2007) is almost certainly not homologous to that in basal salticids: Tarne' s sclerite is counterclockwise from the base of the embolus in the left palp ventral view, and is not cradled by the tegulum.]
Wanless (1985) later preferred the hypothesis that Holcolaetis and Sonoita formed a clade with the spartaeines to the exclusion of Cocalodes . The placement of Cocalodes is therefore unresolved. Wunderlich (2004) synonymized the Spartaeinae with the Cocalodinae , choosing Simon's family-group name Cocalodeae because of priority over Wanless's name Spartaeinae . I reject this synonymy primarily because Cocalodes can be excluded from the Spartaeinae in Wanless's strict sense (1984) because it lacks two spartaeine synapomorphies, a tegular furrow and loss of the median apophysis. Even if these cocalodines and the spartaeines were found to be sister groups, we could still retain them as separate, as I do here. Wunderlich's broad sense of Cocalodinae , which includes the Baltic Amber salticids, is united as far as we know only by plesiomorphic character states (presence of conductor and median apophysis, many retromarginal cheliceral teeth, large posterior median eyes). Here I take a much more restricted concept of the Cocalodinae , to include only the five genera discussed below. The Baltic Amber salticids, other than the hisponines, are therefore Salticidae incertae sedis.
I will not here attempt to resolve the placement of cocalodines in the phylogeny of the Salticidae , as I have found no morphological synapomorphies that link it with particular subgroups of salticids. Some of the cocalodines I describe here resemble spartaeines closely in habitus, but this observation is insufficiently precise to provide evidence for relationship.
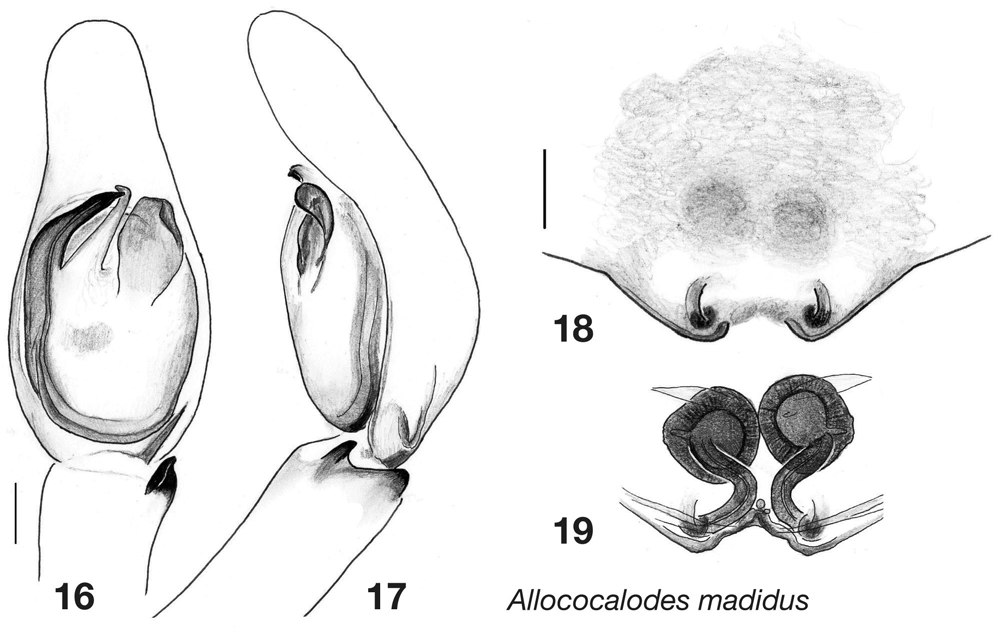
However, the three new genera described here are provisionally placed within the Cocalodinae along with Cocalodes and Allococalodes . This group has one proposed synapomorphy, the internal sclerotized spheres of the epigynum first reported by Wanless in Cocalodes . These are, however, also present in at least a few salticoids (see e.g., Galiano 1963, 1970: Tullgrenella morenensis (Tullgren) and Chira gounellei (Simon)) . These spheres are visible clearly in the new species Yamangalea frewana , Tabuina varirata , Tabuina baiteta , Tabuina rufa , and Cucudeta zabkai ( Figs 30 View FIGURES 20 – 30 , 43, 50 View FIGURES 37 – 50 , 61 View FIGURES 51 – 61 , 78 View FIGURES 72 – 86 ; see arrow in Fig. 43 View FIGURES 37 – 50 ), but were not seen in C. uzet and C. gahavisuka ( Figs 82, 86 View FIGURES 72 – 86 ) and are ambiguous in Allococalodes madidus ( Fig. 19 View FIGURES 16 – 19 ). Additional evidence for the monophyly of the cocalodines comes from preliminary data from the 28S gene, by which Cocalodes , Allococalodes and the three genera described here are resolved as a monophyletic group (Maddison & Zhang unpublished).



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |