
Odocoileus virginianus
|
publication ID |
https://doi.org/10.1206/335.1 |
|
persistent identifier |
https://treatment.plazi.org/id/4E0BBE15-FFDC-77C8-FF03-570293A6EB06 |
|
treatment provided by |
Tatiana |
|
scientific name |
Odocoileus virginianus |
| status |
|
Odocoileus virginianus View in CoL Figures 19 View Fig , 25–29 View Fig View Fig View Fig View Fig View Fig
In ventrolateral view (fig. 28), the pars cochlearis has a hemi-ellipsoid-shaped promontorium, which has one primary bulge for the cochlea (there is a subtle second one farther anteriorly but it is not nearly as pronounced). The fenestra cochleae is circular and larger than the fenestra vestibuli, which assumes a more oval shape. The two fenestrae are separated by a moderately wide crista interfenestralis. There are no transpromontorial sulci or sulci for the stapedial artery on the promontorium. The fossa for the tensor tympani is oval and slightly excavates the adjacent tegmen tympani. The promontorium tapers to a shelflike epitympanic wing anteriorly. This wing is fully continuous with a posteromedial flange, such that the two projections of bone form a shelf surrounding the promontorium both anteriorly and medially. On the posterior aspect of the posteromedial flange is an almost complete foramen that would appear to be positioned to transmit the internal carotid artery; however, this artery is allegedly absent in adult ruminants (Schummer et al., 1981). This foramen pierces the posteromedial flange. In this area, the posteromedial flange bends ventrally.
On the pars canalicularis, the tegmen tympani is inflated moderately and accounts for approximately one-third the width of the petrosal. The anterior process of the tegmen tympani is distinct and terminates in a blunt apex. Where the tegmen tympani meets the external acoustic meatus posteriorly there is a sharp, ventrolateral process that is spikeshaped. The external acoustic meatus is a shallow indistinct trough. The external acoustic meatus leads on to an epitympanic recess with a distinct, rounded pit. Immediately posterior to the fossa for the tensor tympani is an anteriorly positioned secondary facial foramen opening onto the facial sulcus. A thin sheet of bone covers the opening of the secondary facial foramen in ventral view, and the secondary facial foramen has a pointed apex anteriorly. The facial sulcus is a very distinct groove lateral to the stapedial muscle fossa. The tympanohyal is broken on this specimen but was attached to a thin, elongate projection at the end of the crista parotica. At the medial margin of the tympanohyal is the relatively distinct and open stylomastoid notch at the posterior end of the long facial sulcus. This taxon has a wide caudal tympanic process situated posterior and medial to the fenestra cochleae. It is smooth and curls ventrally very slightly at its medial edge. It is also fully continuous with
(Ruminantia, Cervidae ). Scale 5 1 cm.
(Ruminantia, Cervidae ). Scale 5 1 cm.
the posteromedial flange. The mastoid region is large (approximately equal to the length of the pars cochlearis) and wedge-shaped. The pars cochlearis protrudes medially relative to the mastoid region, such that the two meet at an angle of approximately 130 °.
There is extensive contact between the tympanic and the petrosal (fig. 19). The posterior crus of the meatal tube, which is relatively elongate, has an extensive and narrow contact with the mastoid region along a smooth ridge. The anterior crus of the meatal tube contacts the posterior surface of the ventrolateral process, a contact that is relatively small by comparison with the size of the contact for the posterior crus. The bulla itself contacts the anterior process of the tegmen tympani over most of its ventrolateral surface but does not fuse to it. The bulla also contacts most of the ventromedial surface of the petrosal including the epitympanic wing, the posteromedial flange, and the caudal tympanic process.
The dorsomedial surface (fig. 29) is smooth anteriorly and becomes cancellous toward the posterior end of the mastoid region. The internal acoustic meatus is relatively circular with a fold disrupting the dorsal margin. The crista transversa is a delicate ridge tucked deep within the internal acoustic meatus. The prefacial commissure is a broad and flat bar of bone that meets the adjacent tegmen tympani at a right angle. There is no prefacial commissure fossa. The epitympanic wing is visible in this view as a sharp point positioned midway between the ventral and dorsal margins of the bone. Extending posteriorly from the epitympanic wing is a subtle crest with a jagged, ruffled edge, which extends along the anteromedial border to just inferior to the internal acoustic meatus. This demarcated the basicapsular groove. Posterior to the groove is an indistinct, medially oriented triangular knob that is also seen in Antilocapra americana , and is much better developed in the latter taxon. Tucked deep to this protuberance is a small, slit-shaped cochlear aqueduct. Posterior to this knob is a small shelf of bone that covers a very small vestibular aqueduct. Posterior to the internal acoustic meatus, and separated from it by only a slender ridge, is a rectangular shallow depression, the subarcuate fossa. There is no petromastoid canal. Posterior to this is a second square depression with a small pit. The mastoid region tapers to a point posteriorly.
The dorsolateral surface (fig. 25) of the petrosal is flat and the tegmen tympani forms a right angle with the dorsomedial and ventrolateral surfaces. The tegmen tympani projects slightly over the tympanic surface, and its anterior process terminates in a blunt end. The ventrolateral tuberosity is a thin, projecting spike set adjacent to the tegmen tympani. There are no vascular grooves on the surface of the tegmen tympani. The wedgeshaped mastoid region is thickest toward the ventral surface where it forms a rounded knob, and it is thinnest toward the dorsal surface where it tapers to a thin sheet of bone.
The ventromedial pars cochlearis (fig. 26) widens posteriorly from the anterior tip. The smooth, flat anterior part ends about halfway along the length of the bone as a knob, which is an expanded caudal tympanic process just anterior to the mastoid region. There is a lip of bone along the pars cochlearis (made by the epitympanic wing, the posteromedial flange, and the caudal tympanic process) that projects toward the tympanic bulla, when articulated, and wraps onto its external surface. As noted above, a small foramen interrupts this lip of bone adjacent to the stylomastoid notch. The cochlear aqueduct is a small slit adjacent to this foramen. The mastoid region fans into a triangle with a rough, cancellous surface. An elongate mastoid exposure was present on the lateral surface of the skull. There is no mastoid plate.
In anterior view (fig. 27) the hiatus Fallopii is small and tucked into the moderately inflated anterior process of the tegmen tympani. The prefacial commissure fossa is absent, as noted above, with the area superi- or to the commissure being flat and at a right angle to the tegmen tympani.
ARTIODACTYLA – RUMINANTIA – PECORA – GIRAFFIDAE
GIRAFFA CAMELOPARDALIS Figures 25–27 View Fig View Fig View Fig , 30–32 View Fig View Fig View Fig
In ventrolateral view (fig. 30) the promontorium of the pars cochlearis is hemi-ellipsoid
(Ruminantia, Giraffidae ). Hatched area indicates the tympanohyal attachment site. Scale 5 1 cm.
(Ruminantia, Giraffidae ). Scale 5 1 cm.
with a ventral projection over the fenestra cochleae. The fenestra cochleae is relatively round and slightly larger than the more oval fenestra vestibuli. The promontorium has double transpromontorial sulci. A faint sulcus extends toward the fenestra vestibuli but it is so faint that this is not scored as a sulcus for the stapedial artery. There is a very shallow, oval fossa for the tensor tympani that does not excavate the tegmen tympani. A subtle ridge divides this fossa. The promontorium also gives rise to an epitym- panic wing that forms the anteriormost point on the petrosal. This is continuous both with the adjacent tegmen tympani and also medially with the posteromedial flange. A complete anterior and medial flange of bone rings the promontorium.
On the pars canalicularis the tegmen tympani and the mastoid region are both rough in contrast to the smooth promontorium. The tegmen tympani is inflated moderately and consists of approximately one-half to one-third the total size of the ventral surface of the promontorium. The anterior process of the tegmen tympani runs along the entire dorsolateral edge of the promontorium and terminates just at the anterior extreme of the promontorium without extending anterior to it. Posterior to the fossa for the tensor tympani is a large opening for the anteriorly positioned secondary facial foramen onto the facial sulcus. Dorsolateral to this is a ventrally displaced shelf of bone, the epitympanic recess. The area surrounding this recess is rugose with pits and raised bumps. There is one central depression in the epitympanic recess, but overall it is difficult to interpret the exact position for the fossa for the head of the malleus. The external acoustic meatus is a shallow, semicircular notch demarcated by a tall, thin, spike-shaped ventrolateral tuberosity anteriorly. The clear facial sulcus wraps lateral and posterior to an oval stapedial muscle fossa. The stylomastoid notch is positioned at the end of this groove, near the base of the attachment site for the tympanohyal. The caudal tympanic process is knob-shaped, hooked posteriorly, and offset by a deep groove at its posterior end. The caudal tympanic process also forms an extensive shelf posterior to the fenestra cochleae. In ventral view, only a small part of the mastoid region is visible; it is composed of rough, cancellous bone.
Both the anterior and posterior crura of the ectotympanic contacted the petrosal (fig. 31). The posterior crus contacted the mastoid region at a large, elongate ledge, and the anterior crus contacted the ventrolateral tuberosity of the petrosal over a small area. The anterior tympanic bulla contacted the anterior process of the tegmen tympani over a large, roughened area but did not fuse to it. The medial and posterior tympanic bulla contacted the medial and posterior parts of the pars cochlearis and the caudal tympanic process.
The dorsal surface (fig. 32) consists of a smooth anterior aspect around the internal acoustic meatus, which tapers into a cancellous posterior portion. The internal acoustic meatus has an elongate oval edge that is better defined on the posterior margin. The crista transversa is a broad division between the foramen acusticum superius and inferius. The prefacial commissure fossa is absent. A subtle, sharp-edged groove extends from the anterior extreme of the petrosal in endocranial view to the ventral margin where it ends inferior to the internal acoustic meatus. This marks the basicapsular groove, which is present largely on the dorsal surface extending only slightly onto the medial surface. The edge of this groove terminates near the cochlear aqueduct, which is relatively large and round. Posterodorsal to the internal acoustic meatus is a very shallow subarcuate fossa with no petromastoid canal. This is followed posteriorly by a second shallow pit that tapers into the cancellous, triangular mastoid region. The vestibular aqueduct is a slit distomedial to the subarcuate fossa.
Dorsolaterally (fig. 25), the moderately inflated tegmen tympani is predominantly flat and forms a right angle with the dorsomedial and tympanic surfaces. It is roughest toward the tympanic surface. The anterior process of the tegmen tympani terminates in a blunt tip. The tegmen tympani is characterized by one major (the vascular groove) and several minor grooves that cross from ventral to dorsal. Posterior to the tegmen tympani there is a smooth deep pit. The mastoid region has a fan shape and the bone is highly cancellous and distinguished by several vascular foramina. The ventrolateral process is a sharp, projecting spike.
The ventromedial margin (fig. 26) has two main divisions: the anterior part consisting of the pars cochlearis, which is triangular with a flat medial surface, and the pars canalicularis, which consists of the caudal tympanic process and the mastoid region, with the latter being a large, fan-shaped posterior portion. The basicapsular groove extends from the dorsal surface briefly onto the medial surface. The small cochlear aqueduct is a clear hole (not a slit) on the medial surface just anterior to a distinct knob. Ventrolateral to the knob is a foramen (noted above, fig. 26) that is tucked deep to the knoblike caudal tympanic process as it hooks posteriorly. The fan-shaped mastoid region is highly cancellous and extends dorsal and ventral to the promontorium. The posterior mastoid exposure was a rough surface, shaped as an elongate oval, not a triangle.
Anteriorly (fig. 27), the flatness of tegmen tympani is apparent. A small but distinct hiatus Fallopii is offset toward the ventrolateral surface and emerges from under a roughened anterior process of the tegmen tympani. The tegmen tympani does not extend substantially over the endocranial or the ventral surface. The prefacial commissure fossa is absent, and the dorsomedial and dorsoventral surfaces of the petrosal meet at a right angle.
ARTIODACTYLA – CAMELIDOMORPHA – CAMELIDAE
CAMELUS DROMEDARIUS Figures 31 View Fig , 33–37 View Fig View Fig View Fig View Fig View Fig
This bone was firmly sutured to the skull posteriorly and had to be cut at its posterior margin for removal. The bone overall is smooth.
In ventrolateral view, the pars cochlearis of the petrosal (fig. 33) has a hemi-ellipsoid promontorium with a gentle convexity anterior to the fenestra cochleae. The fenestra cochleae is relatively round and slightly open and irregular at its posterior border. The fenestra vestibuli is approximately the same size as the fenestra cochleae but is much more oval. A crista interfenestralis that is relatively broad separates the two fenestrae and extends posterior to the fenestra cochleae. There are no conspicuous transpromontorial or stapedial artery sulci on the promontorium. The promontorium tapers anteriorly into an epitympanic wing that forms the anteriormost part of the petrosal. This is fully continuous with the posteromedial flange and together these form a lip of bone with a jagged edge around the medial and anterior aspects of the promontorium. The fossa for the tensor tympani muscle is an elongate, inconspicuous oval that is very shallow and does not excavate the tegmen tympani at all. The posteromedial flange gives rise to a pyramidal-shaped rostral tympanic process that projects ventrally just anterior to the cochlear aqueduct on the medial side.
On the pars canalicularis, the tegmen tympani is moderately inflated, accounting for approximately one-third the width of the ventral surface, and it has a very subtle and blunt anterior process that merges closely with the epitympanic wing. Posterior to the fossa for the tensor tympani is a small pit. The secondary facial foramen is lateral and slightly posterior to the fenestra vestibuli. It is hidden very deeply within the bony covering that blocks from view much of the path of the facial sulcus to the stylomastoid notch. The stapedial muscle fossa is not visible due to the enclosed nature of this part of the petrosal. Due to the fusion of the bulla and petrosal it is also difficult to discern the exact position of the stylomastoid notch in the isolated petrosal (its approximate position is indicated). Lateral to the secondary facial foramen is a large, round, and shallow epitympanic recess, which, in this species, exhibits little differentiation for the fossa for the head of the malleus. The epitympanic recess merges laterally into the petrosal contribution to the external acoustic meatus, which is defined by tall borders both anteriorly and posteriorly. The tympanohyal is not preserved in this specimen but its hypothesized attachment site is indicated at the posterior end of the sinuous crista parotica. The caudal tympanic process is a wide ledge of bone posterior and medial to the fenestra cochleae. It is fully continuous with the posteromedial flange. The mastoid region is wedge-shaped and relatively large (although damaged some in its removal from the skull due to fusion). The pars cochlearis protrudes relative to the mastoid region with a sharp angle between them.
The ectotympanic meatal tube does not appear to contact the petrosal in many places (fig. 31). The tympanic bulla only contacts the petrosal at the posteromedial flange and rostral tympanic process areas. The rest of the bulla contacts the squamosal and the basioccipital.
(Camelidomorpha, Camelidae ). Scale 5 1 cm.
(Camelidomorpha, Camelidae ). Scale 5 1 cm.
In dorsolateral view, the internal acoustic meatus is the most prominent depression on the endocranial surface; it is roughly oval with a crease at its dorsal margin. The foramen acusticum superius and inferius are separated by a crista transversa that is narrow and sharp. From the anterior extreme of the bone extends a defined groove along the endocranial margin and extends to the vestibular aqueduct. This is the basicapsular groove and it occupied a dorsal position (see also Smuts and Bezuidenhout [1987: fig. 6.2, no. 20] who figured a substantial ‘‘sinus petrosus ventralis’’ on the dorsomedial surface of the petrosal). Posterior to the internal acoustic meatus is a very low ridge followed immediately by a shallow subarcuate fossa. Within the subarcuate fossa is a petromastoid canal. The mastoid portion of the bone continues posteriorly as a shallow concavity. The vestibular aqueduct emerges from deep within a slit at the medioventral margin and is covered dorsally by several sheets of bone. The cochlear aqueduct is positioned at the ventromedial edge of the promontorium inferior and posterior to the internal acoustic meatus.
Dorsolaterally (fig. 35), the tegmen tympani is largely flat and forms a right angle with the tympanic and endocranial surfaces. The anterior process of the tegmen tympani is small with a blunt tip. A vascular groove, set relatively close to the tympanic surface, runs anteromedially. The mastoid region is a thin, triangular wedge.
Ventromedially (fig. 36), the promontorium has a sharp edge between the dorsomedial and ventrolateral surfaces; there is no flattened ventromedial margin. At the distomedial edge of the promontorium the bone becomes increasingly square-shaped and then tapers to a flat plate. Posteriorly, that the mastoid region was exposed on the external surface of the skull can be confirmed in juvenile specimens (e.g., AMNH-M 80114), but this is often hard to confirm on adult specimens due to fusion of the petrosal to the squamosal and occipital bones. The mastoid plate is absent.
Anteriorly (fig. 37), the hiatus Fallopii is visible as a small slit deep to the anterior process of the tegmen tympani. Medial to it are two small foramina (possibly vascular). There is no prefacial commissure fossa and the tegmen tympani is relatively flat and perpendicular to the ventral and endocranial surfaces of the bone.
ARTIODACTYLA – CAMELIDOMORPHA
† POEBROTHERIUM SP. Figures 31 View Fig , 35–39 View Fig View Fig View Fig View Fig View Fig
The hemi-ellipsoid promontorium (fig. 38) of the pars cochlearis has two ovoid bulges, one directly anterior to the fenestra cochleae and one anterior to that. The fenestra cochleae is round and only slightly larger than the oval-shaped fenestra vestibuli; they are separated by a narrow crista interfenestralis that has distinct continuity with the caudal tympanic process. The promontorium is devoid of grooves. Anteriorly, the promontorium attenuates into a large epitympanic wing. This wing forms the anteriormost part of the petrosal and is continuous with the adjacent tegmen tympani. The promontorium also gives rise to a posteromedial flange. Just posterior to the posteromedial flange is a small groove on the medial surface of the petrosal. These projections from the promontorium form a flattened lip of bone that surrounds the anterior and medial sides of the promontorium. The fossa for the tensor tympani muscle is a shallow, oval area anterolateral to the fenestra vestibuli that does not excavate the adjacent tegmen tympani.
The tegmen tympani is moderately inflated and comprises approximately one-third the total width of the petrosal. The anterior process of the tegmen tympani ends bluntly and is separated from the rest of the petrosal medially by a small ridge. The secondary facial foramen emerges from the deep surface of the fossa for the tensor tympani and opens anterior to the fenestra vestibuli. The facial sulcus extends from the facial foramen, over a convexity, to the relatively open stylomastoid notch. The stapedial muscle fossa is a deep oval pit immediately anterior to the stylomastoid notch. The epitympanic recess is lateral to the facial sulcus, offset ventrally, and is separated from the facial sulcus by a clear crista parotica. The epitympanic recess
(Camelidomorpha, Camelidae ). Scale 5 1 cm.
(Camelidomorpha, Camelidae ). Scale 5 1 cm.
does not exhibit particularly distinctive fossa for the head of the malleus. The external acoustic meatus is bordered by a knobshaped ventrolateral tuberosity. The mastoid region is a square-shaped, gnarled knob that is thickest toward the ventral surface. Posterior to the fenestra cochleae is a large and wide caudal tympanic process that is medial to the stapedial muscle fossa. The pars cochlearis does not protrude ventromedially relative to the mastoid region.
The ectotympanic (fig. 31) contacts the petrosal in several places. The anterior bulla contacted the anterior process of the tegmen tympani but did not fuse to it. The anterior and posterior crura of the ectotympanic did not contact the ventrolateral tuberosity or the mastoid region, respectively. The medial and posterior pars cochlearis had extensive contact with the tympanic bulla.
The dorsomedial surface is entirely smooth (fig. 39). The internal acoustic meatus is oval, with a well-defined edge except posteriorly. Superior to the internal acoustic meatus is a small knob; there is no prefacial commissure fossa (see also fig. 37). The foramina acusticum superius and inferius are separated by a thin crista transversa. There are no significant grooves on the anterior or medial margins of the promontorium, and there is no basicapsular groove; instead, the ventromedial surface of the petrosal forms a sharp edge. Posterodorsal to the internal acoustic meatus is a large and very deep subarcuate fossa. Its posterodorsal edge is relatively undefined and is capped by a curved groove. Ventromedial to the subarcuate fossa is a small knob that ends in a posterior lip, deep to which is the vestibular aqueduct. The cochlear aqueduct is a minor slit ventromedial to the internal acoustic meatus. Posterior to the subarcuate fossa, the mastoid region is smooth and flat and terminates in a point posteriorly. The mastoid comprises only about one-fourth the total size of the petrosal.
Dorsolaterally (fig. 35), the tegmen tympani is more flat (see also fig. 37) than convex but has several bumps and a generally uneven surface. The hiatus Fallopii is hard to discern in this specimen (see also fig. 37). There are no vascular grooves. The mastoid region assumes a knoblike shape, not a wedge.
The ventromedial surface (fig. 36) of the promontorium has a sharp edge anteriorly that widens into a flat area at the posterior part of the promontorium, and it ends in a smooth knob. The slitlike cochlear aqueduct is ventrally offset and is positioned about midway between the anterior and posterior ends of the bone. The edge of the open stylomastoid notch can be seen in this view as well. A groove demarcates an abrupt transition between the caudal tympanic process and the more distal, knob-shaped mastoid region. The small squarish posterior end of the mastoid is exposed on the external surface of the skull, and the mastoid plate is absent.
ARTIODACTYLA – † PROTOCERATIDAE
† PROTOCERAS CELER Figures 31 View Fig , 35–37 View Fig View Fig View Fig , 40–41 View Fig View Fig
The ventrolateral surface (fig. 40) has a hemi-ellipsoid promontorium, dropping off toward the anterior and medial edges. There is a subtle transpromontorial groove visible only at the anterodorsal margin where it terminates at a large notch. The sulcus for the stapedial artery is absent. The fenestra cochleae is larger than the fenestra vestibuli; the former is circular, and the latter is oval. The two fenestrae are separated by a wide crista interfenestralis. The fossa for the tensor tympani is an extensive, circular depression that excavates the tegmen tympani. The epitympanic wing and the posteromedial flange both protrude from the promontorium; however, neither forms a large flat wing of bone.
On the pars canalicularis, the tegmen tympani is moderately inflated and occupies one-fourth the width of the ventral surface. Its anterior process terminates as a knob, is separated from the promontorium, and does not extend anterior to it. The ventral opening of the secondary facial foramen is just posterior to the fossa for the tensor tympani and immediately lateral to the fenestra vestibuli. Posterior to the secondary facial foramen, the facial sulcus passed over a ventrally protruding convexity and curved lateral to the stylomastoid foramen. The fossa for the stapedial muscle is a large, deep
( † Protoceratidae ). Scale 5 1 cm. Damaged area is hatched.
( † Protoceratidae ). Scale 5 1 cm.
pit situated posterior to both the fenestra vestibuli and the fenestra cochleae. The epitympanic recess occupies a ventrally displaced platform and does not have a distinct fossa for the head of the malleus or a fossa incudis. The external acoustic meatus has a small, knob-shaped ventrolateral tuberosity at its anterior margin. The tympanohyal is not preserved but may have attached on a ridge posterior to the stylomastoid notch. A clear crista parotica extends from that area and separates the epitympanic recess from the adjacent facial sulcus. The caudal tympanic process was a wide ledge of bone, posterior and posteromedial to the fenestra cochleae. The pars cochlearis does not protrude relative to the mastoid region. The mastoid region is large and wedge-shaped.
There do not appear to be any contacts between the tympanic and the petrosal (fig. 31).
The dorsomedial surface (fig. 41) is divided almost evenly between the pars cochlearis and the mastoid region. It is entirely smooth. The internal acoustic meatus is a teardrop-shaped structure that tapers at the dorsal edge and is relatively open ventrally. It has a broad crista transversa. Anterior and dorsal to the internal acoustic meatus is a semicircular depression. There is no prefacial commissure fossa. There are no pronounced ridges on the endocranial surface except at the medial margin, where there is a gentle, elongate depression, the basicapsular groove. Posterior to the internal acoustic meatus is a flat, rectangular depression that runs dorsoventrally and abuts the shallow subarcuate fossa at its anterior margin. At the center of the subarcuate fossa is a petromastoid canal. Inferior to the subarcuate fossa is a small knob on a dorsoventrally oriented ridge. The vestibular aqueduct is tucked deep to a small shelf of bone on the mastoid region. The mastoid region terminates posteriorly as a point.
Dorsolaterally (fig. 35), the tegmen tympani is relatively flat with several isolated bumps and grooves. The hiatus Fallopii is a small hole at the anterior margin of the tegmen tympani. A vascular groove runs anteroposteriorly along the lateral surface from the base of the ventrolateral tuberosity. The mastoid region is bent strongly toward the dorsomedial side of the bone.
Ventromedially (fig. 36), the bone is narrow anteriorly with a sharp dorsomedial edge extending the length of the pars cochlearis. The basicapsular groove is a very subtle feature, slightly inset on the endocranial surface. The cochlear aqueduct is a small slit on the medial surface. The mastoid region is roughened on the ventromedial surface and has a triangular outline. Overall the shape of the mastoid region is a wedge. A flat, triangular portion of the mastoid was exposed on the external surface of the skull. There was no mastoid plate.
In anterior view (fig. 37) the promontorium can be seen to have been hemi-ellipsoid, in contrast to the much rounder hemispherical shape of many cetaceans. The hiatus Fallopii is a relatively small hole, and there is no prefacial commissure fossa. The tegmen tympani is essentially flat and meets the dorsomedial side of the petrosal at a right angle.
ARTIODACTYLA – CETANCODONTA – HIPPOPOTAMIDAE
CHOEROPSIS LIBERIENSIS Figures 42–47 View Fig View Fig View Fig View Fig View Fig View Fig
The ventrolateral surface of the petrosal (fig. 42) has an overall shape of a rounded triangle with the base of the triangle at the posterior end. The promontorium is hemiellipsoid and has two subtle convexities: one immediately anterior to the fenestra cochleae, and one medial to the fossa for the tensor tympani muscle. The fenestra cochleae is irregular in outline: the anterior aspect of its margin has a well-defined semicircular edge, and the posterior one-third is poorly defined without a clear edge. The fenestra vestibuli, by contrast, has a more typical oval shape and is smaller than the fenestra cochleae. The two fenestrae are separated by a relatively narrow crista interfenestralis. There are no clear sulci on the promontorium. Anterodorsally is an expansive but shallow fossa for the tensor tympani with an irregular-shaped border. It excavates, very slightly, the adjacent tegmen tympani. The promontorium gives rise to both an epitympanic wing and a posteromedial facet. These two relatively flat projections of bone form a continuous shelf
(Cetancodonta, Hippopotamidae ). Scale 5 1 cm.
(Cetancodonta, Hippopotamidae ). Scale 5 1 cm.
around the promontorium and contact the caudal tympanic process posteriorly. A distinct and roughened basicapsular groove runs anteroposteriorly along most of the promontorium; thus, this structure is in the ventral position.
On the pars canalicularis, the entire lateral margin of the tympanic surface consists of a rough-textured, irregular, hyperinflated tegmen tympani that extends to the anterior edge of the promontorium. The anterior process of the tegmen tympani terminates in a blunt knob. Posterior to the tegmen tympani is a large, concave depression with tall anterior and posterior margins that is the petrosal contribution to the external acoustic meatus. The anterior projection is a knobshaped ventrolateral tuberosity. The epitympanic recess is positioned slightly anterior to the external acoustic meatus. It is an indistinct depression that is contiguous with the fossa for the tensor tympani; it has no large fossa for the head of the malleus as seen in cetaceans. The secondary facial foramen emerges deep to the fossa for the tensor tympani through a small slit. This opening is lateral to the fenestra vestibuli and thus is relatively anterior in position. The short facial sulcus is adjacent to a stapedial muscle fossa that is relatively shallow and poorly defined. A hypothesized tympanohyal attachment is labeled at the posterior end of the crista petrosa; however, the tympanohyal is not preserved in this specimen (if present). The stylomastoid notch is a narrow slit. The caudal tympanic process is a large, wide flange of bone posterior to the fenestra cochleae. The mastoid region is a short, irregular knob. The pars cochlearis does not protrude medially relative to the mastoid region.
The ectotympanic has extensive contact with the petrosal (fig. 43). The anterior crus of the ectotympanic contacts the ventrolateral tuberosity, and the posterior crus of the ectotympanic contacts the mastoid region over a relatively large area. The anterior tympanic bulla contacts the tegmen tympani but does not fuse to it. The medial and posterior parts of the bulla have extensive contacts along the ventromedial edge of the pars cochlearis and pars canalicularis. The medial and posterior contacts are inset slightly from the ventromedial edge of the petrosal.
The smooth dorsomedial surface of the petrosal (fig. 44) of this species starts at a point anteriorly and widens posteriorly. The only large opening on the endocranial surface is the internal acoustic meatus, which has a poorly defined outline shape, being sharpest at the dorsal edge. The foramina acusticum superius and inferius are wide holes that are separated by a narrow crista transversa. Dorsal to the internal acoustic meatus is the prefacial commissure, which is a flattened ridge that expands dorsolaterally into a large convexity, the prefacial commissure fossa. The prefacial commissure fossa itself is smooth and abuts the lateral roughened border of the hyperinflated tegmen tympani. A crest defines the edge between dorsomedial and dorsolateral sides. In dorsomedial view, the tegmen tympani is essentially continuous with the mastoid region (in ventral view the division between these is marked by the external acoustic meatus). The subarcuate fossa is absent; dorsolateral and posterior to the internal acoustic meatus is a petromastoid canal. The mastoid region terminates posteriorly as a point. The mastoid region is relatively small, and on the dorsomedial surface it is gently convex, forming a conspicuous swelling posterior to the internal acoustic meatus. The cochlear aqueduct occupies a ventromedial position.
The hyperinflated tegmen tympani can be best viewed dorsolaterally (fig. 45). The surface has a rough texture with a gentle, anteroposteriorly running vascular groove. Several very small vascular foramina dot this surface. The ventrolateral tuberosity is a large knoblike protrusion. The ventromedial (fig. 46) side of the promontorium exposes the elongate basicapsular groove. This groove lies ventrolateral, rather than dorsomedial, to a sharp edge that delineates these two surfaces of the bone. At the posterior end of this groove is a slit that is the cochlear aqueduct. The mastoid plate is absent.
In anterior view (fig. 47) the hiatus Fallopii is a distinct circular foramen opening onto the semirectangular anterior surface of the tegmen tympani. The hiatus is located in the deepest point of a shallow recess. The hyperinflated tegmen tympani occupies most of the lateral surface of the bone and has a convex shape. The deeply excavated prefacial commissure fossa is also clear in anterior view.
ARTIODACTYLA – CETANCODONTA – HIPPOPOTAMIDAE
HIPPOPOTAMUS AMPHIBIUS Figures 43 View Fig , 45–49 View Fig View Fig View Fig View Fig View Fig
The pars cochlearis visible in ventrolateral view (fig. 48) has a large, hemi-ellipsoid promontorium with two gently ovoid areas, one anterior to the fenestra cochleae and one anteromedial to the fenestra vestibuli. The fenestra cochleae has an irregular outline that is distinctly oval anteriorly but widens into an undefined posterior edge. It is larger than the fenestra vestibuli, which is oval, and is directed ventrally. The crista interfenestralis forms a pronounced lateral margin to the fenestra cochleae and becomes a ridge that extends to the posterior edge of the promontorium blending with the caudal tympanic process. The promontorium has very subtle double transpromontorial sulci. These extend in parallel from the medial edge of the promontorium, anterior to the fenestra cochleae, across the medial edge of the anterior ovoid bump, and they terminate at the anterior edge of the promontorium. Tucked medial to the tegmen tympani, and extending deep within it, is a large fossa for the tensor tympani muscle (the thin bony roof of the fossa is slightly broken in this specimen). There is a pit that is fully separate from the fossa for the tensor tympanic and is situated anteromedial to the fenestra vestibuli. The sulcus for the stapedial artery is absent. A conspicuous epitympanic wing extends from the anterior aspect of the promontorium and is contiguous with the tegmen tympani. The epitympanic wing is not flat but instead bends dorsomedially. The posteromedial flange is also present and fully continuous with the epitympanic wing to complete a shelf around the anterior and medial aspects of the promontorium. The basicapsular groove runs along the ventromedial edge of the pars cochlearis, rather than in a dorsal position.
The pars canalicularis comprises a substantial portion of the petrosal. Lateral to the promontorium is a hyperinflated tegmen tympani that occupies half the total width of the promontorium in ventral view. The anterior process of the tegmen tympani is thick and somewhat enlarged but does not extend anterior to the anterior edge of the pars cochlearis. The anterior process ends in a blunt point. The fossa for the tensor tympani, described above, lies adjacent to a large, deep epitympanic recess (and some of the epitympanic recess is out of view due to fused ectotympanic). The epitympanic recess lies lateral to the anteriorly positioned secondary facial foramen. From this the facial sulcus loops lateral to a deep stapedial muscle fossa. The contribution from the petrosal to the external acoustic meatus is a distinctive elongate trough, and this specimen retains part of the ectotympanic fused to the external acoustic meatus. Anterior to the external acoustic meatus is the ventrolateral tuberosity, which is a substantial, pointed spike of bone. It is not clear what remained of the tympanohyal (which was likely small) in this specimen, but I have estimated a possible attachment site. The crista parotica is present but hard to see due to the fusion of parts of the ectoympanic to the petrosal. The caudal tympanic process is a very wide (medially extensive) shelf posterior to the fenestra cochleae. It is fully continguous with the posteromedial flange. It has a dorsoventral ridge on its medial aspect. The mastoid region (the area posterior to the external acoustic meatus) is a wide knob-shaped bone that was not exposed on the external surface of the skull. The mastoid region is scored as ‘‘small’’ because even though it is large and bulbous looking, it only amounts to half the size of the pars cochlearis.
There is extensive contact between the ectotympanic and the petrosal (fig. 43). The anterior crus of the ectotympanic has extensive contact with the posterior surface of the ventrolateral tuberosity. The posterior crus of the ectotympanic also contacts a large portion of the mastoid region and fuses to it. Also, as noted above, part of the tympanic contribution to the external acoustic meatus is fused to the petrosal part of the external acoustic meatus. The anterior tympanic bulla has a small contact with the anterior process of the tegmen tympani and is fused to it. The
(Cetancodonta, Hippopotamidae ). Scale 5 1 cm.
medial edge of the bulla contacts the pars cochlearis and the caudal tympanic process along their medial margins.
The dorsomedial surface (fig. 49) is smooth. The internal acoustic meatus is very large, being a widely open hole that takes up much of the medial surface of the pars cochlearis. It does not have a well-defined outline. Posterior to it is an ovoid swelling marked by a ridge on the posterior surface. The foramen acusticum superius and foramen acusticum inferius are separated by a broad crista transversa. There are no conspicuous grooves on the endocranial surface. The prefacial commissure is a distinct ridge and posterior to it is an extensive excavation, the prefacial commissure fossa, on the endocranial surface of the hyperinflated tegmen tympani. The prefacial commissure fossa is separated from the internal acoustic meatus by a pronounced bar of bone. At the posterior aspect of the prefacial commissure fossa is a petromastoid canal, a small, narrow, and deep hole. Posterior to it is an irregular, convex tuberosity of bone. Endocranially, the transition between the tegmen tympani and the mastoid region is not very clear because the two areas are continuous. At the posterior end of the mastoid region there is small pit. The vestibular aqueduct is tucked deeply within a series of holes and knobs at the posterior end of the promontorium. It does not emerge directly onto the endocranial surface. The cochlear aqueduct emerges at the posteriormost part of the promontorium such that it occupies a relatively ventral position.
The dorsolateral view (fig. 45) exposes the strongly inflated tegmen tympani, which has a texture that is knobby, grooved, and pocked but not cancellous; the bone has a square outline in this view. This side of the bone exhibits a large vascular groove and a number of smaller, irregular grooves. One prominent groove anteriorly winds onto the tympanic surface and runs the entire length of the bone from anterior to posterior. The ventrolateral tuberosity is a pronounced, pointed pyramidal structure. The mastoid region is not particularly enlarged relative to the rest of the petrosal and is rounded in shape.
In ventromedial view (fig. 46) a sharp edge of the pars cochlearis separates dorsomedial and tympanic sides of the bone. The basicapsular groove can be seen to lie on the tympanic surface rather than on the dorsomedial surface. The promontorium ends posteriorly in a series of three knobs. The knob-shaped mastoid region extends dorsal and ventral to the promontorium in this view and has a long crease at its anterior end. No part of the bone is exposed on the external surface of the skull, and the mastoid plate is absent.
In anterior view (fig. 47) the very large hiatus Fallopii is visible on the flattened, anterior surface of the tegmen tympani. On the specimen, the foramen acusticum superius is situated extremely close to the hiatus Fallopii, such that one can see from one foramen into the other. The anterior view shows the hyperinflated condition of the tegmen tympani, which has a convex surface, and the highly excavated prefacial commissure fossa.
ARTIODACTYLA – SUINA – SUIDAE
Figures 43 View Fig , 45–47 View Fig View Fig View Fig , 50–51 View Fig View Fig
The ventrolateral surface (fig. 50) consists of a relatively small, hemi-ellipsoid-shaped promontorium that has two ovoid bulges, with one anterior to the fenestra cochleae and another anterior to that. The fenestra cochleae is approximately twice the size of the fenestra vestibuli. The crista interfenestralis is a narrow ridge that extends from the lateral rim of the fenestra cochleae and runs medially to join the caudal tympanic process at the posterior aspect of the promontorium. Extending from the promontorium anteriorly is a large epitympanic wing, with a rugose texture at its anterior extreme, which contacts the adjacent tegmen tympani. Continuous with the epitympanic wing medially is a substantial posteromedial flange. Together these comprise a larger part of the pars cochlearis than does the promontorium. The fossa for the tensor tympani muscle is a very elongate, somewhat oval area that excavates the adjacent tegmen tympani. A large and wide groove runs from the ventromedial margin of the pars cochlearis to the fenestra
(Suina, Suidae ). Scale 5 1 cm.
(Suina, Suidae ). Scale 5 1 cm.
cochleae. The transpromontorial sulcus and the sulcus for the stapedial artery are, however, absent.
On the pars canalicularis, the tegmen tympani is moderately inflated, occupying about one-fourth the total width of the petrosal. The anterior process of the tegmen tympani terminates anteriorly as a scroll of bone that comes to a point. The secondary facial foramen opened anterior to the fenestra vestibuli onto a facial sulcus that followed a clear groove to the stylomastoid notch. The fossa for the stapedial muscle is relatively shallow. The epitympanic recess is a small area that lacks distinct impressions for the ossicles; in fact, it is difficult to identify a clear epitympanic recess at all. The external acoustic meatus is a very shallow channel with no adjacent ventrolateral tuberosity. This specimen did not preserve evidence regarding the tympanohyal, but the possible site of its attachment is indicated at the posterior end of the sharp crista parotica. The caudal tympanic process is very broad in this taxon, extending medially from a position posterior to the fenestra cochleae. At its medial extreme it is shaped as a large, ventrally projecting knob. The mastoid region is relatively small and has a swollen, knob-shaped dorsolateral margin. The pars cochlearis does not protrude medially relative to the mastoid region.
There are several contacts between the petrosal and the ectotympanic (fig. 43). The contacts are small between both the anterior and posterior crura of the ectotympanic (which is laterally extended into a long tube in this taxon) and the petrosal. The tympanic bulla also contacts the anterior process of the tegmen tympani but is not fused to it. Both the medial and posterior aspects of the bulla contacted the pars cochlearis in one extensive contact.
The entire dorsomedial (fig. 51) surface of the petrosal is smooth. It is divided almost evenly between the pars canalicularis and the pars cochlearis. The internal acoustic meatus is a large oval area with a clearly defined posterosuperior border and a more poorly defined anteroinferior border. Inferior to it there is a small knob. The wide crista transversa puts a clear separation between the foramina acusticum superius and inferius. There is no prefacial commissure fossa. Anteriorly, the spike-shaped anterior extreme of the anterior process of the tegmen tympani is visible in this view. There is a very faint jagged ridge on the medial surface indicating the position of the basicapsular groove. The cochlear aqueduct is positioned relatively ventrally. Posterior to the internal acoustic meatus is a shallow subarcuate fossa with a petromastoid canal at its center. In endocranial view the mastoid plate blocks a view of other parts of the mastoid region.
The dorsolateral (fig. 45) surface is relatively flat but slopes toward the endocranial surface anteriorly. There are no conspicuous vascular grooves. The tegmen tympani is moderately inflated and its anterior process comes to a point anteriorly. The very large hiatus Fallopii is out of view.
Ventromedially (fig. 46), the promontorium has a flattened surface. The basicapsular groove is a subtle feature on the anterior half of the ventromedial promontorium. The cochlear aqueduct is a tiny slit. The caudal tympanic process is a robust, triangular structure just anterior to the stylomastoid notch. The knoblike mastoid region is distinct relative to the mastoid plate and has a large vascular foramen. Posteriorly, the mastoid region is divided into a mastoid plate endocranially and a distinctly separate, knob-shaped mastoid region ventrally. There is no mastoid exposure on the external surface of the skull.
Anteriorly (fig. 47), there is a very large hiatus Fallopii that connects directly to the foramen acusticum superius in the internal acoustic meatus (i.e., one can see through both of these foramina simultaneously as in Hippopotamus amphibius ). Absence of a prefacial commissure fossa is also apparent, as is the flat and only moderately inflated tegmen tympani, which meets the endocranial surface of the petrosal at a right angle.
ARTIODACTYLA – SUINA – SUIDAE
SUS SCROFA Figures 43 View Fig , 52–56 View Fig View Fig View Fig View Fig View Fig
The anatomy of the petrosal, ear ossicles, and tympanic of this taxon were described by
1 cm.
Getty (1975) in some detail but not figured. A detailed description and illustrations of the tympanic surface of the petrosal can be found in Parker (1874: XXXVI–XXXVII), and that description was an important source for characters used here. Finally, in comparisons with artiodactylans, Luo and Gingerich (1999) also figured the petrosal of this taxon.
On the ventrolateral surface (fig. 52) the hemi-ellipsoid promontorium has two pronounced ovoid areas, one directly anterior to the fenestra cochleae and one anterolateral to it. The fenestra cochleae is much larger than the fenestra vestibuli, and its outline shape is circular anteriorly and straight posteriorly. The fenestra vestibuli is oval. The crista interfenestralis was very narrow, and the two fenestrae are positioned relatively close to each other. There are no transpromontorial or stapedial artery sulci, but there is a strong groove starting at the posteromedial aspect of the pars cochlearis that extends posterior to the fenestra cochleae and toward the fenestra vestibuli. The promontorium gives rise to two substantial flat extensions of bone, the epitympanic wing anteriorly and the posteromedial flange posteromedially. Together these form a broad, continuous platform of bone. The epitympanic wing is one of the anteriormost points of the bone and merges with the tegmen tympani laterally. The fossa for the tensor tympani is oval and excavates the adjacent tegmen tympani; however, Getty (1975) stated that this muscle is not well developed in the pig.
The tegmen tympani of the pars canalicularis is moderately inflated, occupying approximately one-fourth the size of the petrosal. Its anterior process becomes a thin sheet of bone that ends in a point. The anterior process also partially laps onto the ventral surface of the promontorium and epitympanic wing. The secondary facial foramen is relatively posterior in position (i.e., it is fully posterior to the fenestra cochleae), such that it opens adjacent to the stapedial muscle fossa. The facial sulcus is thus relatively short as it winds lateral to the stapedial muscle fossa. The epitympanic recess is an indistinct area with no distinctly separate fossae for ossicles. The petrosal contribution to the external acoustic meatus is unremarkable and shallow without clear landmarks anteriorly or posteriorly; the ventrolateral tuberosity is absent. The attachment site for the tympanohyal (structure not preserved in this specimen) was likely at the end of the crista parotica just lateral to the stylomastoid notch. The crista parotica is a sharp crest extending from the lateral side of the secondary facial foramen to the site of attachment of the tympanohyal. The caudal tympanic process is a wide, square area of bone posterior to the fenestra cochleae and extending medial to it. It is continuous with the crista interfenestralis, and the groove noted above travels across it. The mastoid region is a small, rounded knob; the pars cochlearis does not protrude medially relative to the mastoid region.
There are several areas of contact between the bulla and the petrosal (fig. 43). The anterior crus of the ectotympanic contacts the petrosal over a relatively small area, and the anterior tympanic bulla contacts a large area on the anterior process of the tegmen tympani but does not fuse to it. Antero- and posteromedially there are contacts between the pars cochlearis and the tympanic bulla. Finally, there is a small contact between the posterior crus of the ectotympanic and the mastoid region of the petrosal.
The smooth dorsomedial surface of the petrosal (fig. 53) is divided evenly between the pars cochlearis and the pars canalicularis. The internal acoustic meatus is a large, rectangular opening with a clearly defined posterior border and a more open, undefined anterior border. The foramen acusticum superius is separated from the foramen acusticum inferius by a narrow crista transversa. There is no prefacial commissure fossa, just a thin prefacial commissure superior to the internal acoustic meatus. Dorsal and posterior to the internal acoustic meatus is a small knob. The basicapsular groove appears to be present on the medial aspect of the endocranial surface but is extremely subtle, suggested only by a slight ruffled margin. At the dorsolateral edge of the petrosal is a very shallow subarcuate fossa distinguished by a curved crease rather than having the characteristic rounded shape. The petromastoid canal is present. The mastoid plate dominates the dorsomedial surface of the mastoid. The cochlear aqueduct is a thin, oval slit that is directly medioinferior to the internal acoustic meatus. Posterior to this is a small shelf of bone that covers the vestibular aqueduct.
Dorsolaterally (fig. 54), the tegmen tympani is relatively flat but has several knobs and small creases. It is moderately inflated with a small, pointed anterior process. The ventrolateral tuberosity is absent, and there are no vascular grooves. The hiatus Fallopii is very large and meets the foramen acusticum superius almost immediately (one can see between the two foramina). In this view the ventral part of the mastoid region can be clearly seen to be a knob, but medially there is a mastoid plate that is the most posteriorly extensive part of the bone.
The ventromedial surface (fig. 55) is flat with a triangular outline. The basicapsular groove is a very subtle structure on the dorsomedial surface, barely sculpting the petrosal at all. The ventrally positioned cochlear aqueduct is visible in this view. The extent of the caudal tympanic process as a prominent knob-shaped structure can also be seen. At the posterior margin there is a groove between the mastoid region and the mastoid plate. The mastoid region has a number of vascular foramina; no mastoid exposure existed on the external surface of the skull. From the endocranial surface of the petrosal a mastoid plate protrudes as the most posterior structure.
In anterior view (fig. 56) the very large hiatus Fallopii is visible. This view also shows how the tegmen tympani meets the endocranial surface of the bone at a right angle.
ARTIODACTYLA – SUINA – SUIDAE
HYLOCHOERUS MEINERTZHAGENI Figs. 54–59 View Fig View Fig View Fig View Fig View Fig View Fig
On the ventrolateral surface (fig. 57) the pars cochlearis has a hemi-ellipsoid promontorium. The promontorium has two pronounced convexities: one anterior to the fenestra cochleae and one anteromedial to the fossa for the tensor tympani. Traversing the promontorium is a single transpromontorial sulcus. The fenestra vestibuli is much smaller than the fenestra cochleae and has an oval-shaped rim. The fenestra cochleae has a rim with a generally circular outline that is irregular in places. The two fenestrae are separated by a narrow and sinuous crista transversa. Medial to the tegmen tympani is a shallow, oval, and wide depression, the fossa for the tensor tympani. This fossa excavates the surrounding tegmen tympani. The promontorium gives rise to a very large epitympanic wing at the anteromedial aspect of the bone. This wing ends in a point anteriorly, which is the most anterior aspect of the bone. The bone also has a posteromedial flange that is fully continuous with the epitympanic wing, such that together they form a broad shelf of bone around the promontorium.
The pars canalicularis has a moderately inflated tegmen tympani, which occupies about one-fourth the total size of the bone. Its anterior process is small, with a blunt anterior apex. The epitympanic recess is small and nondescript; it does not have distinct fossae for the ossicles. The petrosal contribution to the external acoustic meatus is present but does not form a clear channel or groove. Posterior to the fossa for the tensor tympani is the secondary facial foramen. This foramen occupies a relatively posterior position near the stapedial muscle fossa, being posterolateral to the fenestra vestibuli rather than anterolateral to it. The facial sulcus is distinct but short due to the posterior position of the secondary facial foramen. The stapedial muscle fossa is shallow and vague except for its medial margin, which is defined by a sharp crest. At the medial extreme of this fossa is the caudal tympanic process, which is a wide extension of bone posterior and medial to the fenestra cochleae that also has a small, ventrally projecting spike. The mastoid region is smooth and ends in a knob; the pars cochlearis does not protrude medially relative to the mastoid region. There is a mastoid plate (see also fig. 55) that forms the posteriormost part of the mastoid region.
There are several areas of contact between the tympanic and the petrosal (fig. 58). From the preparation of this particular specimen it is not completely clear whether the crura of the ectotympanic contacted the petrosal. It is clear, however, that the anterior part of the tympanic bulla contacted the anterior process
53672) (Suina, Suidae ). Scale 5 1 cm.
53672). Scale 5 1 cm.
of the tegmen tympani but did not fuse to it. Furthermore, the medial parts of the bulla contacted the entire medial and posterior edge of the pars cochlearis.
On the dorsomedial surface (fig. 59) the bone is completely smooth. The internal acoustic meatus is a gently recessed oval with a strong lip only at the posterior margin; the anterior margin is relatively open. The foraminae acusticum superius and inferius are separated by a substantial crista transversa. The prefacial commissure is a flat ridge of bone, and there is no prefacial commissure fossa. The dorsal surface of the epitympanic wing forms the anteriormost aspect of the petrosal in this view. Extending posteriorly from this is the basicapsular groove. This groove gives a flaked, irregular contour to the anterior part of the dorsomedial surface. The groove ends inferior to the internal acoustic meatus adjacent to the slitlike cochlear aqueduct, with the latter occupying a relatively ventral position. The pars canalicularis of the endocranial surface is distinguished by a shallow subarcuate fossa. At the center of this fossa is a pin-sized opening of the petromastoid canal. The area adjacent to this is entirely smooth, lacking other tubercles or processes. A mastoid plate extends posteriorly from the subarcuate fossa and is the prominent part of the mastoid region in the dorsomedial view.
The dorsolateral surface of the petrosal (fig. 54) is flat with an overall triangular outline that tapers anteriorly. At approximately the midpoint between anterior and posterior is the small hiatus Fallopii, which is positioned fully on the dorsal surface. The hiatus Fallopii is slightly recessed. The tegmen tympani is moderately inflated with a blunt anterior process. There are no vascular grooves on the dorsal surface, and there is a small ventrolateral tuberosity that is spikeshaped. The knob-shaped mastoid region is clearly visible in this view.
Ventromedially (fig. 55), the surface of the bone is also flat, widening posteriorly from a narrow anterior extreme. The basicapsular groove is positioned closest toward the dorsomedial surface. The small, recessed cochlear aqueduct is positioned about halfway from the anterior tip of the bone. Posteriorly, there is a mastoid plate adjacent to a knob-shaped mastoid region. The plate is oriented at an angle and is the posteriormost part of the pars cochlearis. There is no mastoid exposure on the external surface of the skull in this taxon.
Anteriorly (fig. 56), the relatively small hiatus Fallopii is positioned slightly toward the ventrolateral surface. The lack of prefacial commissure fossa and the moderate inflation of the tegmen tympani are also visible. The dorsal surface of the petrosal extends into the endocranial cavity (medially) to a small degree.
ARTIODACTYLA – SUINA – SUIDAE
BABYROUSA BABYRUSSA Figures 54–56 View Fig View Fig View Fig , 58 View Fig , 60–61 View Fig View Fig
The ventrolateral surface (fig. 60) of the petrosal is piriform in outline, and the promontorium is hemi-ellipsoid. The promontorium has a bulbous central surface with the most inflated region immediately anterior to the fenestra cochleae. The fenestra cochleae is relatively large with a circular rim that has a poorly defined posterior edge, in contrast to the relatively small and oval fenestra vestibuli. The two fenestrae are separated by a narrow and sinuous crista interfenestralis. A transpromontorial sulcus is conspicuous just anterior to the fenestra cochleae; this is the only clear sulcus on the promontorium. A roughly oval-shaped (but also irregular in outline) fossa for the tensor tympani muscle is present and excavates the adjacent tegmen tympani. The promontorium gives rise to a large epitympanic wing that comes to a point anteriorly and is the anteriormost aspect of the petrosal. The epitympanic wing is fully continuous with the posteromedial flange and together the two form a flattened ring of bone around the anterior and medial promontorium. The basicapsular groove is present on the ventrolateral (rather than dorsomedial) surface.
On the pars cochlearis, the tegmen tympani is moderately inflated, occupying approximately one-fourth the total size of the bone. It is a raised bump with a very inconspicuous anterior process that ends in a small, blunt area. A slit that appears not to
(Suina, Suidae ). Scale 5 1 cm.
(Suina, Suidae ). Scale 5 1 cm. be the primary hiatus Fallopii (the latter is not visible in ventrolateral view) is present at the anterior margin of the tegmen tympani. Fully posterior to this, and just lateral to the fenestra vestibuli, is the secondary facial foramen, which is dorsolateral to the fenestra vestibuli and therefore anterior in position. Lateral to this is a shelf of bone that is offset ventrally. This smooth area is the epitympanic recess. It does not show distinct impressions for ossicles. The petrosal contribution to the external acoustic meatus is a shallow, flat area that is rather unremarkable relative to the rest of the ventrolateral surface. The facial sulcus emerges lateral to the shallow, oval stapedial muscle fossa. The stylomastoid notch is indistinct, situated adjacent to the hypothesized attachment site for the tympanohyal. From this attachment site extended the crista parotica anteriorly and medial to the epitympanic recess. The caudal tympanic process is extensive both mediolaterally and anteroposteriorly. It is generally smooth. The mastoid region is a diminutive knob that has a distinct (?vascular) foramen. The pars cochlearis does not protrude medially relative to the mastoid region. The mastoid region is irregular in shape and is more posteriorly extensive on the dorsomedial surface where there is a mastoid plate.
There are several areas of contact between the ectotympanic and the petrosal (fig. 58). The anterior and posterior crura of the ectotympanic (which are expanded into a meatal tube in this taxon) contact the ventrolateral tuberosity and the mastoid region, respectively. The anterior tympanic bulla has extensive contact with the anterior process of the tegmen tympani but does not fuse to it. The medial and posterior parts of the bulla contact the entire ventromedial margin of the petrosal in one continuous area.
Dorsomedially (fig. 61), the petrosal has a piriform outline. The internal acoustic meatus is a broad, open oval with a poorly defined anterior border. It is surrounded by smooth bone, and the fenestrae acusticum superius and inferius are distinctly separated by a broad crista transversa. Immediately posterior to the internal acoustic meatus is a distinct ridge. There is no prefacial commissure fossa. On the dorsolateral edge (fig. 54) is the relatively large hiatus Fallopii in profile; at its posterior edge is a knob. As noted above, there is no distinct basicapsular groove on the dorsomedial surface; instead, it is positioned ventrolaterally. The pars canalicularis has a deep subarcuate fossa with a sharp ringlike edge that is particularly well defined posteriorly. The posterior edge of the fossa is characterized by small notches. The cochlear aqueduct is an inconspicuous hole situated ventromedially; the vestibular aqueduct is largely out of view, tucked under a ledge of bone. There is a small petromastoid canal within the subarcuate fossa. The mastoid plate is also visible in this view as a distinct endocranial structure separate from the rest of the mastoid region.
Dorsolaterally (fig. 54), the bone is flat with no distinct vascular grooves. The tegmen tympani is moderately inflated with a small, blunt anterior process. The ventrolateral tuberosity is a small knob, and the large hiatus Fallopii is visible at an angle. Ventromedially (fig. 55), the bone has a triangular shape with the apex at the anterior margin; it widens toward the posterior end. The cochlear aqueduct is situated approximately two-thirds the distance from the anterior end of the bone. The mastoid region ends as a small knob, and endocranially adjacent to it is a mastoid plate. No part of the bone is exposed on the external surface of the skull.
Anteriorly (fig. 56), the very large hiatus Fallopii is visible, offset slightly toward the ventrolateral surface. There is no prefacial commissure fossa, and the moderately inflated tegmen tympanic is flat, forming a perpendicular edge to the dorsomedial (endocranial) surface.
ARTIODACTYLA – SUINA – TAYASSUIDAE
TAYASSU TAJACU Figures 58 View Fig , 62–66 View Fig View Fig View Fig View Fig View Fig
On the ventrolateral surface (fig. 62) the promontorium of the pars cochlearis has two distinct convexities, one anteromedial to the fenestra cochleae and one anterolateral to it. The fenestra cochleae is approximately double the area of the fenestra vestibuli. The fenestra cochleae is circular anteriorly with a more widely opened, irregular rim posteri- orly. The fenestra vestibuli is oval. The crista interfenestralis is a narrow ridge separating the two fenestrae. A single transpromontorial sulcus is apparent, wrapping anterior to the fenestra cochleae and running to the anterodorsal edge of the promontorium. There is no sulcus for the stapedial artery branching from the transpromontorial sulcus toward the fenestra vestibuli. Extending from the promontorium anteriorly is a shelflike epitympanic wing that thins to a point, forming an anterior apex of the promontorium. This wing expands medially to join a separate shelf, the posteromedial flange, and the site of their union is characterized by a series of faint ridges. Together they form a flat and continuous lip on the anterior and medial sides of the pars cochlearis. At the medial margin of this lip is an indication of the basicapsular groove, which is partly visible although primarily positioned on the dorsomedial surface of the petrosal. Tucked deep within its ventral surface of the petrosal is a large fossa for the tensor tympani, an elongate, generally oval trough that tapers to a point anteriorly. The fossa is so extensive that it excavates part of the adjacent tegmen tympani.
On the pars cochlearis, the tegmen tympani is inflated and occupies approximately one-fourth the ventral surface of the petrosal. The anterior process is blunt and does not extend anterior to the promontorium. Just posterior to the fossa for the tensor tympani, on a ventrally displaced shelf of bone lateral to the secondary facial foramen, is the epitympanic recess. This is a poorly defined area with no pronounced separate fossa for the head of the malleus as often seen in cetaceans. The petrosal contribution to the external acoustic meatus is flanked anteriorly and posteriorly by two small bumps; the anterior of these is the ventrolateral tuberosity, which is spike-shaped. The facial sulcus is a distinct channel winding lateral to the stapedial muscle fossa. The small, shallow, and oval stapedial muscle fossa lies immediately posterior to the fenestra cochleae. The stylomastoid notch is a narrow opening that has a small spicule of bone just anterior to it. The tympanohyal was attached just posterior to the stapedial muscle fossa. From its attachment site extended a distinct crista parotica. On the ventromedial surface is a raised, triangular wedge of bone, the caudal tympanic process, which extends broadly across the posterior edge of the fenestra cochleae and has a knob at its medial edge. Posterior to the facial sulcus, the mastoid region drops off to form a flat, gently slanted knob.
The ectotympanic contacted the petrosal in several places (fig. 58). Both the anterior and posterior crura of the ectotympanic (expanded into a tube in this taxon) contacted the external acoustic meatus portion of the petrosal rather extensively, although the exact area of contact is approximated in this specimen. The anterior tympanic bulla contacted the anterior process of the tegmen tympani over much of its ventromedial margin. The medial and posterior aspects of the bulla contacted the medial and posterior parts of the petrosal at its medial edge; these contacts were not continuous.
The entire dorsomedial petrosal surface (fig. 63) is smooth. This surface has one large opening, a transversely teardrop-shaped internal acoustic meatus that comes to a point dorsally where there is a small crease. Anteriorly, the extensive epitympanic wing is visible in endocranial view. A narrow crista transversa separates the foramina acusticum superius and inferius. There is no prefacial commissure fossa; superior to the internal acoustic meatus is a small knob. The basicapsular groove begins anterior to the internal acoustic meatus at the anterior margin of the bone. As the groove extends posteromedially, it winds briefly onto the tympanic surface of the bone. Posterior to the internal acoustic meatus is a shallow, rectangular-shaped depression that is followed immediately by a shallow subarcuate fossa. There are no significant features within the subarcuate fossa; the petromastoid canal is absent (a microscopic hole is present on the bone, but I do not consider this a large enough structure to be a petromastoid canal). The vestibular aqueduct is just ventromedial to the subarcuate fossa, emerging as a small hole deep to a shelf of bone. A short, flattened mastoid plate comprises the posterior end of this surface (fig. 63, drawing).
Dorsolaterally (fig. 64), the tegmen tympani is both moderately inflated and relatively flat. It has a small hiatus Fallopii positioned almost exactly between the ventromedial and dorsolateral surfaces. The posterior part of the lateral surface has a small, sunken foramen in its center and several small foramina adjacent to it. The mastoid region is a small knob with a mastoid plate adjacent to it. There is a small, spike-shaped ventrolateral tuberosity.
Ventromedially (fig. 65), the bone is triangular with a very small cochlear aqueduct just at the posterior end of the basicapsular groove. There are several small foramina on this surface that appear to open into the basicapsular groove. This side exposes a triangular, robust, and projecting caudal tympanic process. The cochlear aqueduct is hard to distinguish but was most likely one of several very small holes on the ventromedial margin. The mastoid plate is offset endocranially from the knob-shaped mastoid region. No part of the bone was exposed on the external surface of the skull.
Anteriorly (fig. 66), the small but distinct size of the hiatus Fallopii is clear, being positioned at the anterior margin of the dorsal surface of the tegmen tympani. The moderately inflated tegmen tympani forms a right angle with the endocranial surface of the bone. This view also reveals the lack of prefacial commissure fossa.
ARTIODACTYLA – † ANTHRACOTHERIOIDEA – † ANTHRACOTHERIIDAE
† ELOMERYX ARMATUS Figures 58 View Fig , 67–71 View Fig View Fig View Fig View Fig View Fig
The specimen is extremely well preserved with no broken areas. The right and left sides from the same individual have been illustrated in ventral view, one with (photograph) and one without (drawing) the tympanohyal.
The ventrolateral surface (fig. 67) has a hemi-ellipsoid shape with two bulges on the promontorium, one immediately anterior to the fenestra cochleae and a second anterior and dorsal to that. The round fenestra cochleae is larger than the oval fenestra vestibuli. The crista interfenestralis is a narrow crest and the two fenestrae are relatively close together. Double transpromontorial sulci extend from the ventromedial edge of the promontorium and pass anterior to the fenestra cochleae (fig. 67), where they are particularly conspicuous. They then turn laterally toward the more anterior bulge on the promontorium. There is no sulcus for the stapedial artery branching from the transpromontorial sulcus. The promontorium then tapers anteriorly to form an epitympanic wing that comes to a point anteriorly. The epitympanic wing is continuous with the posteromedial flange, a second, flattened projection off the promontorium. These two continuous flanges are notable for having several shallow depressions. Tucked medioventral to the tegmen tympani is a large fossa for the tensor tympani muscle, a fossa that excavates the adjacent tegmen tympani.
The pars cochlearis consists of a moderately inflated tegmen tympani and a relatively short mastoid region. In ventrolateral view (fig. 67) the tegmen tympani occupies about one-fifth the width of the petrosal. It has a blunt-ended anterior process that does not extend anterior to the promontorium. The secondary facial foramen (fig. 67, photograph) opens just posterior to the fossa for the tensor tympani muscle and lateral to the fenestra vestibuli, thus occupying a relatively anterior position. It feeds onto the facial sulcus and lies medial and deep to the epitympanic recess. The epitympanic recess is a nondescript shelf that is ventrally offset relative to the facial sulcus. The external acoustic meatus is a low, shallow trough characterized by a particularly tall anterior border. Demarcating the medial edge of the epitympanic recess is the crista petrosa, which winds posteriorly to the base of the tympanohyal. It has postmortem damage
( † Anthracotherioidea, † Anthracotheriidae ). Scale 5 1 cm.
(AMNH-VP LUSK 0781572), and † Archaeotherium sp. (AMNH-VP 96433). Ventrolateral surface is gray.
that has crushed it against the petrosal (fig. 67, photograph); however, its shape is entirely intact. It is an elongate rod with a slightly widened tip that has a small pit. The stylomastoid notch is a small concavity medial to the attachment site for the tympanohyal. The caudal tympanic process is a wide and smooth shelf posterior and posteromedial to the fenestra cochleae. A deep, oval stapedial muscle fossa is present with the facial sulcus running lateral to it. The mastoid region is a small knob, and the pars cochlearis does not protrude medially relative to the mastoid region.
The ventrolateral surface (fig. 58) has a number of contacts with the ectotympanic. The anterior and posterior crura of the ectotympanic (expanded into a tube in this taxon) both contact the petrosal at the external acoustic meatus over a continuous area. On one of the specimens illustrated (fig. 67, photograph), part of the meatal tube is fused to the external acoustic meatus in the region where the anterior and posterior crura would have attached. The posterior contact is small and the bone is smooth in that area. The anterior tympanic bulla contacts the anterior process of the petrosal over an extensive area. The medial and posterior tympanic bulla also contacted the medial edge of the petrosal over an extensive and continuous area.
The entire dorsomedial surface (fig. 68) is smooth. The pars cochlearis comes to a point anteriorly; this is the dorsal surface of the epitympanic wing. The internal acoustic meatus has a gently rectangular outline and immediately posterior to it is a very shallow, rectangular depression (long axis oriented dorsoventrally). The oval-shaped foramen acusticum superius and foramen acusticum inferius are separated by a narrow crista transversa. The anteroventral margin of the petrosal has an inset basicapsular groove that is positioned fully on the dorsal or endocranial surface of the bone. This groove extends posteriorly to the inferior border of the internal acoustic meatus. Superior to the internal acoustic meatus is the prefacial commissure, which is an elongate ridge. There is no prefacial commissure fossa. The moderately inflated tegmen tympani meets the endocranial surface of the bone at a right angle. Just ventral to the end of the basicapsular groove is a small cochlear aqueduct. Posterior to this and slightly dorsal is a very shallow subarcuate fossa. This species lacks a petromastoid canal. At the caudoventral margin is an elongate slit with a hood of bone over its opening in endocranial view. This is the vestibular aqueduct. The mastoid region is small, smooth, and featureless, ending in a squared posterior margin.
The dorsolateral surface (fig. 69) is triangular in outline with a conspicuous hiatus Fallopii at the anterior margin of the anterior process of the tegmen tympani. This surface is relatively flat, although it has some knobs. The ventrolateral tuberosity is knob-shaped and relatively indistinct in lateral view. There are no vascular grooves.
In ventromedial view (fig. 70) the bone is triangular in outline, widening posteriorly to a thickened knob. The minute cochlear aqueduct is offset slightly toward the endocranial surface but clearly holds a ventromedial position. The basicapsular groove is positioned distinctly on the dorsomedial surface. The mastoid region ends as a knob; there is no mastoid plate. The posterior mastoid region is thick and square-shaped, with minor foramina (possibly vascular). No exposure of the mastoid region existed on the external surface of the skull.
The anterior view (fig. 71) reveals the flat dorsal surface of the tegmen tympani and the subtle dorsal protrusion of the petrosal into the cranial cavity. The hiatus Fallopii is a distinct and relatively large opening at the anterior tegmen tympani. This view also reveals that the dorsolateral and dorsomedial surfaces are essentially perpendicular and there is no prefacial commissure fossa.
ARTIODACTYLA – † MERYCOIDODONTIDAE
† MERYCOIDODON SP. Figures 69–74 View Fig View Fig View Fig View Fig View Fig View Fig
The specimen is relatively well preserved except for the damage to the fenestra vestibuli and the deep part of the fossa for the tensor tympani.
On the ventrolateral surface (fig. 72) the hemi-ellipsoid promontorium bulges anterior to the fenestra cochleae and then flattens into
0781572) ( † Oreodontoidea, † Oreodontidae ). Scale 5 1 cm.
0781572) ( † Oreodontoidea, † Oreondontidae). Illustration (above), photograph (below). Scale 5 1 cm.
an epitympanic wing and a posteromedial flange. There are no transpromontorial or stapedial sulci on the promontorium. The fenestra cochleae is circular anteriorly with a straight posterior border. The fenestra vestibuli shape cannot be clearly discerned due to damage. The two fenestrae are separated by a narrow crista interfenestralis. A large, oval fossa for the tensor tympani excavates the tegmen tympani of the pars canalicularis.
On the pars canalicularis, the tegmen tympani is moderately inflated (e.g., fig. 71); it does not extend anterior to any part of the promontorium and terminates in a blunt anterior process. Lateral to the secondary facial foramen is the epitympanic recess, characterized by one depression with no clear separate fossae for the ossicles. The external acoustic meatus is a shallow, partly rough excavation. Immediately posterior to the fossa for the tensor tympani is the opening of the secondary facial foramen, which is anterior in position. The facial sulcus continues from this opening medially to the relatively narrow stylomastoid notch. The stapedial muscle fossa is difficult to discern, but it appears to be a deep, oval depression posteromedial to the fenestra vestibuli. Posteromedial to the fenestra cochleae is a wide caudal tympanic process. The attachment site for the tympanohyal is just posterior to the stapedial muscle fossa. Extending from it is the crista parotica, which forms a distinct border between the epitympanic recess and deeper structures such as the secondary facial foramen. The mastoid region is large and wedge-shaped, and it is about the same length as the pars cochlearis. The pars cochlearis protrudes medially relative to the mastoid region (the two meet at an angle of approximately 130 °). The stylomastoid notch is a tight area between the caudal tympanic process and the posterior end of the crista parotica.
There is significant contact between the tympanic and the petrosal (fig. 73), and these reconstructions have been supplemented by information from AMNH Lusk-0-84-1623. Both the anterior and posterior crura of the ectotympanic contacted the petrosal over a contiguous area at the external acoustic meatus. The anterior tympanic bulla contacted the anterior process of the tegmen tympani. The medial tympanic bulla contacted the medial and posterior petrosal extensively in one continuous contact.
The anterior two-thirds of the dorsomedial surface (fig. 74) is smooth. At the anterior extreme is an apex, which is the epitympanic wing in dorsal view. The internal acoustic meatus has a poorly defined oval border, and the foramina acusticum superius and inferius are separated by a crista transversa that is relatively wide. Superior to the internal acoustic meatus there is no prefacial com- missure fossa (see also fig. 71). The area is instead flat and abuts the tegmen tympani at a right angle. A basicapsular groove on the inferomedial surface extends from the anterior extreme of the bone and terminates inferior to the internal acoustic meatus. Just posterior to it is a small slit, the cochlear aqueduct, which is relatively ventral in position. Posterior to the internal acoustic meatus is a low, wide ridge that separates it from the subarcuate fossa. The subarcuate fossa is shallow and wide and has a petromastoid canal at its center. Directly inferomedial to the petromastoid canal is the vestibular aqueduct, a small slit under a ledge of bone and situated relatively close to the cochlear aqueduct. Posteriorly, the endocranial surface becomes roughened toward the posterior termination of the mastoid region. The mastoid region ends in a point.
Dorsolaterally (fig. 69), the petrosal is very smooth with a gently cylindrical anteroposterior convexity. There are no pronounced or continuous vascular grooves on this surface, only two pits at the posterior margin. The ventrolateral tuberosity is absent and the hiatus Fallopii appears only as a small slit in profile on the anterior aspect of the dorsal surface.
Ventromedially (fig. 70), the most conspicuous structures are the basicapsular groove, which is positioned dorsally, and the slit for the ventrally positioned cochlear aqueduct. Posteriorly, the bone widens into a triangle that is the large, rough, fan-shaped mastoid region. This area is abruptly separated from the anterior part of the petrosal. The part of the mastoid region that was exposed on the external surface of the skull is a tall, narrow strut of bone that represents the terminus of a fan-shaped mastoid region. This elongate mastoid region exposure had a smooth external surface; there is no mastoid plate. Dorsolaterally, the absence of any vascular grooves on the tegmen tympani is apparent, as is the relatively flat shape of the tegmen tympani (which meets the endocranial margin of the bone at a right angle).
Anteriorly (fig. 71), the relatively small but distinct hiatus Fallopii is deep to a small ledge of bone at the anterior extreme of the tegmen tympani.
ARTIODACTYLA – † ENTELODONTIDAE
† ARCHAEOTHERIUM SP. Figures 69–71 View Fig View Fig View Fig , 73 View Fig , 75–76 View Fig View Fig
The specimen has suffered some damage; however, because petrosals of this taxon have not previously been described it is difficult to know the extent of the damage. The specimen is not fully preserved at its anteromedial margin. The mastoid region is also not fully preserved.
The ventrolateral surface (fig. 75) has a clear convexity anterior to the fenestra cochleae. The promontorium is hemi-ellipsoid in shape. The promontorium tapers anteriorly into an epitympanic wing, which is broken medially. This wing appears to have been continuous with the posteromedial flange, which is present more posteriorly. There are no transpromontorial or stapedial artery sulci on the promontorium. The crista interfenestralis is relatively narrow, and the fenestra cochleae is relatively large and oval in outline shape. The fossa for the tensor tympani muscle is a shallow, oval pit that very slightly excavates the tegmen tympani laterally.
Laterally, the pars cochlearis is well developed. The tegmen tympani is moderately inflated (figs. 69, 71) and does not extend anterior to the promontorium. Its anterior process terminates in a subtle, blunt end. The external acoustic meatus is a shallow, inconspicuous trough. The epitympanic recess is poorly defined and preserved with no remarkable pits for ossicles; it is offset ventrally relative to the facial sulcus. The secondary facial foramen is not preserved, but the facial sulcus is clear, as is the fossa for the stapedial muscle, which is an elongate, oval depression. The crista parotica demarcates a clear boundary between the facial sulcus and the end of several ventrally offset structures such as the epitympanic recess. The tympanohyal is not preserved in this specimen, but its inferred site of attachment at the posterior edge of the crista parotica is indicated. The caudal tympanic process is a broad shelf of bone extending well posterior to the fenestra cochleae and medial to the stapedial muscle fossa. The mastoid region is an irregular, somewhat jagged stump but may not be fully preserved in this specimen. It was partially fused to the surrounding bone in life. In life it appears that the pars cochlearis protruded medial relative to the mastoid region.
There was extensive contact between the petrosal and the ectotympanic (fig. 73). In intact specimens the tympanic is often tightly fused to the petrosal, even in subadults. Because of this fusion, separation of the bulla and petrosal is destructive, and the area covered by contacts could only be estimated here. Both the anterior and posterior crura of the ectotympanic contacted the external acoustic meatus of the petrosal. Anteriorly, the tympanic bulla contacted the anterior process of the petrosal over an extensive area. Medially and posteriorly, the bulla contacted the ventromedial margin of the petrosal over a large area.
The dorsomedial surface (fig. 76) is smooth. The anterior margin of the specimen ends anteriorly in an apex (part of the epitympanic wing) with some damage posteromedially (making the sharpness even more pronounced). The basicapsular groove curves around the ventromedial margin adjacent to the internal acoustic meatus. Separate from this is the cochlear aqueduct, which has a slit shape and is positioned relatively ventromedial. Dorsal and posterior to the internal acoustic meatus is a ridge that forms the prefacial commissure. Immediately dorsal to this, the bone remains convex, indicating that there is no prefacial commissure fossa. The dorsomedial surface abuts the tegmen tympani at roughly a right angle. Posterior to the internal acoustic meatus is a broad excavation, the subarcuate fossa, which is wide and very shallow. There is no petromastoid canal. A small vestibular aqueduct emerges posterior to a pronounced ledge of bone ventromedial to the subarcuate fossa. The mastoid region is knoblike but may not be fully preserved.
The dorsolateral side of the petrosal (fig. 69) is dominated by the moderately inflated tegmen tympani, which has a roughened surface. There are no conspicuous vascular grooves and there is no ventrolateral tuberosity. Ventromedially (fig. 70), there is a pointed anterior apex and the bone widens posteriorly. The cochlear aqueduct is a slit
( † Entelodontoidea, † Entelodontidae ). Scale 5 1 cm.
that is distinct from the basicapsular groove. Posterior to this is a pit that terminates in a stumplike mastoid region. The mastoid plate appears to have been absent.
Anteriorly (fig. 71), there is a relatively large hiatus Fallopii. This is a distinct opening dorsal to the pars cochlearis and within the inflated tegmen tympani.
ARTIODACTYLA – CETACEA – MYSTICETI – BALAENOPTERIDAE
BALAENOPTERA ACUTOROSTRATA Figures 73 View Fig , 77–80 View Fig View Fig View Fig View Fig
The derived condition of this bone makes it difficult to demarcate ventromedial and dorsomedial sides, which are very clear in other artiodactylans.
In ventrolateral view (fig. 77) the petrosal has a boomerang shape. The promontorium is smooth only in its center and becomes rough and jagged at its medial and lateral borders. The promontorium is hemispherical and lacks an epitympanic wing and a posteromedial flange. There are no transpromontorial or stapedial artery sulci. The fenestra vestibuli is oriented laterally and is well separated from the fenestra cochleae by a wide crista interfenestralis. The fenestra vestibuli is an oval, kidney-shaped hole that is smaller than the fenestra cochleae. The fenestra cochleae is oval with a slightly irregular posterior margin; it is oriented caudally. There is no distinct fossa for the tensor tympani.
The pars canalicularis accounts for most of the size of the petrosal due to both a relatively large tegmen tympani and mastoid region. Anterolateral to the promontorium is a very large, cancellous, and jagged area of bone that is triangular in outline shape. This is the enormous anterior process of the tegmen tympani that extends well anterior to the promontorium. The tegmen tympani overall is hyperinflated and accounts for almost half the size of the petrosal. Lateral to the proximal part of the facial sulcus is the circular fossa for the head of the malleus, which is flanked posteriorly by a narrow, mediolaterally oriented groove that terminates in a very small depression that may be the fossa incudis. Posterior to this is the petrosal contribution to the external acoustic meatus. There is no ventrolateral tuberosity marking the anterior border of the external acoustic meatus. The secondary facial foramen, which occupies a relatively anterior position, is situated just anterolateral to the fenestra vestibuli. A clear, elongate facial sulcus extends lateral to the deep stapedial muscle fossa. The caudal tympanic process is a narrow and rough area of bone posterior to the fenestra cochleae. The caudal tympanic process nearly abuts the medial margin of the crista parotica and the hypothesized tympanohyal attachment site (although it is not known whether a tympanohyal was present). The close proximity of these structures makes it difficult to see the stapedial muscle fossa. The mastoid region is a very elongate column of thick bone that accounts for 50% of the total size of the petrosal. The pars cochlearis protrudes medially relative to the mastoid. The pars cochlearis protrudes medially relative to the mastoid region; the two structures meet at approximately 90 ° to each other.
The ventrolateral surface (fig. 73) had several articulations with the ectotympanic. The anterior crus of the ectotympanic did not contact the petrosal; however, the posterior crus did at two small areas, one near the external acoustic meatus and one on the mastoid region. The anterior tympanic bulla contacted the anterior process of the tegmen tympani but the medial and posterior bulla had no contacts with the petrosal.
In dorsomedial view (fig. 78) the surface of the bone is rough and jagged, particularly surrounding the internal acoustic meatus. The foramen acusticum superius and foramen acusticum inferius are situated deep within the internal acoustic meatus and are separated by an indistinct crista transversa. Posterior to the internal acoustic meatus are the cochlear aqueduct and vestibular aqueduct, both of which are positioned largely on the endocranial surface. There is no prefacial commissure fossa. There is also no clear basicapsular groove. The tegmen tympani does not meet the endocranial surface at a right angle as in many noncetacean artiodactylans. The contact is instead gently convex. There is no subarcuate fossa or petromastoid canal. The mastoid region is largely convex in
504952) ( Cetacea, Mysticeti ). Scale 5 1 cm.
this view, although it is also highly irregular in shape and hard to characterize.
Ventromedially (fig. 80), several structures are very apparent. These include the hyperinflated and pointed anterior process of the tegmen tympani. Posterior to this, the hemispherical shape of the promontorium and the lack of basicapsular groove are apparent. The cochlear aqueduct is not visible in this view because it is positioned on the endocranial surface. There was no mastoid plate. Dorsolaterally (fig. 79), there were no vascular grooves on the petrosal.
Anteriorly (fig. 81), the hiatus Fallopii may have been one of the very small slits within the large area of cancellous bone that characterizes the dorsal surface of the petrosal. However, there is no clear hiatus Fallopii resembling that of noncetacean artiodactylans. In this view the convexity of the tegmen tympani extends dorsal to the endocranial surface of the petrosal. It is hyperinflated and, as noted above, the prefacial commissure fossa is absent because the area is instead convex.
ARTIODACTYLA – CETACEA – ODONTOCETI – INIIDAE
INIA GEOFFRENSIS Figures 73 View Fig , 79––83 View Fig View Fig View Fig View Fig View Fig
The ventrolateral (fig. 82) pars cochlearis is dominated by the smooth promontorium, which lacks transpromontorial or stapedial artery sulci. The promontorium is hemispherical and also lacks an epitympanic wing and posteromedial flange. The fenestra cochleae has an open distomedial margin that extends onto a groove. The fenestra vestibuli is oval and is positioned almost directly anterior to the fenestra cochleae. The crista interfenestralis is very wide and extends posteriorly. There is no conspicuous fossa for the tensor tympani muscle.
On the pars cochlearis, the hyperinflated tegmen tympani accounts for approximately one-third the width of ventral surface. The anterior process of the tegmen tympani is relatively large, extending anterior to the promontorium and terminating as a point. The epitympanic recess has a distinct and large fossa for the head of the malleus anterior to the small fossa incudis. The external acoustic meatus is a gentle trough, and anterior to it is a small, knob-shaped ventrolateral tuberosity. Medial to this is the distinct opening for the secondary facial foramen, which lies immediately lateral to the fenestra vestibuli, thus relatively anteriorly. This opening continues as a distinct facial sulcus that empties onto a wide stylomastoid notch at the posteromedial aspect of the mastoid region. There is no distinct outline distinguishing the borders of the stapedial muscle fossa from the facial sulcus (similar to the petrosal of the odontocete † Waipatia , figured by Fordyce [1994: fig. 11C]). A potential attachment site for the tympanohyal may have been just posterolateral to the stylomastoid notch, although it is not known whether this species has a tympanohyal. Extending from this area along a crest medial to the epitympanic recess is the crista parotica. The caudal tympanic process is a shelf of bone posterior to the fenestra cochleae and medial to the stylomastoid notch. It is narrow and smooth. The stylomastoid notch is tucked between the caudal tympanic process and the mastoid region. The mastoid region is small; its shape is ventrally flat and dorsally convex.
There is contact between both the anterior and posterior crura of the ectotympanic and the petrosal (fig. 73). The anterior crus contact is relatively small. The contact between the posterior crus and the mastoid region was large and rough in texture. The anterior tympanic bulla contacted the anterior process of the tegmen tympani and was fused to it. There were no contacts between the medial or posterior parts of the bulla and the medial petrosal. The pars cochlearis protrudes medially relative to the mastoid region.
The dorsomedial (fig. 83) surface is generally smooth with slight roughening on the anterior process of the tegmen tympani. The pars cochlearis is bulbous. The internal acoustic meatus has a loosely defined edge that approximates a pyriform shape and tapers anteriorly. Figure 83 View Fig shows the distinct foramen acusticum superius and inferius along with the foramen singulare. The crista transversa creates a broad separation be- tween the foramina acusticum superius and inferius. The basicapsular groove is absent. The pars cochlearis lacks a prefacial commissure fossa. Posterior to the internal acoustic meatus is the vestibular aqueduct and the cochlear aqueduct, both on the endocranial surface, and thus the cochlear aqueduct occupies a relatively dorsal posi- tion. The tegmen tympani is hyperinflated and convex. There is no subarcuate fossa and no petromastoid canal. The mastoid region occupies about one-fourth the size of the bone.
Dorsolaterally (fig. 79), the bone is convex overall. A knob-shaped ventrolateral tuberosity is apparent, as is the lack of vascular groove. The convex and hyperinflated tegmen tympani is particularly clear in this view. In ventromedial (fig. 80) view the promontorium is hemispherical with no conspicuous basicapsular groove. The cochlear aqueduct is dorsally positioned, making it relatively close to the internal acoustic meatus. The mastoid region is small with a convex dorsal border and a flat ventral border in this view.
In anterior view (fig. 81) there is no distinct anterior hole indicating a hiatus Fallopii as in noncetacean artiodactylans. There are several small holes on the dorsal surface of the tegmen tympani but their homologies are unclear. This view also demonstrates the large size and the overall convex shape of the tegmen tympani and the lack of prefacial commissure fossa.
ARTIODACTYLA – CETACEA – ODONTOCETI – KOGIIDAE
KOGIA BREVICEPS Figures 79–81 View Fig View Fig View Fig , 84–86 View Fig View Fig View Fig
The petrosal of Kogia was figured by Kasuya (1973).
On the ventrolateral (fig. 84) surface of the petrosal the full promontorium is hemispherical with no epitympanic wing, no posteromedial flange, and no transpromontorial or stapedial artery sulci. The fenestra vestibuli is round, not oval, and is larger than the fenestra cochleae. The fenestra cochleae is oval in outline anteriorly and becomes slightly irregular toward its posterior margin. There is a subtle, separate convexity anterior to the fenestra cochleae. The crista interfenestralis is relatively wide and consists of a bulging ventral ridge. The fossa for the tensor tympani is absent as any kind of distinct trough.
The pars canalicularis consists of a hyperinflated tegmen tympani with a large anterior process that extends anterior to the promontorium and terminates as a point. Posterior and slightly medial to the anterior process is a very large fossa for the head of the malleus, as well as the much smaller fossa incudis posterolateral to it. Together these make up the epitympanic recess. Anteromedial to the fossa for the head of the malleus is a small, pointed spike. The external acoustic meatus appears to be relatively narrow, and offset anterior to it is a ventrolateral tuberosity that is knob-shaped. The crista parotica is a sharp ridge extending along the medial surface of the external acoustic meatus and the fossa incudis. Posterior to the fossa for the head of the malleus and dorsolateral to the fenestra vestibuli is the secondary facial foramen, which occupies a relatively anterior position. It opens onto a facial sulcus that continues as a distinct channel winding lateral to the area for the stapedial muscle fossa, which is very poorly defined. The caudal tympanic process is broad and forms a distinct rounded ledge of bone posterior to the fenestra cochleae and the crista interfenestralis. A tentatively identified landmark for the attachment of the tympanohyal (if present) was most likely adjacent to the stylomastoid notch. The stylomastoid notch is relatively wide and open (see also fig. 80). The promontorium protrudes medially relative to the mastoid region (the pars cochlearis and pars canalicularis meet at an angle of approximately 140 ° on the medial side). The mastoid region is large and in ventrolateral view its surface is relatively flat.
Contact between the petrosal and the tympanic occurred in several places (fig. 85). While there is no contact between the anterior crus of the tympanic ring and the petrosal, there is a substantial contact between the posterior crus of tympanic ring and the mastoid region. The posterior crus of the tympanic ring has become a large, flattened plate that contacts half of the mastoid region in a smooth articulation. The anterior tympanic bulla is fused to part of the anterior process of the tegmen tympani. The medial aspect of the tympanic bulla has no contact with the petrosal, but the posterior aspect of the bulla does have a small contact slightly inset from the ventromedial edge of the promontorium.
The dorsomedial (fig. 86) surface is smooth. The internal acoustic meatus is relatively open, particularly anteriorly, where its margin is poorly defined. The foramen acusticum superius is smaller than the foramen acusticum inferius. The crista transversa is a wide ridge with a dorsal crest. Anterior and superior to the foramen acusticum superius is a rounded prominence that forms part of the prefacial commissure. The prefacial commissure fossa is absent and that area of the petrosal is gently convex. Im- mediately posterior to the foramen acusticum inferius is the vestibular aqueduct. Medial to it is the cochlear aqueduct, a distinct, circular hole, rather than a slit, which occupies a relatively dorsal position. There is no clear basicapsular groove on the anterior or medial surface. More posteriorly still, the petrosal is convex and there is no subarcuate fossa or petromastoid canal. The posterior end drops off abruptly to a smooth mastoid region that is very gently convex on the dorsal side (figs. 79–80).
In dorsolateral view (fig. 79) the tegmen tympani hyperinflation is very clear. The tegmen tympani is smooth with a vascular groove anteriorly and a convex surface overall. As noted above, there is a small, knobshaped ventrolateral tuberosity just anterior to the external acoustic meatus. In ventromedial view (fig. 80) the promontorium is smooth and bulbous and the lack of medial promontory groove is apparent. The cochlear aqueduct is not visible in this view because it is dorsally positioned.
Anteriorly (fig. 81), the hiatus Fallopii is absent as a distinct anterior hole seen in many noncetacean artiodactylans. There is a small hole visible in this area; however, it is not clear that it is the hiatus Fallopii and not a vascular foramen. The shape of the tegmen tympani and the prefacial commissure are also apparent in this view.
ARTIODACTYLA – CETATCEA – ODONTOCETI – DELPHINIDAE
TURSIOPS TRUNCATUS Figures 85 View Fig , 87–91 View Fig View Fig View Fig View Fig View Fig
Oelschläger (1990) and, more recently, Mead and Fordyce (2009) described the anatomy of the petrosal of this species, a cetacean that is widely used for comparative work.
The ventrolateral (fig. 87) surface of the promontorium is fully ovoid and has no significant separate bulge in the region of the fenestra cochleae and no clear transpromontorial sulcus or sulcus for the stapedial artery. The stapes is occasionally fused within the fenestra vestibuli (fig. 87, drawing). The fenestra vestibuli is similar in size to the fenestra cochleae but more oval in outline shape. The fenestra cochleae has an irregular outline, being oval anteriorly and straight posteriorly. There is no round or oval fossa for the tensor tympani, only a small, sharp fold on the ventromedial surface of the anterior process of the tegmen tympani. There is no epitympanic wing or posteromedial flange protruding from the promontorium.
The tegmen tympani is a large structure dorsolateral to the promontorium with a large anterior process that extends distinctly anterior to the promontorium and terminates in a point. The epitympanic recess consists of a large fossa for the head of the malleus and a raised area posterior to it that may represent the fossa incudis (see also Oelschläger, 1990: 149). Medial to the epitympanic recess and tucked deep within the tegmen tympani is the secondary facial foramen. From it emerges the facial sulcus, which is a relatively indistinct channel to the stylomastoid notch; the stapedial muscle fossa is very poorly defined. The stapedial muscle fossa and facial sulcus are partially obscured in ventrolateral view by the relatively large mastoid region and parts of the tympanic bulla that have fused to it. Mead and Fordyce (2009:162) stated that Tursiops truncatus often has a tympanohyal that is very small; however, this structure does not appear to be present in this specimen (I have estimated the position of its attachment). The mastoid region is relatively large with a flat ventral side and a convex dorsal side (figs. 89–90). It comes to a point at the dorsolateral edge where its texture is slightly rough. The caudal tympanic process is a distinct, wide ridge posterior to the fenestra cochleae.
There are several lateral contacts between the ectotympanic and the petrosal (fig. 85). The anterior crus of the tympanic ring contacts the petrosal in a small area just posterior to a large, fused contact between the bulla and the anterior process of the tegmen tympani. There are no contacts between the medial aspect of the bulla and the petrosal. The posterior crus of the tympanic ring is expanded into a large, flat process that has a wide contact with the mastoid region of the petrosal. When not fused, this contact surface on the petrosal is rough.
( Cetacea, Odonotoceti ). Scale 5 1 cm.
The dorsomedial surface (fig. 88) is entirely smooth, including around the internal acoustic meatus. The internal acoustic meatus is a large, reniform opening. The crista transversa is low and poorly defined. Mead and Fordyce (2009:130, using the terminology ‘‘suprameatal fossa of the periotic’’) stated that the prefacial commissure fossa is an indistinct groove lateral to the internal acoustic meatus in Tursiops truncates . I think, however, that the prefacial commissure fossa is absent in this species and that the area dorsolateral to the internal acoustic meatus is flat to convex, not concave. There is a small fold running anteroposteriorly on the pars cochlearis but no clear basicapsular groove. Posterior to the internal acoustic meatus is a circular cochlear aqueduct, which has a raised lip, and a similar-sized vestibular aqueduct with no lip. The cochlear aqueduct is situated on the dorsomedial, not ventromedial, surface of the bone.
Dorsolaterally (fig. 89), the tegmen tympani is gently convex in both anteroposterior and mediolateral directions. Mead and Fordyce (2009) did not observe a vascular groove on the dorsolateral surface of the tegmen tympani; however, the specimen examined here does have a distinct, shallow groove on the dorsolateral surface of the anterior process of the tegmen tympani. This side of the bone also has several very tiny foramina and becomes slightly rough toward the posterior end. The ventrolateral tuberosity is a small bump anterior to the internal acoustic meatus.
In ventromedial view (fig. 90) the hemispherical shape of the promontorium and lack of basicapsular groove are visible. The cochlear aqueduct is partly visible even though it is largely situated toward the dorsomedial surface. The stylomastoid notch is a subtle groove that does not extend along the mastoid region.
Anteriorly (fig. 91), the absence of the prefacial commissure fossa is clear. The tegmen tympani is strongly convex. At its apex, the anterior process of the tegmen tympani assumes a wedge shape. The hiatus Fallopii is absent as a distinct hole seen in many noncetacean artiodactylans. The prefacial commissure fossa is absent.
ARTIODACTYLA – CETACEA – ODONTOCETI – MONODONTIDAE
MONODON MONOCEROS Figures 85 View Fig , 89–93 View Fig View Fig View Fig View Fig View Fig
The ventrolateral (tympanic) surface
(fig. 92) has a smooth hemispherical pro- montorium that lacks an epitympanic wing and posteromedial flange. The promontorium has neither transpromontorial sulci nor a sulcus for the stapedial artery. A broad crista interfenestralis separates the fenestra cochleae and the fenestra vestibuli. The fenestra vestibuli and fenestra cochleae are both relatively round in outline shape. The fenestra cochleae has a slightly irregular outline that is pinched at its medial extreme; the fenestra cochleae is also larger than the fenestra vestibuli. Lateral to the promontorium there is no oval fossa for the tensor tympani muscle; instead, there is simply a shallow channel that does not excavate the adjacent tegmen tympani.
On the pars canalicularis, the anterior process of the tegmen tympani is large, extending well anterior to the promontorium and terminating in a point. The tegmen tympani is hyperinflated, occupying about one-third the total width of the bone. In the epitympanic recess area, there is a large round fossa for the head of the malleus; posterior to this is a much smaller fossa incudis. The external acoustic meatus is a shallow trench lined by ridges of bone with a small, knob-shaped ventrolateral tuberosity at its anterior margin. The crista parotica is distinguished as a clear shelf medial to the fossa incudis and lateral to the secondary facial foramen. No tympanohyal was visible on this specimen, and the approximated place of attachment (if it was present) is indicated. Lateral to the fenestra vestibuli is the secondary facial foramen where it empties into the facial sulcus, which then wraps around the lateral side of the promontorium. The secondary facial foramen is lateral and just slightly anterior to the fenestra vestibuli. The stapedial muscle fossa is not particularly distinct from the narrow channel of the facial sulcus. The facial sulcus ends in a fairly open stylomastoid notch. On the anteromedial side of the pars cochlearis is a wide, knob-shaped caudal tympanic process, which is continuous with the wide crista interfenestralis. The mastoid region is large and the pars cochlearis protrudes medially relative to it (i.e., the pars cochlearis and pars canalicularis meet at an angle medially).
There are several contacts between the bulla and the petrosal (fig. 85). The anterior
( Cetacea, Odonotoceti ). Hatched areas indicated fused parts of tympanic bulla. Scale 5 1 cm.
( Cetacea, Odonotoceti ). Scale 5 1 cm.
crus of the ectotympanic contacts the anterior part of the external acoustic meatus, and the posterior crus of the ectotympanic contacts the mastoid region. The mastoid region is roughened where it articulated with the ectotympanic. The anterior bulla contacts the anterior process of the tegmen tympani where the two bones fused. The contact covers about 50% of the size of the anterior process of the tegmen tympani. Antero- and posteromedially, there is no contact between the bulla and the petrosal.
The dorsomedial (cerebellar) surface (fig. 93) exposes the pars cochlearis anteriorly, which consists of rugose, jagged bone around the internal acoustic meatus. The foramen acusticum superius is circular in outline compared to the foramen acusticum inferius, which is oval in outline, and is the larger of the two foramina. The foramen singulare can be clearly identified in this specimen just posterior to the crista transversa. The pars cochlearis is spherical medially and distinguished by the absence of a basicapsular groove. The prefacial commissure fossa is absent; instead, that area of the petrosal is slightly convex. The hyperinflated tegmen tympani is also convex both mediolaterally and anteroposteriorly. As noted above, but also visible in this view, is the anterior process of the tegmen tympani as it extends anterior to the pars cochlearis. The mastoid region has a convex dorsal surface and a flat ventral surface (see also figs. 89– 90). Two other openings visible on the endocranial surface are the cochlear aqueduct and the vestibular aqueduct, both of which are relatively dorsal in position. There is no subarcuate fossa or petromastoid canal.
The dorsolateral side (fig. 89) of the petrosal is dominated by the hyperinflated tegmen tympani, which has a curved, smooth surface. The overall outline of the bone in this view is rounded. Posteriorly, the surface texture of the mastoid region is pocked. There are no vascular grooves or spikes on the lateral surface. The ventrolateral tuberosity is a small, knob-shaped structure.
Ventromedially (fig. 90), the transition on the pars cochlearis from the roughened area adjacent to the internal acoustic meatus to the smooth promontorium ventrally is clearly delineated. The absence of the basicapsular groove is again visible here. The cochlear aqueduct is positioned within the jagged area of bone on the endocranial surface. The mastoid plate is absent.
Anteriorly (fig. 91), the hemispherical shape of the promontorium is apparent, as is the absence of a hiatus Fallopii that is a distinct anterior hole. Despite the hyperinflated and convex tegmen tympani, as noted above, the bone does not have a prefacial commissure fossa.
ARTIODACTYLA – CETACEA – ODONTOCETI – ZIPHIIDAE
ZIPHIUS CAVIROSTRIS Figures 85 View Fig , 89–91 View Fig View Fig View Fig , 94–95 View Fig View Fig
The petrosal of Ziphius was figured by Kasuya (1973).
The ventrolateral (tympanic) surface of the petrosal (fig. 94) has a smooth promontorium that is hemispherical and that lacks both an epitympanic wing and a posteromedial flange extending either anteriorly or medially from it. The fenestra vestibuli is positioned anterior to the fenestra cochleae and has a circular outline shape. The fenestra cochleae is much larger than the fenestra vestibuli and has a generally oval, but slightly irregular, outline shape. A broad crista interfenestralis separates the two fenestrae. A probe inserted in the fenestra cochleae can pass directly to the large cochlear aqueduct. There is a ridge running on the promontorium just lateral to the fenestra cochleae. There are no transpromontorial or stapedial artery sulci on the promontorium. A fully defined oval fossa for the tensor tympani is absent.
On the pars cochlearis, the anterior process of the tegmen tympani is large and ends anteriorly in a point, from which extends a ridge posterolaterally. The tegmen tympani itself is hyperinflated, accounting for almost half the size of the bone, with a convex lateral margin. Directly anterior to the secondary facial foramen is an oval depression that is the fossa for the head of the malleus. Within the epitympanic recess is also a small fossa incudis inset on a triangular wedge of bone that abuts the anterior extreme of the crista
( Cetacea, Odonotoceti ). Arrow indicates connection between two foramina. Scale 5 1 cm.
parotica. The crista parotica wraps as a distinct crest lateral to the facial sulcus; a possible, but unconfirmed, attachment site for the tympanohyal is indicated. Just poste- rior to a knob-shaped ventrolateral tuberosity is the external acoustic meatus, a shallow trough lateral to the opening of the secondary facial foramen, which emerges from deep within the roof of the fossa for the head of the malleus and feeds onto the facial sulcus. The facial sulcus is a relatively open and broad trough that winds lateral to the stapedial muscle fossa, with the latter being a shallow, oval area near the stylomastoid notch. A caudal tympanic process arises posterior to the fenestra cochleae and stapedial muscle fossa and is a low, wide bump that exhibits continuity with the crista interfenestralis. The large mastoid region has a smooth, featureless flat surface ventrally, as observed by Kasuya (1973: 18). The pars cochlearis protrudes medially relative to the mastoid region, such that there is an angle formed approximately at the anteromedial aspect of the mastoid region.
There are several contacts (fig. 85) between the petrosal and the tympanic. Two of these are between the ectotympanic and the lateral petrosal. The anterior crus of the tympanic ring contacts the ventrolateral tuberosity. The posterior crus of the tympanic ring contacts a large, smooth area on the mastoid region. The anterior tympanic bulla contacts and fuses to the anterior process of the tegmen tympani. This contact occurs over approximately one-third the total size of the anterior process of the tegmen tympani. The medial tympanic bulla contacts the promontorium, not at its ventromedial edge as in many other taxa but at a small area anterior to the fenestra cochleae. The posterior bulla does not contact the petrosal.
The dorsomedial (cerebellar) surface (fig. 95) is completely smooth. On the pars cochlearis, the internal acoustic meatus lacks a well-defined edge surrounding both the foramen acusticum superius and the foramen acusticum inferius. The foramen acusticum superius is smaller than the foramen acusticum inferius. A wide and relatively tall crista transversa separates the two foramina. Medially, the pars cochlearis is ovoid and appears to have a very subtle indication of a basicapsular groove. In this view it is apparent that on the pars canalicularis, the anterior process of the tegmen tympani is extremely large, that it terminates anteriorly as a point, and that the tegmen tympani is convex and hyperinflated. Dorsolateral to the internal acoustic meatus, the pars cochlearis is convex and lacks a prefacial commissure fossa. Posterior to the internal acoustic meatus are two foramina, the vestibular and cochlear aqueducts. These are relatively large, are positioned close to each other, and are on the dorsomedial surface. The cochlear aqueduct is so large that it wraps partially around to the ventromedial surface. There is no subarcuate fossa or petromastoid canal. There is a large, irregular depression on the dorsal surface of the mastoid region. The mastoid region is convex on the dorsal surface.
The dorsolateral surface (fig. 89) is convex in two directions, mediolaterally and anteroposteriorly. The tegmen tympani is hyperinflated. It has a small curved groove at the posterior part of its anterior process, a feature that was also observed in specimens examined by Kasuya (1973). It also has a small mediolaterally oriented groove just anterior to that, both of which may have been vascular grooves. The anterior process has a smooth square facet and the mastoid region has three small (vascular?) foramina. The ventrolateral tuberosity is small and knob-shaped.
Ventromedially (fig. 90), a faint ridge is visible on the pars cochlearis, roughly demarcating the dorsomedial from the tympanic side. The faint basicapsular groove noted above is situated toward the dorsomedial side. Most directly visible in medial view is the large cochlear aqueduct, as well as the relatively large size of the anterior process of the tegmen tympani and the mastoid region. The full continuity between the cochlear aqueduct and the fenestra cochleae is also visible from this side of the bone (fig. 90, arrow). The mastoid region has a flat ventral surface and a convex dorsal surface; the mastoid plate is absent. The dorsolateral view (fig. 89) demonstrates the knob-shaped ventrolateral tuberosity, the shape of the mastoid region, and the small vascular groove situated at the anterior aspect of the tegmen tympani.
Anteriorly (fig. 91), it is apparent that there is no distinct anterior hole demarcating the hiatus Fallopii as seen in many noncetacean artiodactylans. The convexity of the dorsal surface of the tegmen tympani is also apparent, as is its large size and the absence of the prefacial commissure fossa.
ARTIODACTYLA – CETACEAMORPHA – † PAKICETIDAE
† PAKICETUS INACHUS Figure 96 View Fig
This specimen has been described and figured elsewhere ( Gingerich and Russell, 1981; Oelschläger, 1987; Thewissen and Hussain, 1998; Luo, 1998; Luo and Gingerich, 1999); however, because of its relevance to the phylogeny, its transitional morphology, and its relative completeness, it is reviewed again here to facilitate direct comparison with many artiodactylans to which it has not previously been compared. Other † pakicetid basicranial material has been noted for this taxon ( Nummela et al., 2004), but no other petrosal has actually been described or figured for this taxon. This description is based on a cast.
The petrosal is in situ, and thus only the ventrolateral surface is visible for description (fig. 96). The promontorium is smooth and oval in outline (hemi-ellipsoid) and does bulge ventrally except just over the fenestra vestibuli. The fenestra cochleae and fenestra vestibuli are both oval in outline, are roughly the same size, and are separated by a relatively narrow crista interfenestralis. As noted by Luo and Gingerich (1999), there are no transpromontorial sulci or sulci for the stapedial artery crossing the promontorium. I agree with Luo and Gingerich (1999) that one cannot tell from this specimen if there was a basicapsular groove. Extending from the promontorium anteriorly and medially are an epitympanic wing and a posteromedial flange.
The pars canalicularis is not fully visible given that the specimen is in situ and the adjacent squamosal bone covers part of the tegmen tympani. There is a deep oval pit between the promontorium and what is visible of the tegmen tympani, which is most likely the fossa for the tensor tympani muscle. This is not well preserved on the specimen here but can be corroborated by in HGSP-96386, where it is also clear that this fossa does not excavate the surrounding tegmen tympani. Posterior to the fossa for the tensor tympani muscle is a second elongate pit that may be slightly broken on the edges. This pit is probably the secondary facial foramen feeding onto the facial sulcus. The size and shape of the tegmen tympani are difficult to assess because the petrosal is tightly articulated with the adjacent squamosal. It is not clear whether the ventrolateral tuberosity was present. The epitympanic recess is nondescript and has no large fossa for the head of the malleus as in extant cetaceans. There is a wide caudal tympanic process posterior to the fenestra cochleae that separates it distinctly from the stapedial muscle fossa. Luo and Gingerich (1999) interpreted a small bump in the region of the stapedial muscle fossa to be the attachment site for the tympanohyal. The mastoid region is rough and elongate, exceeding 100% of the length of the promontorium. The pars cochlearis meets the mastoid region at an angle of approximately 150 °, and thus it protrudes medially relative to the mastoid region. The mastoid region is tightly articulated with the adjacent occipital and squamosal bones. The mastoid region of Pakicetus forms the posterior wall of the dorsal portion of the external acoustic meatus. I point this out because in many other taxa this wall is formed by the squamosal.
On the right side of the skull the tympanic bulla is in situ; however, it is difficult to tell exactly how this bone contacted the petrosal. As noted by Luo and Gingerich (1999), there does appear to have been a clear articulation between the posterior crus of the ectotympanic (which they noted is quite expanded in this species) and the large mastoid region superior to it. Anteriorly, there may have been contact between the anterior crus of the ectotympanic and the petrosal but it is difficult to be certain of this. There was a clear articulation between the anterior tympanic bulla and the squamosal; however, it is not clear whether this contact also extended to the petrosal. The medial bulla certainly contacted the basioccipital, but it is not clear whether there was a separate contact between the bulla and the medial promontorium.
ARTIODACTYLA – CETACEAMORPHA – † BASILOSAURIDAE
† BASILOSAURUS ISIS Figures 97 View Fig , 99–102 View Fig View Fig View Fig View Fig
The specimen is an isolated petrosal that has a broken mastoid region, and it was also figured by Luo and Gingerich (1999: 19, 20B, D). I have no new information on contacts of this bone with the auditory bulla.
The ventrolateral pars cochlearis (fig. 97) of the petrosal consists of a smooth promontorium with a pronounced convexity anteromedial to the fenestra cochleae. The fenestra cochleae and fenestra vestibuli are roughly equal in size, but the fenestra cochleae is slightly more oval than the fenestra vestibuli. Extending from the fenestra cochleae is a short but distinct groove that runs to the ventromedial edge of the tympanic surface. The crista interfenestralis is a wide ridge such that the fenestrae are separated in a pronounced fashion. The promontorium is ovoid and drops off steeply at its anterior and medial edges, indicating an absence of the epitympanic wing and posteromedial flange. The texture of the medial surface of the promontorium is characterized by thin vertical grooves. The promontorium has a clear, broad groove running posteromedial to anterolateral; however, in previous descriptions ( Luo and Gingerich, 1999) this has not been considered a transpromontorial sulcus. There is no clear fossa for the tensor tympani muscle. Luo and Gingerich (1999: fig. 20B, D) suggested that this muscle occupied a shallow groove between the promontorium and the tegmen tympani.
On the pars canalicularis, the tegmen tympani is hyperinflated and extends well anterior to the promontorium. The entire anterior process of the tegmen tympani ends at an apex that tapers to a point. There is a large fossa for the head of the malleus that is round, shallow, and marked by a knobshaped ventrolateral tuberosity anteriorly. Posterior to this is a small fossa incudis ( Luo and Gingerich, 1999). The secondary facial foramen emerges just posterior to the fossa for the head of the malleus and feeds onto a facial sulcus directed toward the medial edge of the promontorium. The stapedial muscle fossa may have been present but the area is broken posteriorly and it is difficult to determine the extent of this structure. The crista parotica is partly preserved where it runs lateral to the secondary facial foramen. The caudal tympanic process is wide mediolaterally and situated posterior to the fenestra cochleae. Although the mastoid region is not preserved in this specimen, the promontorium protruded medially relative to it based on comparisons with complete specimens ( Kellogg, 1936; Luo and Gingerich [1999] did not present new information on the mastoid region).
Luo and Gingerich (1999), in a general description of the tympanic-petrosal complex in Basilosauridae , described contacts between the petrosal and the modified anterior and posterior crura of the tympanic (fig. 98). The articulation of the posterior crus of the tympanic ring is particularly large (fig. 98). These authors did not indicate contacts between the medial petrosal and the medial tympanic bulla.
The dorsomedial surface of the petrosal (fig. 99) is extremely rough, including the area surrounding the internal acoustic meatus (this is the original, undamaged surface; P.D. Gingerich, personal commun.). The specimen has a large, oval, internal acoustic meatus with a tall lip at the anterior and posterior ends. Posterior to this are the cochlear and vestibular aqueducts (see also fig. 101), which are relatively small, closely appressed, and situated on the internal surface. Dorsolaterally, there is an extremely large prefacial commissure fossa that occupies a significant portion of the petrosal. This convex area extends both anterior and posterior to the internal acoustic meatus and excavates the dorsal surface of the hyperinflated tegmen tympani. There is a small, deep hole at the junction of the anterior process and the pars cochlearis ( Luo and Gingerich [1999] also did not label this). Both the subarcuate fossa and the petromastoid canal are absent.
Dorsolaterally (fig. 100), the hyperinflated tegmen tympani is visible with a vascular groove at the base of the anterior process. This groove runs the full length of the lateral surface dorsally to medially. The lateral surface is generally smooth but is marked by a few knobs and pits, and it is convex in overall shape. The ventrolateral tuberosity can also be seen in this view as a small, knobshaped structure.
Ventromedially (fig. 101), as noted above, the promontorium forms a distinct hemisphere with a sharp edge where it meets the endocranial surface of the petrosal. There is no basicapsular groove. Luo and Gingerich (1999: fig. 20B, D) suggested that the hiatus Fallopii lies deep within the anterior process on the medial surface, which means it should therefore be visible in anterior view (fig. 102). I think, however, it is not clear exactly where this foramen was positioned, but it is evident that a distinct anterior hole as is seen in many noncetacean cetartioadactylans was absent. The large prefacial commissure fossa is clear in this view as a large excavation in the hyperinflated tegmen tympani.
(Cetaceamorpha, Basilosauridae ). Scale 5 1 cm.
ARTIODACTYLA – CETACEAMORPHA - † BASILOSAURIDAE
† DORUDON ATROX Figures 100–104 View Fig View Fig View Fig View Fig View Fig
The specimen examined here has also been described and figured by Luo and Gingerich (1999: figs. 1E, 2C, 20A, C) and noted by Uhen (2004). It is refigured here because of its comparative relevance to the homologies in many artioadactylans. This petrosal is incompletely preserved. The posterior end, which consisted of an elongate mastoid region ( Kellogg, 1936; see also Luo and Gingerich, 1999), is not present. Part of the anterior process of the tegmen tympani is also broken. Figure 103 View Fig does not show restored anterior and posterior processes as in Luo and Gingerich (1999: fig. 2C), but the matrix reflects the information in Luo and Gingerich (1999: fig. 20C) on more complete specimens. I present no new information on the bulla in this taxon.
On the ventrolateral pars cochlearis (fig. 103), the promontorium is smooth and very bulbous, particularly anterior to the fenestra cochleae, where there is a pronounced, elongate bulge. The fenestra vestibuli and fenestra cochleae are close in size. The fenestra cochleae is oval and irregular in outline, and the fenestra vestibuli is round in outline shape. Luo and Gingerich (1999: 45) made the interesting observation that there is a clear groove extending from the fenestra cochleae to the medial edge of the promontorium in this species. This is indeed the case. The crista interfenestralis is a wide, bulging process of bone intervening between the two fenestrae. There are no sulci crossing the promontorium. The promontorium is more hemispherical than hemi-ellipsoid in shape (see also fig. 102), and the pars cochlearis lacks both an epitympanic wing and a posteromedial flange, as there are no distinct, flattened processes extending anterior or medial from the promontorium. There is no distinct excavation indicating a fossa for the tensor tympani as seen in noncetacean artiodactylans. Luo and Gingerich (1999) hypothesized that the muscle would have instead occupied an open groove anterior to the fenestra vestibuli, and they suggested that the groove between the promontorium and the tegmen tympani was the most likely candidate for the homolog of the fossa for the tensor tympani.
The lateral third of the ventrolateral surface is occupied by a hyperinflated tegmen tympani, which has a large anterior process that extends anterior to the promontorium. Luo and Gingerich (1999) stated that the anterior process of the tegmen tympani was pointed, but this cannot be verified in this specimen and may have been based on other data. A large fossa for the head of the malleus that is shallow and round sits between the inflated tegmen tympani and the promontorium and occupies a relatively anterior position. Posterior to this is a small fossa incudis ( Luo and Gingerich, 1999). Luo and Gingerich (1999: 45; see also Geisler and Luo, 1996) stated that the relative sizes and depths of these structures resembled those of odontocetes but not mysticetes. Both of these fossae are ventrally offset relative to the adjacent secondary facial foramen, just posterior to the fossa for the head of the malleus and medial to the fossa incudis. A distinct facial sulcus extends from the secondary facial foramen toward the medial edge of the promontorium. The area where the stapedial muscle fossa may have been is damaged and the nature of this structure is uncertain ( Luo and Gingerich [1999] indicated that it is present). The original specimen does not show clear evidence of a tympanohyal as indicated by Luo and Gingerich (1999); however, this area is poorly preserved and the presence or absence of this structure cannot be decided with confidence. The crista parotica is only partially intact. The ventrolateral tuberosity anterior to the external acoustic meatus is small and knob-
(Cetaceamorpha, Basilosauridae ). Scale 5 1 cm. Hatching indicates damaged areas.
(Cetaceamorpha, Basilosauridae ). Hatched areas are broken. Scale 5 1 cm.
shaped. The external acoustic meatus has to be approximated due to poor preservation.
Luo and Gingerich (1999: 46), in a general description of the tympanic-petrosal complex in Basilosauridae , described contacts between the lateral side of the petrosal and the anterior and poster crura of the ectotympanic (which are highly transformed in cetaceans). The articulation of the posterior crus is particularly large. They did not describe contacts between the medial petrosal and the medial bulla. As noted above, I have no new information to report here on bullar contacts.
The dorsomedial surface of the petrosal (fig. 104) is roughly square given that the mastoid region is not preserved. The dorsomedial surface texture is rough, including the area surrounding the internal acoustic meatus. This species has a large, oval internal acoustic meatus divided by a thin, sharp crista transversa. At the posteroventral margin of the internal acoustic meatus is a small bump. Dorsolateral to the internal acoustic meatus is a large prefacial commissure fossa, an extensive convexity on the endocranial surface of the tegmen tympani. The extent of the hyperinflated tegmen tympani, which is convex (‘‘barrel-shaped,’’ Geisler and Luo [1998]), can be seen in this view as well as in anterior (fig. 102) and dorsolateral (fig. 100) views. Posterior to the internal acoustic meatus are two smaller foramina, the endolymphatic and perilymphatic foramina (as indicated by Luo and Gingerich, 1999). The subarcuate fossa and petromastoid canal are both absent.
In dorsolateral view (fig. 100), crossing the lateral side of the anterior process of the tegmen tympani is a large dorsoventrally oriented vascular groove. Luo and Gingerich (1999: 45) stated that this groove is also present in † pakicetids and † protocetids. Ventromedially (fig. 101), the promontorium transitions from smooth on the tympanic side to very rough on the dorsomedial side. There is no basicapsular groove and the cochlear aqueduct is not visible because it is on the endocranial surface.
An anterior view (fig. 102) shows the excavation associated with the prefacial commissure fossa, as well as the sharp edge demarcating the division between the dorsal and ventral surfaces of the bone. It also indicates that there is no distinct hiatus Fallopii. Luo and Gingerich (1999) hypothesized that the greater petrosal nerve branches within the tympanic cavity in this taxon.
† MESONYCHIA – † MESONYCHIDAE
† DISSACUS PRAENUNTIUS Figures 105–109 View Fig View Fig View Fig View Fig View Fig
The petrosal specimen discussed here was also described and figured by Luo and Gingerich (1999: figs. 3B, 6–7). The specimen is broken at its anterolateral margin and posteriorly across the pars canalicularis (with the latter piece being separately preserved). I had access to the anterior part of this specimen only, and thus information in the matrix regarding the mastoid region comes from Luo and Gingerich (1999).
The promontorium (fig. 105) is hemiellipsoid and bulbous over the fenestra cochleae, tapering into a flat shelf, the epitympanic wing, toward the anteromedial border. More posteriorly off the pars cochlearis is a posteromedial flange that was probably continuous with the epitympanic wing; however, the specimen is not fully intact in this area. There are no distinct transpromontorial grooves; however, note that Luo and Gingerich (1999: fig. 7) did reconstruct a transpromontorial course for the internal carotid artery in this taxon. These authors also suggested that there may be a sulcus for the stapedial artery, but I find this difficult to see. The tegmen tympani is moderately inflated (see also Luo and Gingerich, 1999: 23), measuring approximately one-third the total width of the petrosal. The anterior process of the tegmen tympani is broken, so its full size and shape are unknown, as is its relationship to the promontorium. Luo and Gingerich (1999) stated, however, that there was an anterior process of the tegmen tympani that bore a ‘‘striking resemblance to the anterior process of the cetacean petrosals’’; however, based on what is preserved of this specimen, I think this process is no more similar to the anterior process of cetaceans than it is to the anterior process of most noncetacean artiodactylans.
( † Mesonychia , † Mesonychidae ). Hatched areas are broken. Scale 5 1 cm.
( † Mesonychia , † Mesonychidae ). Hatched areas are broken. Scale 5 1 cm.
Luo and Gingerich (1999: 23) described the presence of a ventrolateral tuberosity; this is very small in this taxon and is shaped as a small bump.
The fossa for the tensor tympani is slightly broken and was likely relatively deep and oval-shaped. It appears not to have excavated the adjacent tegmen tympani. The fenestrae cochleae is round (with part of its margin broken); the fenestra vestibuli is not fully preserved, and thus it is hard to characterize other than identification of its general position. Luo and Gingerich (1999) stated that the specimen had a small hiatus Fallopii at the anterior aspect of the tegmen tympani; however, the preservation in this area makes this hard to confirm. The secondary facial foramen is not well preserved ( Luo and Gingerich [1999: 26] also suggested that it is present) but it appears to have occupied a relatively anterior position. The facial sulcus is distinct as it wraps lateral to the stapedial muscle fossa. The epitympanic recess is small and there is no large fossa for the head of the malleus as in cetaceans. The external acoustic meatus is only partially preserved but did receive a petrosal contribution. The crista parotica denotes a clear rim adjacent to the facial sulcus. The tympanohyal is not preserved, but Luo and Gingerich (1999) suggested that it was present and its position is inferred here. The mastoid region (not figured here) is elongate. Luo and Gingerich (1999: 23) hypothesized that it did not articulate with the tympanic bulla (fig. 98); however, the bulla is not preserved and no information about bullar contacts is provided here. Posterior to the fenestra cochleae on the pars canalicularis is broad bump that is the caudal tympanic process.
The dorsomedial (fig. 106) surface is smooth. The internal acoustic meatus is a long, oval structure with a well-defined posterior border and a poorly defined anterior border. The crista transversa is a narrow ridge.
Superior to it is a distinct prefacial commissure fossa bordered distolaterally by a sharp ridge that runs anteroposteriorly. The tegmen tympani is inflated and convex. Luo and Gingerich (1999: 23) stated that the tegmen tympani inflation in † Dissacus is roughly proportional to that seen in cetaceamorphans, however, I observe that in cetaceamorphans the inflation is much more pronounced.
Extending from the anterior margin of the petrosal on the dorsal side, along its medial edge, is a distinct basicapsular groove. Posterior to the groove is a small indentation, the cochlear aqueduct. Adjacent to the cochlear aqueduct is a small triangular tuberosity. Posterior and slightly dorsal to the internal acoustic meatus is a shallow subarcuate fossa with a petromastoid canal. The mastoid region is broken and was not available for study.
† Mesonychidae ) and † Mesonyx obtusidens (AMNH-VP 12643) ( † Mesonychia , † Mesonychidae ).
In dorsolateral view (fig. 107) the convex tegmen tympani is conspicuous, as is a clear groove on it that Luo and Gingerich (1999: 23) suggested was for the ‘‘ramus superior of the stapedial artery.’’ There is a small, knobshaped ventrolateral tuberosity.
In ventromedial view (fig. 108) the basicapsular groove is clear. There is also a protuberance at the posterior aspect of the promontorium. The endolymphatic and perilymphatic foramina cannot be seen very clearly but can be approximated.
In anterior view (fig. 109) the prefacial commissure fossa can be seen as a clear concavity dorsal to the internal acoustic meatus. The promontorium has a convex bulge limited to its central region, in contrast to the fully hemispherical shape seen in cetaceans. The hiatus Fallopii is not well preserved.
† MESONYCHIA – † MESONYCHIDAE
† MESONYX OBTUSIDENS Figures 98 View Fig , 107–111 View Fig View Fig View Fig View Fig View Fig
The bone is generally well preserved and only slightly broken at the anterodorsal margin; it is smooth. This specimen was also
( † Mesonychia , † Mesonychidae ). Scale 5 1 cm. Hatched areas is site of tympanohyal attachment.
( † Mesonychia , † Mesonychidae ). Scale 5 1 cm.
figured and described by Geisler and Luo (1998: fig. 5).
The promontorium (fig. 110) is hemiellipsoid with a gentle bulge for the underlying cochlea. The fenestra cochleae and fenestra vestibuli are similar in size, relatively circular in outline shape, and are separated by a distinct, ridgelike crista interfenestralis. There is a very subtle transpromontorial sulcus and a more pronounced sulcus for the stapedial artery. The extent of the fossa for the tensor tympani is not clear because this part of the bone is broken. The pars cochlearis lacks significant flanges extending from the promontorium; both the epitympanic wing and the posteromedial flange are absent.
The pars cochlearis constitutes a significant part of the bone’s size. The tegmen tympani is inflated, accounting for approximately 30% of the width of the petrosal in ventral view. The secondary facial foramen is not preserved but there is a facial sulcus, which contains a deep depression, and a clear stylomastoid notch. The fossa for the stapedial muscle sits immediately posterior to the fenestra cochleae. The epitympanic recess is an unremarkable shallow pit that is positioned more ventrally than the facial sulcus; there is no distinct fossa for the head of the malleus as seen in cetaceans. The crista parotica is preserved, except at its most anterior point, and is a clear crest running to the base of the tympanohyal attachment site. The tympanohyal is not preserved but its attachment site may be inferred. There is a broad caudal tympanic process that flares posteromedial to the fenestra cochleae. The caudal tympanic process is continuous with a sinuous crest that extends from the crista interfenestralis. Regarding the medial side of the specimen in ventral view, there is some protrusion of the pars cochlearis relative to the mastoid region.
Although part of the tympanic is preserved, the contacts between the tympanic and the petrosal are not clearly preserved. Two potential sites of contact are indicated, one for the anterior crus of the ectotympanic and a second for the medial edge of the bulla.
In dorsomedial view (fig. 111), the internal acoustic meatus is relatively ventromedial in position and has a nondescript margin. The crista transversa is a small ridge. Superior to the internal acoustic meatus is a prefacial commissure fossa that is broken at its anteromedial edge. The tegmen tympani is convex and moderately inflated. A basicapsular groove extends from the anterior and medial edge to just medial to the internal acoustic meatus. The entire dorsomedial surface is smooth. The subarcuate fossa is defined endocranially by a low, convex depression; there is no petromastoid canal. A ridge that merges with the relatively large mastoid region defines the fossa laterally. The mastoid region is generally wedgeshaped and relatively large, with a blunt posterior end. The cochlear aqueduct and the vestibular aqueduct are not clearly preserved; however, it is apparent that this taxon does not have a very dorsally positioned enlarged cochlear aqueduct as seen in some cetaceans.
The dorsolateral (fig. 107) surface of the bone is convex both anteroposteriorly and mediolaterally. It has a distinct vascular groove. The ventrolateral tuberosity is absent and the large wedge shape of the mastoid region is visible in this view (as in fig. 108), as is the small, blunt anterior process. Ventromedially (fig. 108), the pars cochlearis is distinguished medially as having a flat edge, and the basicapsular groove is positioned closer to the dorsomedial surface. There is no mastoid plate.
Anteriorly (fig. 109), the prefacial commissure fossa is a clear excavation dorsal to the internal acoustic meatus. The tegmen tympani is convex and moderately inflated. The hiatus Fallopii is not clearly preserved but it may have been a small foramen, because no such hole is preserved and the bone is not enormously damaged in that area.
PERRISODACTYLA – HIPPOMORPHA – EQUIDAE
EQUUS CABALLUS Figures 112–117 View Fig View Fig View Fig View Fig View Fig View Fig
On the pars cochlearis, the ventrolateral or tympanic (fig. 112) surface has a smooth promontorium with no sulcus for the stapedial artery or transpromontorial grooves. The promontorium is hemi-ellipsoid in shape and the underlying cochlea causes the surface to bulge ventrally. The fenestra cochleae is oval and smaller than the fenestra vestibuli, and the fenestra vestibuli is slightly reniform in outline. Between the fenestra cochleae and the fenestra vestibuli is a fairly pronounced crista interfenestralis that is sharp. The fossa for the tensor tympani is a large, shallow, oval fossa that lies anterior to the fenestra vestibuli. In life it was apparently occupied by a muscle that Wilkie (1931: 108) described as being of ‘‘considerable size.’’ The thin roof of this fossa has broken in this specimen. The fossa does not significantly excavate the surrounding tegmen tympani. Anteriorly, the promontory gives rise to a large epitympanic wing. This wing comes to a point anteriorly and extends to the tegmen tympani. The relatively small opening for the hiatus Fallopii is situated at the junction of the epitympanic wing and the tegmen tym- pani (figs. 112, 117). Getty (1975: 722) remarked that a groove for the superficial petrosal nerve is present on the petrosal, and I can confirm that a groove at the hiatus is visible just anterior to the fossa for the tensor tympanic muscle (fig. 112). There also extends from the promontorium a posteromedial flange that joins the epitympanic wing such that the promontorium is surrounded by a complete, flat flange of bone.
The pars canalicularis is a large, rough section of the petrosal that encircles the promontorium on the lateral and posterior sides. The tegmen tympani is moderately inflated and contributes to about one-fifth the total width of the ventrolateral view of this bone. The anterior process of the tegmen tympani is large, extends anterior to the promontorium, and terminates in a point. Posterior and lateral to the fossa for the tensor tympani noted above is a large, semicircular excavation, which is the petrosal contribution to the external acoustic meatus. Both the anterior and posterior extremes of this trough are ventrally extensive (into the plane of fig. 112). The ventrolateral tuberosity is present at the anterior aspect of the external acoustic meatus and is knob-shaped. Medial to the external acoustic meatus is a nondescript epitympanic recess. The epitympanic recess is lateral to the secondary facial foramen and offset ventrally from it. The secondary facial foramen feeds onto the facial sulcus immediately lateral to the fenestra vestibuli, and thus occupies a relatively anterior position. The crista parotica is a sharp ridge separating the more deeply set facial sulcus and extends medial to the epitympanic recess to the base of the tympanohyal attachment site. The facial sulcus passes lateral to a fossa for the stapedial muscle that is particularly round and deep as noted by Wilkie (1931). The stylomastoid notch is a distinct, elongate trough medial to the tympanohyal attachment site. The caudal tympanic process is a wide and relatively smooth ledge of bone that is both posterior to the fenestra cochleae and also extends medially. The mastoid region is large, occupying approximately half the size of the bone. It is wedge-shaped, irregular, and knobby. It meets and joins the caudal tympanic process. The medial aspect of the pars cochlearis protrudes relative to the pars canalicularis such that there is an angle between these two parts of the petrosal in ventrolateral view.
The tympanic bulla contacted the petrosal in several areas (fig. 113). These include contact between the anterior crus of the ectotympanic and the petrosal, and the posterior crus of the ectotympanic and the mastoid region; the latter is a site of partial fusion. The anterior aspect of the auditory bulla contacted and fused with the anterior process of the tegmen tympani. The medial and posterior aspects of the bulla contacted an elongate, continuous area on the medial and posterior promontorium extending along the posteromedial flange and the caudal tympanic process. These parts of the bulla do not abut the medial extreme of the petrosal but are inset laterally.
The dorsomedial (fig. 114) surface has a smooth texture with one large, irregularshaped opening, the internal acoustic meatus, which is offset toward the dorsal surface of the bone. The crista transversa is a sinuous ridge of bone between the foramina acusticum superius and inferius. The prefacial commissure is a nondescript ridge, and the prefacial commissure fossa is absent. Superi- or to the internal acoustic meatus is the large tegmen tympani, the superior and medial edges of the bone meet at right angles and distinctly demarcate the superior border of the posterior cranial fossa. The ventromedial edge of the bone has two elongate ridges that demarcate the basicapsular groove. At the posterior edge of these is the pin-sized cochlear aqueduct. Posterior to the internal acoustic meatus is a convexity that is followed posteriorly by a shallow, nondescript pit, the subarcuate fossa. There is no petromastoid canal. At the ventral edge of the convex ridge is the vestibular aqueduct, which opens as a slit. The surface tapers posteriorly into a highly cancellous mastoid region (partially cut in this specimen) that terminates in a point.
In dorsolateral view (fig. 115) the extensive irregular shape of the tegmen tympani is clear. As noted above, this surface is smooth and is set at a right angle to the endocranial (dorsomedial) surface. The anterior process of the tegmen tympani ends in a blunt tip and the hiatus Fallopii is tucked deep within it. Toward the mastoid region, the lateral surface has several raised bumps. There are no vascular grooves. The knob-shaped ventrolateral process marks the anterior border of the external acoustic meatus.
The ventromedial surface (fig. 116) of the petrosal is narrow anteriorly and expands into an extensive fan-shaped mastoid region posteriorly. The medial surface is somewhat flat, and the basicapsular groove can be seen to occupy a relatively dorsal position. The cochlear aqueduct is a very small hole at the ventromedial margin. In this view also visible just medial to the caudal tympanic process is a groove (possibly vascular). The posterior aspect of the mastoid region is a large triangular area that forms a knob ventrally and a point dorsally. Posteriorly, the petrosal mastoid region is exposed on the external surface of the braincase over an elongate area and the mastoid plate is absent.
The anterior view (fig. 117) not only shows the relatively small hiatus Fallopii mentioned above, but also shows that the superiormost aspect of the petrosal projects endocranially (medially).
PERRISODACTYLA – HIPPOMORPHA – EQUIDAE
TAPIRUS TERRESTRIS Figures 113 View Fig , 115–119 View Fig View Fig View Fig View Fig View Fig
In ventrolateral (tympanic; fig. 118) view, the pars cochlearis is hemi-ellipsoid and the promontorium has a subtle bulge anterior to the fenestra cochleae for the underlying cochlea. The promontorium has no sulcus for the stapedial artery or other conspicuous transpromontorial grooves. The fenestra cochleae is larger than the fenestra vestibuli, and both are relatively oval in outline shape. The crista interfenestralis is a thick bar of bone that terminates at the anterior margin of the facial sulcus. Extending from the promontorium anteriorly is a flattened flange, the epitympanic wing. The epitympanic wing comes to a point medially and also extends laterally such that it is fully continuous with the tegmen tympani. The epitympanic wing is also continuous with a posteromedial flange, a flattened shelf that extends medially from the promontorium. Together the epitympanic wing and posteromedial flange comprise a full lip around the anterior and medial aspect of the promontorium. A large, very shallow fossa for the tensor tympani muscle lies anterior to the fenestra vestibuli and medial to the tegmen tympani. The tegmen tympani is partially excavated by the fossa for the tensor tympani.
The pars canalicularis is extensive, forming a broad, thick shelf that wraps lateral and posterior to the pars cochlearis. The tegmen tympani is moderately inflated, occupying approximately half of the ventral width of the petrosal. It terminates anteriorly in a blunt anterior process that does not extend far anterior to the promontorium. Dorsal to it, floored by the thin sheet of bone that forms the roof of the fossa for the tensor tympani, is the anteriorly positioned secondary facial foramen emerging onto the facial sulcus. The facial sulcus follows a long channel toward the ventromedial edge of the bone. Lateral to the facial sulcus is the epitympanic recess. This area is ventrally displaced relative to the facial sulcus and lacks a distinct, prominent fossa for the head of the malleus. The external acoustic meatus contribution from the petrosal is an indistinct trough at the anterior base of the tympanohyal. Anterior to the external acoustic meatus there is no ventrolateral process. The tympanohyal is still attached in this specimen and is a prominent, ventrally extended structure that is several centimeters long and terminates in a wide, blunt tip. The crista parotica extends as a clear ridge of bone from the base of the tympanohyal, medial to the epitympanic recess, along the lateral border of the facial sulcus. The fossa for the stapedial muscle is a shallow, oval trench. The caudal tympanic process is a broad shelf of bone posterior to the fenestra cochleae; this process is continuous with the posteromedial flange and abuts the mastoid region. The mastoid region is a relatively large, wedge-shaped area characterized by bony spicules at its edges. The pars cochlearis does not protrude significantly in a medial direction from the pars canalicularis; the mastoid region extends posterior to the pars canalicularis.
The tympanic bulla of this taxon is present but is relatively loosely attached, often missing in dry skulls. A complete tympanic bulla is indeed present (e.g., AMNH-M 73595) in this taxon. It is not as wide as the promontorium and thus does not completely cover it; however, it does contact the petrosal in several places (fig. 113). The anterior crus of the ectotympanic, which is expanded into a meatal tube, contacts the petrosal as does the posterior crus of the ectotympanic (also forming part of the meatal tube) at small roughened areas on either side of the external
( Perissodactyla, Ceratomorpha ). Scale 5 1 cm.
acoustic meatus. The anterior aspect of the auditory bulla also contacted a small area of the large tegmen tympani. Medially and posteriorly, there was extensive contact with the auditory bulla along the posteromedial flange and the caudal tympanic process.
The dorsomedial (fig. 119) surface of the petrosal is smooth but pocked and sharp along the edges of the bone. The internal acoustic meatus is relatively inconspicuous and reniform with the foramina acusticum superius and inferius set deeply within. These are fully separated by a distinct crista transversa. Starting at the anterior edge of the bone and extending from it, along the ventromedial edge, is the basicapsular groove on the medial side of the bone. Medial to the internal acoustic meatus is a small, circular cochlear aqueduct, which occupies a relatively anterior position on the bone. Superior to the internal acoustic meatus is a flattened tegmen tympani that is oriented perpendicular to the dorsomedial surface of the bone. The prefacial commissure is a rounded ledge of bone, and there is no prefacial commissure fossa. The moderate inflation of the tegmen tympani can also be seen in this view. Posterior to the internal acoustic meatus is a large and very shallow subarcuate fossa; there is no petromastoid canal. Medial to the very shallow subarcuate fossa is the vestibular aqueduct, a small slit tucked deeply into the bone. Posteriorly, the mastoid region tapers slightly, retaining a smooth endocranial surface and terminating in a roughened, cancellous border.
Dorsolaterally (fig. 115), the tegmen tympani and mastoid region can be seen to be roughened and pocked with small holes, but this side of the bone does not contain any clear vascular grooves. The dorsolateral side of the petrosal has a flat to convex shape. The blunt anterior process of the tegmen tympani and the moderate inflation of this area are also visible in this view. As noted above, the ventrolateral tuberosity is absent and the hiatus Fallopii is tucked deep within the anterior process of the tegmen tympani.
Ventromedially (fig. 116), the promontorium is narrow anteriorly and transitions posteriorly to a fan-shaped, expanded mastoid region. The basicapsular groove noted above occupied a relatively dorsal position. Posteriorly, the petrosal mastoid region was exposed in a tall (dorsoventrally oriented) rectangle. The mastoid plate is absent.
As can be seen in anterior view (fig. 117), the hiatus Fallopii is a relatively small hole deep to the knoblike anterior process of the tegmen tympani.
CHARACTER DESCRIPTIONS AND CHARACTER STATE OPTIMIZATIONS
CHARACTERS DESCRIBING THE PETROSAL
In this section I compare the variation the characters coded for the petrosal sample, discuss variation in the taxa under study, and make broader comparisons to other studies of mammalian petrosals. Discussions of polarity and optimization are based on patterns seen on the 20 individual most parsimonious trees used to generate the strict consensus tree (fig. 1; Spaulding et al. [2009]). Characters were derived both from the literature (citations below) and from my observations. All characters are illustrated in the drawings and photographs.
1. Texture of area around internal acoustic meatus: rough (0); smooth (1). The bone surrounding the internal acoustic meatus where cranial nerves VII and VIII enter the petrosal is most commonly smooth and featureless in artiodactylans and outgroups (state 1; e.g., fig. 14). In certain cetaceans, however, specifically the odontocete Monodon monoceros (fig. 93), the mysticete Balaenoptera acutorostrata (fig. 78), and the cetaceamorphans † Basilosaurus isis (fig. 99) and † Dorudon atrox (fig. 104), this area is roughened, sometimes with jagged spicules of bone (state 0). The condition is most pronounced in M. monoceros , which has sharp spurs of bone in this area. In B. acutorostrata a mesh of rough, cancellous bone surrounds the internal acoustic meatus. In the two extinct cetaceamorphans studied, the endocranial surface is simply rough in general, without exhibiting the jutting processes.
Optimization indicates that the presence of roughened bone surrounding the internal acoustic meatus is convergent in Monodon and Balaenoptera . Appearance of the smooth area around the internal acoustic meatus is a synapomorphy of the clade defined by the common ancestor of Delphinidae and Physeteridae . A roughened internal acoustic meatus is present in the two most highly nested fossil stem taxa to crown Cetacea . Because this character has not been scored in other more basal cetaceamorphans (e.g., † Ambulocetus , † Indohyus ), however, it is ambiguous as to what point on the cetacean stem it first appears.
2. Subarcuate fossa ( Cifelli, 1982; Novacek, 1986; MacPhee, 1994]); O’Leary and Geisler, 1999; Luo and Gingerich, 1999; Geisler, 2001; Thewissen et al., [2001]): present (0); absent (1). The subarcuate fossa is a variably present depression on the endocranial surface of the pars canalicularis of the petrosal. The anterior semicircular canal (also referred to as the ‘‘superior’’ semicircular canal [ Gannon et al., 1988]) underlies the medial rim of the ostium of the subarcuate fossa ( Gannon et al., 1988; Wible et al., 1995). As noted above, the soft tissue structure housed in the subarcuate fossa is the paraflocculus of the cerebellum (also called the petrosal lobule of the cerebellar paraflocculus; Gannon et al., 1988), and the subarcuate fossa is sometimes referred to as the subfloccular fossa ( Fordyce, 1994: 161) or the cerebellar fossa ( Mead and Fordyce, 2009:114, 129).
Although this character has been used frequently in cladistic studies of mammals, its variation has not been investigated in depth in ungulates. Previous cladistic treatments of the subarcuate fossa have varied considerably; for example, MacPhee (1994: 172) described this character as ‘‘subarcuate fossa: (0) deep; (1) very shallow or absent.’’ This type of coding, however, does not permit cladistic grouping on the basis of the presence of the fossa (regardless of its shape, which varies greatly in ungulates) and was not followed here. The presence and relative depth of the fossa are scored here as separate characters.
This character groups taxa that have any kind of subarcuate fossa. This fossa is typically a depression with a diameter that is at least the size of the internal acoustic meatus, but that may vary greatly in depth (see character 3). I scored as ‘‘present’’ any depression, even a subtle one (e.g., figs. 5, 14), in contrast to ‘‘absent,’’ which I considered the case when the area was convex (e.g., cetaceans).
The fossa lies posterior and slightly dorsolateral to the internal acoustic meatus. It is often separated from the internal acoustic meatus by a ridge, which can be a subtle or distinct convexity, or sometimes a fossa (e.g., † Elomeryx armatus ; fig. 68). Some kind of subarcuate fossa is present (albeit in greatly varying depths) in all taxa examined here except living (e.g., figs. 78, 83) and extinct cetaceamorphans (e.g., figs. 99, 104) that could be sampled, hippopotamids (figs. 44, 49), and Orycteropus afer . The absence of this structure is very clear in extant odontocetes (e.g., fig. 86). The petrosals of these taxa are convex posterior to the internal acoustic meatus and the morphology differs distinctly from the presence of a fossa, even a shallow one. In Balaenoptera acutorostrata (fig. 78) there is little space posterior to the internal acoustic meatus and no depression that is homologous to the subarcuate fossa. The proximal end of the mastoid region in this taxon has a large fossa, which most likely does not correspond to the subarcuate fossa because the bone in this region is too thin to have housed the semicircular canals.
Both hippopotamids (figs. 44, 49) have an internal acoustic meatus that is followed posteriorly by a distinct convexity. The specific area homologous to the subarcuate fossa is most easily localized by identifying the petromastoid canal, which is often near the center of the subarcuate fossa. No shallow depression approximating the size of the internal acoustic meatus surrounds the petromastoid canal; thus, the subarcuate fossa is considered absent in these two taxa. This part of the petrosal is generally contiguous with the enormous, adjacent prefacial commissure fossa present in hippopotamids.
Luo and Gingerich (1999) noted that the size of the subarcuate fossa is not necessarily related to the anatomy of the underlying semicircular canals, and indeed the variability seen here in the subarcuate fossa depth may exceed the variability in the underlying shape of the semicircular canals. The relationship of theses structures warrants further study using both CT scans and surface anatomy so that direct comparisons of these structures can be made.
When optimized, the absence of a subarcuate fossa is a synapomorphy of the common ancestor of the crown clades Cetacea and Hippopotamidae .
3. Subarcuate fossa shape: deep (0); shallow (1), inapplicable in taxa lacking the subarcuate fossa. Although always approximately as wide as the internal acoustic meatus, the subarcuate fossa varies dramatically in depth among the taxa examined here, from a deep pit to a very shallow depression. Deep subarcuate fossae have been described and well illustrated, for example, in Cretaceous eutherians (e.g., † Zalambdalestes, Wible et al., 2004 ) and the ‘‘insectivoran’’ † Leptictis dakotensis (Novacek, 1986) . † Protungulatum sp. (fig. 5) also exhibits the deep condition.
For artiodactylans, however, the fossa is typically much more shallow but depth does vary among both basal and highly nested taxa. Among extant cetartiodactylans, I have observed that Lama glama (not illustrated) has a deep subarcuate fossa but Camelus dromedarius (fig. 34) does not. Most of the suines have a shallow subarcuate fossa. However, contrary to more generalized discussions of this structure in suids (e.g., Norris, 1999: 747), the deep fossa does occur in some members of this clade; it is present in Babyrousa babyrussa (fig. 61), whose ear region had not previously been studied. Among ruminants, all Pecora studied here have a shallow subarcuate fossa, except the extinct † Leptomeryx (fig. 12). The ruminant Tragulus napu (fig. 10) also has a deep subarcuate fossa.
Several other extinct artiodactylans also have deep subarcuate fossae: † Cainotherium ( Hürzeler, 1936) , † Poebrotherium sp. (fig. 39), † Protoceras cele r, and † Perchoerus sp. In other extinct taxa such as the entelodont † Archaeotherium sp. (fig. 76), the anthracothere † Elomeryx armatus (fig. 68), and † Merycoidodon (fig. 74), the fossa is shallow, as it is in both † mesonychians studied, † Dissacus praenuntius (fig. 106) and † Mesonyx obtusidens (fig. 111).
Among outgroups, † Hyopsodus sp. has a deep subarcuate fossa ( Cifelli, 1982), but the perissodactylans (figs. 114, 119) have shallow subarcuate fossae.
Optimization indicates that the shallow subarcuate fossa is a condition that characterized the common ancestor of artiodactylans and perissodactylans and was the primitive condition for Artiodactyla . However, within Artiodactlya are eight taxa that have the deep condition. Homoplasy in this structure occurs across the artiodactylan tree and is associated with all major subclades ( Camelidae , Suina, Ruminantia) where the fossa is present. These results are in general agreement with observations by Norris (1999: 747) that the subarcuate fossa of artiodactylans is generally shallow. Joeckel and Stavas (1996, drawing on Webb and Taylor, 1980) wrote that a character of Pecora (fig. 1) is to have a reduced subarcuate fossa, which here would be equivalent to state 1. This is corroborated as a condition of extant pecorans examined here in the trees of Spaulding et al. (2009), however, it is ambiguous as to whether or not it is a primitive retention. Novacek (1993: 468; see also Novacek, 1986; Novacek and Wyss, 1986), who studied this feature across Mammalia, hypothesized that the ‘‘reduction or loss of the subarcuate fossa occurred independently several times within therians,’’ and that in ungulates, in particular, this structure is often poorly developed. My results corroborate the hypothesis that the relative depth of the subarcuate fossa exhibits homoplasy.
4. Petromastoid canal in subarcuate fossa ( Gannon et al., 1988): absent (0); present (1). Gannon et al. (1988) observed that in certain taxa (e.g., humans), the subarcuate fossa is largely, but not completely, closed. In fetal humans they observed that a large pit similar to a deep subarcuate fossa is present; however, in the adult human, the fossa is absent and there remains only a narrow channel in its place, the petromastoid canal. The petromastoid canal is an attenuated communication between the posterior cranial fossa and the mastoid air cells ( Gannon et al., 1988: fig. 4g). Gannon et al. (1988: 160) stated that they considered the petromastoid canal to be homologous to ‘‘vascular foramina in the walls of the subarcuate fossa.’’ By this definition, this feature can coexist with the subarcuate fossa. Mazzoni (1970) reported that the petromastoid canal transmits the petromastoid artery, a branch of the inferior cerebellar artery.
The petromastoid canal is present in a variety of taxa, including both hippopotamids (figs. 44, 49), four suids ( Sus scrofa [fig. 53], Hylochoerus meinertzhageni [fig. 59], Babyrousa babyrussa [fig. 61], and Potamochoerus porcus [fig. 51), the camelid Camelus dromedarius (fig. 34), the extinct artiodactylans, † Protoceras celer (fig. 41), † Merycoidodon sp. (fig. 74), and also, the † mesonychian † Dissacus praenuntius (fig. 106; see also Luo and Gingerich, 1999: fig. 3B). Although the petromastoid canal appears to occur most frequently when the subarcuate fossa is also shallow, the two characters are anatomically distinct. B. babyrussa (see also Canis familiaris , personal observation) is an example of a taxon in which the petromastoid canal coexists with a deep subarcuate fossa. Also, the petromastoid canal is present in hippopotamids even though they lack a subarcuate fossa.
This structure appeared convergently several times within Artiodactyla but never occurs in crown Cetacea . It is a separate synapomorphy for each of the clades Suidae and Hippopotamidae (the latter being an ambiguous synapomorphy due to lack of data for stem fossil taxa); however, because these clades are not sister taxa or particularly closely related in any of the most parsimonious trees, the petromastoid canal appears to be independently derived in each clade, and at least seven times across the tree as a whole.
5. Hiatus Fallopii distinct anterior hole ( Luo and Gingerich, 1999): present (0); absent (1). As described in early sections of this paper, this opening transmits the greater petrosal nerve. The presence of a distinct foramen (regardless of its size) at the anterior aspect of the expanded tegmen tympani is a regularly occurring feature of most noncetacean artiodactylans (e.g., fig. 56) and outgroups (fig. 117). It differs from the condition in cetaceamorphans that could be examined here, where a distinctive hole is often absent. This character was only scored when the petrosal could be examined in isolation in anterior view to ensure clear comparison of this area (e.g., fig. 91). This character is, therefore, more a description of the presence/absence of the distinct type of foramen than it is an exhaustive study of the course of the greater petrosal nerve in cetaceans. Dissection-based studies, as suggested by Mead and Fordyce (2009), along with use of CT scanning to document internal connections between openings in the petrosal will be important for a full description of how the greater petrosal nerve travels in cetaceans. For example, regarding the fossil platanistoid, † Waipatia, Fordyce (1994: 158 , 160) suggested tentatively that this taxon had a ‘‘supplementary opening (for the greater petrosal nerve?),’’ a small hole at the anterior aspect of the internal acoustic meatus. Several of the cetaceans examined here do have one or more smallsized and irregularly spaced holes on the dorsal surface of the expanded tegmen tympani. These should be particular targets for examination in dissection or CT scanning. When the hiatus Fallopii is absent, the greater petrosal nerve has sometimes been inferred to have branched directly from the facial nerve in the tympanic cavity to course anteriorly (e.g., Ekdale et al., 2004: 172).
Optimizations indicate that the absence of a distinct hole at the anterior tegmen tympani is a feature of cetaceans and their stem taxa (through † Dorudon ) only. Due to a lack of information on this part of the petrosal in many cetacean stem taxa, the node where this loss appears is ambiguous.
6. Hiatus Fallopii size, if distinct anterior hole: small (0); large (1), inapplicable in taxa, such as cetaceans, where the homolog to this opening has not been established. I have observed that certain taxa have a very large hiatus Fallopii relative to the rest of the petrosal. The taxa that have a large hiatus Fallopii are the pecoran Bos taurus (fig. 17), both extant hippopotamid species (fig. 47), three suids, Sus scrofa (fig. 56), Babyrousa babyrussa (fig. 56), and Potamochoerus porcus (fig. 47), the † entelodont † Archaeotherium sp. (fig. 71), and the † anthracotheres † Elomeryx (fig. 71) and † Bothriogenys (not illustrated). This contrasts with other taxa studied where this foramen is a pin-sized slit or hole (e.g., fig. 17). I am not aware of studies that compare the relative size of the greater petrosal nerve in taxa where the hole is small versus large, or the functional implications of this size variation.
Optimizations indicate that at the base of the artiodactylan tree and among outgroup taxa have a relatively small hiatus Fallopii. However, among suids, hippopotamids, ru- minants and ruminantiamorphans, there are several examples of convergent appearances of the large hiatus Fallopii. The large hiatus Fallopii is a synapomorphy of suids and is completely convergent in Bos taurus . The condition of the hiatus Fallopii in the common ancestor of ruminants and hippopotamids is ambiguous. A large hiatus Fallopii is presently an ambiguous synapomorphy of Cetancodontamorpha.
7. Prefacial commissure fossa of the petrosal ( Luo and Marsh, 1996; Luo and Gingerich, 1999: char. 4, synonym, suprameatal fossa): absent (convex, flat, or bump in suprameatal region) (0); present (1). This feature is a large concavity on the endocranial surface of the petrosal that lies immediately superior (dorsal) to the internal acoustic meatus. In many taxa the suprameatal area consists of a relatively simple flat or convex bar of bone that meets the dorsolateral surface of the petrosal at a right angle (e.g., figs. 2A, 17). In some artiodactylan taxa, however, this area is a broad excavation that occupies much of the dorsomedial surface of the tegmen tympani (e.g., fig. 102). Luo and Gingerich (1999: 52) called it a ‘‘concave area on the medial side of the tegmen tympani.’’ Luo and Gingerich (1999: 26) were the first to observe the similarity in this structure between † mesonychians (e.g., † Dissacus ) and cetaceamorphans.
The taxa in this sample with a prefacial commissure fossa are: both hippopotamids (figs. 44, 47, 49), the † mesonychians † Dissacus praenuntius , † Mesonyx obtusidens (figs. 106, 109, 111), and † Harpagolestes orientalis , cetaceamorphans (fig. 102) † Dorudon and † Basilosaurus , † Pakicetidae , and † Indohyus , along with the † anthracothere † Bothriogenys .
Presence of a prefacial commissure fossa is a synapomorphy of Cetancodonta (the clade that includes hippopotamids and cetaceans). This feature reverses to absent at the base of crown Cetacea and is absent in all extant cetaceans sampled here. Presence of the prefacial commissure in several † mesonychids and in † Bothriogenys are examples of independent origins of this feature. Presence of the feature is an ambiguous synapomorphy of † Mesonychidae . That this character state is absent in more derived cetaceans (e.g., figs. 86, 91) but present in fossil stem taxa to Cetacea underscores the significance of fossils for establishing a hippopotamid-cetacean link.
8. Shape of the dorsolateral tegmen tympani ( Luo and Gingerich, 1999; fig. 121): flat (0); pronounced convexity (1). The shape of the tegmen tympani varies among ungulates and cetaceans, sometimes in a subtle fashion. The different shapes can generally be broken into two groups: a convex (‘‘barrel,’’ according to Geisler and Luo, 1998: 201, fig. 47) shape, and a flat shape (e.g., figs. 2, 17). The character in general can be best observed in anterior views of the petrosal, indicating the right angle made between the dorsomedial and dorsolateral surfaces of the bone. This character is essentially identical to character 12 of Luo and Gingerich (1999: 54), only they described it as the ‘‘lateral surface of [the] anterior process [of the tegmen tympani].’’ Note, however, that in many taxa studied here this surface is actually the dorsaldorsolateral petrosal surface (fig. 2).
Luo and Gingerich (1999: 54) made the important observation that a convex dorsolateral tegmen tympani is shared by † mesonychids and cetaceans. Here; however, I show that the feature also appears elsewhere among noncetacean artiodactylans and that it exhibits homoplasy. The less common convex shape is present in extant (e.g., fig. 81) and fossil stem cetaceans (fig. 102; for which the petrosal was preserved in isolation for study here), † mesonychids (fig. 109), hippopotamids (fig. 47), the † anthracothere † Bothriogenys , and in † Agriochoerus .
A pronounced convexity to the tegmen tympani is a synapomorphy of Cetancodonta and occurs in all members of this clade for which data were available. The appearances of this feature in † mesonychids, † Bothriogenys , and † Agriochoerus optimize as independent origins.
9. Inflation of the tegmen tympani ( Luo and Gingerich, 1999): absent (0); present (1). The width of the tegmen tympani (relative to the promontorium) has been identified as an important character distinguishing ungulates from other mammals ( Novacek and Wyss, 1986; Geisler and Luo, 1998), and cetaceans exhibit hypertrophy of this area ( Luo and Gingerich, 1999). The only ungulate taxon in the matrix for which inflation is currently scored as absent is the extinct † Protungulatum (fig. 3), in which the tegmen tympani is merely a thin sheet of bone dorsolateral to the fossa for the tensor tympani. In contrast (in tympanic, endocranial, or anterior views), in the petrosals of artiodactylans and perissodactylans, the tegmen tympani is a broad bar of bone that accounts for as much as onethird to one-half the size of the promontorium in ventrolateral view. In certain taxa where I have not been able to examine a petrosal isolated from other bones of the skull (especially the squamosal) this character has been scored as a ‘‘?’’ because assessment of the size of the tegmen tympani in situ was too uncertain.
Cifelli (1982: 795) stated that the tegmen tympani was thin and uninflated in artiodactylans based on information in † Homacodon , and that this distinguished artiodactylans from other ‘‘higher ungulates.’’ Luo and Gingerich (1999: 51), in contrast, stated that † condylarths and Early Tertiary artiodactylans do show some inflation of the tegmen tympani; I concur.
Coombs and Coombs (1982) stated that † Diacodexis , † Helohyus , and † Gobiohyus each had an uninflated tegmen tympani, and they specifically contrasted this condition with that of the † condylarth † Hyopsodus , which Cifelli (1982) described as having an inflated and pneumatized tegmen tympani. Here I have scored these early Eocene artiodactylans (not figured here) as ‘‘?’’ for this character because the petrosal is in situ making the size of the tegmen tympani difficult to assess. I do not agree with the statement by Coombs and Coombs (1982; see also Cifelli, 1982) that ‘‘artiodactyls … are primitive in lacking the inflated pneumatized tegmen tympani of other ungulate groups.’’
Inflation of the tegmen tympani was present in the common ancestor of the clade including Artiodactylamorpha + † Mesonychia .
10. Degree of inflation of the tegmen tympani ( Luo and Gingerich, 1999; see also Novacek and Wyss, 1986): hyperinflation (transverse width greater than or equal to width of the promontorium) (0); moderate inflation (one-third the width of the promontorium or less) (1). This feature is best viewed and compared in anterior and dorsolateral views, and, as noted by Luo and Gingerich (1999: 52), this character reflects relative size of the tegmen tympani in comparison to the transverse width of the promontorium. Among living mammals the ‘‘hyperinflation’’ condition is found in both hippopotamid species (figs. 42, 48), as well as in cetaceans except for Pontoporia blainvillei . The presence of this feature in hippopotamids has not previously been described. Luo and Gingerich (1999: 23) described the † mesonychian † Dissacus praenuntius as having a tegmen tympani that is ‘‘greatly inflated, making it much larger than those of other extant and archaic ungulates’’; however, comparisons here actually indicate that this area is no more inflated in † Dissacus praenuntius (fig. 105) than in many other noncetacean artiodactylans (e.g., figs. 21, 52). † Mesonyx , however, has been grouped in the ‘‘hyperinflation’’ state because of the width of the tegmen tympani relative to the promontorium; greater relative inflation has occurred in hippopotamids and cetaceans. Among other fossils, both extinct cetaceans † Dorudon and † Basilosaurus (figs. 97, 103) exhibit the hyperinflated state (for † Pakicetidae no isolated petrosal was available to score this feature). Finally, the † anthracothere † Bothriogenys also exhibits this condition.
Hyperinflation is a synapomorphy of Cetandonta even though it does show some homoplasy in the clade (reversing in Pontoporia ). The hyperinflation in † Bothriogenys and † Mesonyx are independent derivations.
Novacek and Wyss (1986: table 1) suggested that presence of an ‘‘inflated, pneumatized tegmen tympani’’ provided tentative support for a clade of perissodactylans, cetaceans, hyracoids, sirenians, and proboscideans, to the exclusion of artiodactylans. These authors drew on remarks in earlier papers ( Coombs and Coombs, 1982: 232) that artiodactylans lacked this feature. However, based on data collected here, an inflated tegmen tympani is present in artiodactylans, making it unlikely that this character would support the clade proposed by Novacek and Wyss (1986) to the exclusion of artiodactylans.
11. Anterior process of the tegmen tympani (O’Leary and Geisler, 1999; Luo and Gingerich, 1999; Geisler, 2001; Thewissen et al., 2001): absent (0); present (1). Luo and Gingerich (1999: 54) defined the anterior process as ‘‘the additional portion of the tegmen tympani anterior to both the vascular groove and the gap between the tegmen and the pars cochlearis.’’ Use of the term ‘‘anterior process’’ of the petrosal by Luo and Gingerich (199: fig. 2) for noncetaceans was a precedent for identifying this structure broadly among mammals. In this study I have included many more noncetacean artiodactylans and other ungulates than in Luo and Gingerich (1999), and I have been able to reexamine this character more broadly. Because the presence of a vascular groove is variable, it seems appropriate to recast the definition of the anterior process. Here I define it as the anteriormost aspect of the tegmen tympani (bordered posteriorly by the external acoustic meatus). In my observation, contra Luo and Gingerich (1999), the size of this structure in † mesonychians (figs. 105, 110) is not significantly larger than the size of it in many noncetacean artiodactylans (e.g., figs. 28, 62). The presence of the anterior process, therefore, is not exclusive to † mesonychians and cetaceans.
I find that the anterior process is also present in all artiodactylan and perissodactylan extant and fossil taxa (for which it could be scored) studied here, but not in † Protungulatum sp. (fig. 3). The presence of this feature is a synapomorphy of the large clade that is the common ancestor of Artiodactyla + † mesonychians.
12. Size of the anterior process of the tegmen tympani ( Kellogg, 1936; Heyning, 1989; Luo and Gingerich, 1999; Messenger and McGuire, 1998; Geisler and Luo, 1998): small, does not extend anterior to the promontorium (0); large, extends anterior to promontorium (1).
As noted above, I have observed in the taxa studied here that, contra Luo and Gingerich (1999), many artiodactylans have an anterior process of the tegmen tympani that is of comparable size to that in † mesonychids (e.g., † Mesonyx obtusidens [fig. 110]). Those authors suggested that in † mesonychids the anterior process had a ‘‘striking resemblance to the anterior process of … cetacean petrosals’’ ( Luo and Gingerich, 1999: 23). In neither † mesonychians nor noncetacean artiodactylans, however, does the anterior process extend anterior to the promontorium, so the two groups are scored similarly as having a small anterior process. The large state is present in living and extinct cetaceamorphans, and also in Equus caballus (fig. 112). Although not as enormously hypertrophied as in some cetaceans, the enlargement in E. caballus fits best with state 1.
The large state is an ambiguous synapomorphy of the common ancestor of Delphinidae (fig. 87) and † Dorudon atrox (fig. 103), because the condition is not known in certain stem taxa to whales. The occurrence of this feature in Equus caballus (fig. 112) is a homoplasy. A small anterior process is the primitive and widespread condition among the taxa studied here.
Heyning (1989: 49) described the anterior process as reduced in extant odontocetes. I have observed considerable variation in the size of this character among cetaceans; however, relative to artiodactylans, all cetaceans have large anterior processes of the tegmen tympani. My coding is not sensitive to variations within odontocetes, and I did not assess whether within cetaceans the anterior process is reduced from a larger precursor or simply derived as moderately enlarged anterior to the tegmen tympani. The anterior process, however, is quite hypertrophied in the mysticete Balaenoptera acutorostrata (fig. 77).
13. Apex of the anterior process of tegmen tympani ( Luo and Gingerich, 1999): pointed (0); blunt (1). The anterior end of the tegmen tympani can be broadly categorized into two major shapes. I have found that the process is pointed in all extant cetaceans examined. Among noncetacean artiodactylans the process is typically blunt, a condition that does show some homoplasy. The pointed condition occurs in some extant suids (e.g., fig. 50) and in the stem ruminant † Cainotherium .
Appearance of the pointed condition is a feature of crown cetaceans and their stem taxon † Basilosaurus ; however, it is ambiguous as to where exactly this appears in Cetaceamorpha. While a more blunt shape of the anterior process is the primitive and more commonly held condition in the taxa studied here, modification to a pointed condition exhibits homoplasy in several parts of the tree, including among suines, stem camelids, and even the perissodactylan Equus .
14. Vascular groove on the lateral surface of the anterior process of the tegmen tympani ( Luo and Gingerich, 1999): present (0); absent (1). A vascular groove that runs either anteroposteriorly or mediolaterally may appear on the surface of the petrosal that contacts the squamosal. Several authors have described such furrows, inferred their contents, and used their morphology for cladistic characters in artiodactylans, particularly in fossil taxa (e.g., Fordyce, 1994; Geisler and Luo, 1998; Luo and Gingerich, 1999; Geisler and Sanders, 2003). As noted above, I describe no new soft tissue information in this paper and thus do not expand on these inferences, particularly because six taxa with these furrows are extant and should be dissected to reveal the detailed soft tissue relationships with this structure. I am homologizing the presence of a groove at a general level.
Giraffa , Kogia , Ziphius , † Dorudon , † Basilosaurus , † Mesonyx , and † Dissacus zanabazari each have a more mediolaterally oriented furrow, of differing relative lengths and breadths. In contrast, Tursiops , Hippopotamus , Choeropsis , and † Leptomeryx each has a more anteroposteriorly oriented furrow. Dissections of the extant taxa could confirm whether the groove is being redirected but has similar contents or if the contents vary.
Luo and Gingerich (1999: 64) observed that this groove was shared between † mesonychids and cetaceans. Optimization on the cladogram of Spaulding et al. (2009) indicates that the presence of a groove is a synapomorphy of Cetancodonta. The appearance of the groove in † mesonychids is a parallel origin, and the common ancestor of † mesonychids and cetaceans did not share this feature.
15. Distinct fossa for the head of the malleus in epitympanic recess ( Coombs and Coombs, 1982; Geisler and Luo, 1998; Luo and Gingerich, 1999): absent (0); present (1). The epitympanic recess and the relative positions of the ossicles can be difficult to assess in dry skulls because the ossicles are almost never in position. At the same time few dissection-based studies exist that illus- trate the tympanic cavity of extant ungulates dissected in ventral view with ossicles and soft tissues in place. This character is intended to distinguish between two extremes in variation: the relatively large and distinct fossa for the head of the malleus in cetaceans versus the very small pit that is often present in noncetaceans. The character does not distinguish variation among those taxa that share a small indistinct fossa for the head of the malleus. In taxa for which the fossa for the head of the malleus is indistinct, the epitympanic recess may consist of only a single concave surface.
The distinct fossa for the head of the malleus in cetaceans lies anterolateral to the fenestra vestibuli (see Mead and Fordyce [2009: fig. 25v, ‘‘mallear fossa’’] for Tursiops ). It tends to be displaced ventrolaterally from the secondary facial foramen (i.e., the ventral exit of cranial nerve VII is a deeper structure than the epitympanic recess and its specific parts). Luo and Gingerich (1999: 53; see also Geisler and Luo, 1996) stated that these fossae were absent in mysticetes, an observation echoed by Mead and Fordyce (2009). I have observed, however, that the fossa for the head of the malleus is clearly present in the mysticete Balaenoptera acutorostrata (fig. 77). Note also that in cetaceans the large and distinct fossa for the head of the malleus is found in the position that in many other species examined here is occupied by the fossa for the tensor tympani muscle.
The presence of a clear fossa for the head of malleus is an ambiguous synapomorphy of the clade consisting of the common ancestor of Delphinidae ( Tursiops truncatus ) and † Dorudon atrox (fig. 1), and because this feature cannot be scored in other cetaceamorphans, it is not clear exactly where it appeared.
16. Ventrolateral tuberosity ( Luo and Gingerich [1999]): absent (0); present (1). This is a ventrally projecting protuberance on the lateral surface of the petrosal. It typically appears just anterior to the external acoustic meatus and assumes a variety of shapes. This process has been described under varying names by different authors (see discussions by Fordyce, 1994: 161; Luo and Gingerich, 1999). The ventrolateral tuberosity equals the ‘‘lateral tuberosity’’ of Barnes (1978), the ‘‘ventral tuberosity’’ of Muizon (1987), and the ‘‘ventrolateral tuberosity’’ of Luo and Marsh (1996: 335, see for discussion of its presence in archaeocetes and odontocetes). I am using a broad definition of this term, but see also discussions in Geisler and Luo (1996) and Geisler and Sanders (2003).
Luo and Gingerich (1999) described this state as present only in taxa with a very inflated tegmen tympani, namely extinct cetaceamorphans studied and certain odontocetes. I have found, however, that there is a homologous structure in a variety of ungulates, many of which do not have a hyperinflated tegmen tympani.
I find that the presence of this structure exhibits a large degree of homoplasy. In † mesonychids it is present in † Dissacus praenuntius but not in † Mesonyx obtusidens (fig. 107) (see also Luo and Gingerich, 1999: 52–53). I have also found that this feature is not uniformly well developed in odontocetes (contrast figs. 92 and 94), and that it is variably present in some living and extinct noncetacean artiodactylans including certain ruminants (e.g., figs. 18, 28) and suines (figs. 52, 62). Corroborating Luo and Gingerich (1999), I found this structure to be absent in the one mysticete sampled (fig. 77); however, see the discussion in Geisler and Sanders (2003: 53–54), who found this to be present in mysticetes.
No particular pattern emerges from study of the optimization of this character. The tuberosity is present in the common ancestor of artiodactylans and perissodactylans but primitively so.
17. Shape of ventrolateral tuberosity: knob (0); spike (1). Although a variety of taxa have a projection of some sort on the ventral petrosal just anterior to the external acoustic meatus, this projection varies in shape from a spike to a more rounded bump. This process typically abuts the ectotympanic ring if it is expanded into a tube and if there is a contact between it and the petrosal (e.g., Odocoileus virginianus , fig. 28). In crown ruminants and their stem taxa that have this process it is spike-shaped. Among hippopotamids, this structure varies in shape: bump-like in Choeropsis and spike-shaped in Hippopotamus (figs. 42, 45). Among suines (figs. 57, 60, 62) and fossil relatives that have this feature (e.g., fig. 54), some taxa have the knob shape and others have a spike shape.
Optimizations indicate that the bump/ knob shape is the primitive condition; however, transition to a spike shape exhibits homoplasy when it appears in hippopotamids, suines, and ruminants.
18. Fossa for the tensor tympani muscle round or oval depression (O’Leary and Geisler, 1999; Luo and Gingerich, 1999; Geisler, 2001; Thewissen et al., 2001): present (0); absent (1). Just anterior or anteromedial to the secondary facial foramen on the petrosal there is sometimes a pit of varying shapes and sizes for the tensor tympani muscle. In life, this muscle extends from the ventral surface of the petrosal, and its tendon inserts on the muscular process of the malleus ( Evans and Christensen, 1979: 1064). Evans and Christensen (1979: 1067) described this as a ‘‘spherical’’ muscle, the belly of which lies in the fossa on the ventral petrosal surface. In Canis they noted that contraction of this muscle tends to pull the tympanic membrane medially, increasing tension on the membrane. How much of the fossa is actually filled by muscle during life is poorly documented for the extant taxa described here.
I have found that † Protungulatum (fig. 3) lacks a distinct tensor tympani fossa. Wible et al. (1995) observed that although there is a distinct bony fossa for the tensor tympani in taxa such as † multituberculates, many Mesozoic mammals (e.g., the Khoobur petrosal) lack a distinct bony fossa for this muscle. In the Cretaceous eutherian † Zalambdalestes (Wible et al., 2004) , for example, the origin of the tensor tympani was most likely a faint excavation on the lateral part of the promontorium (specifically on the ‘‘anterolateral flange’’ [Wible et al., 2004: 73]). A relatively subtle fossa for the tensor tympani also characterizes the insectivoran fossil † Leptictis dakotensis (Novacek, 1986) . Thewissen (1985) remarked that this fossa is absent in perissodactylans; however, I have observed it in both Equus caballus (fig. 112) and Tapirus terrestris (fig. 118).
I have found that this fossa is absent in extant cetaceans and the extinct cetaceamorphans † Dorudon and † Basilosaurus (figs. 97, 103). In cetaceans the area for this muscle is often described as a ‘‘groove’’ ( Luo and Gingerich, 1999: 57) that does not resemble the pit-shaped fossa seen in many other taxa. The fossa for the tensor tympani is present in all other taxa in which this region was preserved, including the extinct cetaceamorphan † Pakicetus (it is not currently scored in † Indohyus ). Nonetheless, both this muscle and the stapedius muscle are reported to be present in extant cetaceans ( Fraser and Purves, 1960). Oelschläger (1990: 150) stated that in Tursiops truncatus the tensor tympani attaches in a ‘‘minute shallow ovoid fovea … [and] the reduction of the processus muscularis does not imply a reduction of the muscle proper.’’ He illustrated the groove for the tensor tympani in T. truncatus (but not Inia or Pontoporia ) on p. 149. Oelschläger (1990: 151) also stated that the fossae for these muscles were well developed in extinct cetaceamorphans (namely, † Dorudon atrox ). Other than in † Pakicetus (figs. 96–97, 103), however, I have not observed this fossa in the extinct cetaceamorphans studied here (which is consistent with findings of Luo and Gingerich, 1999).
Oelschläger (1987) stated that Gingerich et al. (1983) erred when they argued that † Pakicetus was primitive in having a large tensor tympani muscle, inferred from the presence of a large fossa for the tensor tympani. Oelschläger (1987: 680) noted that ‘‘this muscle [the tensor tympani] is well developed even in advanced odontocetes … so that in this respect † Pakicetus does not show the generalized condition.’’ We have no direct evidence of the actual size of the muscle in the extinct † Pakicetus ; however, its fossa for the tensor tympani much more closely resembles the more generalized condition of noncetacean artiodactylans than it does the condition in cetaceans, consistent with Gingerich et al. (1983).
Relevant to interpreting bony anatomy, Veggetti et al. (1982: fig. 10) described variation in the tensor tympani muscle, both the gross morphology and the muscle fiber types, of several of the taxa also examined here (horse, cow, pig, sheep, and dog). They described this muscle as having a tympanic portion and a tubal portion and identified three distinct gross types of tensors tympani. The types are: ‘‘club-shaped’’ (with a spheroidal tympanic portion and an elongate spindle-shaped muscular tubal portion), present in the sheep; ‘‘spheroidal-shaped’’ (with a large spheroidal tympanic portion and no significant muscular tubal portion), present in the dog, cow, and pig; and ‘‘spindleshaped’’ (with a larger tubal than tympanic portion, and a tympanic portion that is less spheroidal), present in the horse. These authors noted that, based on their analysis of the presence and type of muscle fibers, the tensor tympani in the cow and the pig ‘‘performs more the function of a ligament than a muscle (p. 350).’’ The relationship between muscular differences and adjacent osteological variability are, however, poorly documented. Overall, all of the taxa that Veggetti et al. (1982) described have roundoval fossae of varying sizes.
Presence of this fossa is the primitive and widespread condition for the taxa studied here. The loss of the fossa occurs in the common ancestor of † Dorudon and Tursiops ; however, this is an ambiguous synapomorphy of this clade because other stem taxa to Cetacea are currently not scored for this character. The lack of the fossa in † Protungulatum is an independent loss.
19. Fossa for tensor tympani extends into an excavated portion of the tegmen tympani: absent (0); present (1). In certain taxa the tensor tympani muscle fossa extended laterally into the petrosal, deep to the tegmen tympani, excavating a large pit. The presence of this feature is highly homoplastic, appearing in some ruminants, some suines, perissodactylans, the † mesonychid † Harpagolestes , and in several extinct noncetacean artiodactylans.
Optimization of this character indicates that it is highly homoplastic. Presence of a large fossa that excavates the tegmen tympanic is an ambiguous synapomorphy of artiodactylans and perissodactylans. There is no excavation of this area in Cetaceamorpha.
20. Transpromontorial sulcus ( Cifelli, 1982; Thewissen and Domning, 1992; Luo and Gingerich, 1999; O’Leary and Geisler, 1999; Geisler, 2001; Thewissen et al., 2001): present (0); absent (1). This character records the presence of a groove across the middle of the ventral surface of the promontorium directed primarily anteroposteriorly (however, positions exhibit some variation in the sample here). A transpromontorial sulcus is often attributed to the presence of an internal carotid artery that crosses the ventral surface of the promontorium; however, internal carotid nerves and the tympanic plexus may also leave grooves on the promontorium. Because I present osteological data only, including descriptions of many extant taxa for which middle ear dissections have not been published, I refrain from explicitly inferring what soft tissue structures occupied this groove and simply describe and illustrate the presence of a groove. Another complication for inferring soft tissue that has long been recognized is that when the internal carotid artery travels on the ventral surface of the promontorium, it may or may not leave a groove in its path (Wible, 1986). Thus, the artery may be present even if the groove is absent, and grooves may be present if the artery is absent because the internal carotid nerve or nerves that follow a similar course across the promontorium remain. That said, what is known of extant artiodactylans indicates that the extant forms lack a transpromontorial portion to the internal carotid artery (see Wible [1984] for an excellent summary). Thus, it would be parsimonious to predict, at present, that grooves on the promontoria of artiodactylans may have been made by the internal carotid nerve; however, this should be confirmed by dissection.
Novacek and Wyss (1986: 278) stated that the transpromontorial sulcus was ‘‘lost in all ungulate lineages save primitive artiodactyls ( Coombs and Coombs, 1982).’’ This conclusion is not supported by the data presented here because several highly nested artiodactylans have a transpromontorial groove (e.g., fig. 18). These authors also noted that loss of the sulcus shows homoplasy throughout Mammalia, which is supported in this study. Geisler and Luo (1998: 195) also described the transpromontorial artery in artiodactylans and stated that presence of the internal carotid artery was primitive for Paraxonia (‘‘artiodactylans’’ + † mesonychians + cetaceans).
Some kind of sulcus on the promontorium is present in † Protungulatum and Orycteropus . For noncetacean artiodactylans a sulcus is present in the ruminants Bos , Giraffa , Antilocapra , Moschus , the hippopotamid Hippopotamus , and the suines Babyroussa, Hylochoerus , and Tayassu . Regarding extinct taxa, the sulcus is present in two † mesonychians, † Mesonyx and † Hapalodectes , along with the artiodactylans † Helohyus , † Agriochoerus , † Diacodexis , and † Elomeryx . Wible (1984; see also Kampen, 1905) mentioned that a transpromontorial sulcus was present in Cervus , but I did not observe this in the specimen studied here.
Absence of the transpromontorial groove is the condition in the common ancestor of artiodactylans and perissodactylans. Within Artiodactyla , however, a transpromontorial groove does reappear at least once in each of the major clades (suines, hippopotamids, ruminants, and camelids), either within the crown clade itself or on its stem or both.
Wible (1986: table 1) reported that ‘‘artiodactylans’’ (based on † Diacodexis , personal commun.) and cetaceans had the primitive mammalian condition for the internal carotid artery: transpromontorial. Here it is interesting that not only is absence of the transpromontorial sulcus primitive for Artiodactyla , it is also the condition common to most members of the clades. It may be that, as suggested by Wible (1986), it is primitive for Artiodactyla to have a transpromontorial internal carotid artery but that in this clade the artery happens not to leave a groove. Alternatively, the artery may be primitively absent. Resolution of this question, as well as the question of whether promontorial grooves in all cases are even made by the internal carotid artery, internal carotid nerve, or another structure (such as the tympanic nerve), requires dissections and soft tissue descriptions for the extant taxa scored followed by inclusion of the soft tissue data as a character separate from the osteological transpromontorial groove in the cladistic analysis and optimization.
21. Double transpromontorial sulci: absent (0); present (1). In some taxa studied here there are two distinct and generally parallel grooves on the promontorium running in a generally anteroposterior direction. MacPhee (1981: 36; see also Novacek, 1986) illustrated the internal carotid nerve wrapping tightly with the transpromontorial internal carotid artery and forming lateral and medial nerve branches ( MacPhee, 1981: 43). These structures associated with the internal carotid artery may also leave grooves on the ventral surface of the promontorium. Other soft tissue candidates for these grooves (depending on how soft tissue structures may vary in position) are branches of the tympanic plexus, which lie on the promontorium of the petrosal supplying the tympanic mucosa ( Evans and Christensen, 1979: 1064). Wilkie (1936: fig. 4) illustrated (but did not discuss extensively) grooves on the ventral promontorium that he stated represented the path of the tympanic nerve.
As noted above, however, in this paper I describe only the osteological features (not reconstructed soft tissues). In some taxa, I have observed two distinct transpromontorial grooves, for example, in Bos taurus (fig. 18), Giraffa camelopardalis (fig. 30), Hippopotamus amphibius (fig. 48), and the extinct artiodactylan, † Elomeryx (fig. 67). The presence of double transpromontorial sulcus is highly homoplastic and was independently derived in almost every case (except in ruminants where the origin of this feature is ambiguous).
22. Sulcus for stapedial artery: present (0); absent (1). A sulcus branching from the transpromontorial sulcus and running toward the fenestra vestibuli is attributed to the stapedial artery (also called the ‘‘proximal stapedial artery’’ by Wible [1987], Geisler and Luo [1998], and Luo and Gingerich [1999]). Geisler and Luo (1998: 176) reviewed the literature on this structure and stated that although this sulcus is present in † Diacodexis specimens from North America (see Geisler and Luo, 1998: fig. 2), it was absent in extant Artiodactyla , Perissodactyla , and Cetacea . I find also that there are no examples of this sulcus in the extant artiodactylans sampled. The sulcus is present in the extinct artiodactylans † Cainotherium , † Diacodexis (North American specimens), and † Helohyus . It is also present in two † mesonychians, † Mesonyx and † Hapalodectes , as well as † Protungulatum .
Optimizations here indicate that absence of this groove is a synapomorphy of artiodactylans and perissodactylans (except in some most parsimonious trees, where unscored fossil stem taxa to this clade render this conclusion ambiguous). Also as noted by Geisler and Luo (1998), it is the most ancient members of the artiodactylan clade that have a sulcus branching from the transpromontorial sulcus; however, in the Spaulding et al. (2009) tree, these ancient taxa (e.g., † Helohyus , † Diacodexis ) are relatively highly nested. This means that despite being present in older members of the clade, the tree does not support the hypothesis that the presence of this structure is primitive for Artiodactyla , Perissodactyla , or their common ancestor.
Cifelli (1982) observed that the stapedial sulcus was absent in the ‘‘ † condylarthran’’ † Hyopsodus . Wible (1987) argued that there had been many independent acquisitions of the stapedial artery within Eutheria, and indeed the tree produced here certainly indicates that there is homoplasy of the bony channel made by this structure in Artiodactyla alone. Wible (1987) also suggested that absence of the stapedial artery might unite Cetacea , Perissodactyla , and other ungulates as a clade, to the exclusion of Artiodactyla , an argument supported in part by the presence of this sulcus in Early Tertiary artiodactylans. Based on the tree of Spaulding et al. (2009), showing that certain Early Tertiary artiodactylans are highly nested, the common ancestor of Artiodactyla is reconstructed most parsimoniously as lacking this structure. Under this scenario, the presence of this structure would not distinguish artiodactylans from other ungulates.
23. Pars cochlearis protrudes ventromedially: absent (0); present (1). This character captures the overall shape variation visible in ventrolateral (tympanic) view. In artiodactylans there are variations in the way the pars cochlearis meets the pars canalicularis on the medial side of the petrosal. In ventrolateral view it can be seen that in some taxa there is an angle between the pars cochlearis and the pars canalicularis (e.g., extant cetaceans [figs. 77, 92]); the mastoid region is offset distolaterally relative to the pars cochlearis. In other artiodactylans the mastoid is not offset from the mediolateral extent of the promontorium (e.g. fig. 40), such that a straight line would demarcate the medial border of both the pars cochlearis and pars canalicularis.
Extant and extinct cetaceans, Hippopotamus and † Mesonyx obtusidens (fig. 110) have a medially protruding pars cochlearis, a character state that varies among noncetacean artiodactylans (compare Ovis [fig. 21] with Bos [fig. 18]). Most ruminants have a medially protruding pars cochlearis and most suines do not.
This character exhibits a great deal of homoplasy, making patterns of support difficult to tease out. In the clade defined as the common ancestor of Artiodactyla and † Protungulatum , the absence of a protruding pars cochlearis is a synapomorphy; however, internal to this clade are many instances of homoplasy to the present condition.
24. Mastoid region size ( Luo and Gingerich, 1999): small (, 50% the length of the promontorium) (0); large (, 100% or more the length of the promontorium) (1). The mastoid region often has an irregular shape and is difficult to characterize. Among the taxa sampled here, all cetaceans (except Inia ), ruminants (except Tragulus ), camelids, perissodactylans, and † mesonychians have large mastoid processes. Suines have small mastoid processes, as do hippopotamids. In the latter group, even though the mastoid region is very swollen, relative to the promontorium its length is not particularly pronounced.
Presence of a large mastoid region (e.g., figs. 21, 77, 110) is a widespread feature among the taxa studied here. A small mastoid region is a synapomorphy of each of the following clades: suines and their fossil stem taxa and hippopotamids. However, each of these clades derived this feature independently.
25. Shape of mastoid region ( Kasuya, 1973; Messenger and McGuire, 1998): square, robust knob (0); wedge (1); convex dorsal surface and flat/irregular ventral surface (seen in the following views:posterior, ventrolateral; ventromedial, dorsal, internal). The mastoid region can be difficult to characterize, but three general groups of shape differences emerge in the sample of taxa examined here. Those three groups are: a square, knoblike, posteriorly blunt mastoid regions (e.g., fig. 55); a wedge or fan-shaped mastoid often with an attenuated posterior end (e.g., fig. 26); or a mastoid region with a convex dorsal surface and flat/irregular ventral surface (e.g., fig. 90). Extant cetaceans and some extinct cetaceamorphans, although quite heterogeneous in their mas- toid region shape, generally exhibit a mastoid region with a convex dorsal surface and the flat/irregular ventral surface. The wedgeshaped mastoid region is found in ruminants (except Tragulus ), camelids, perissodactylans, and † mesonychians. Suines exhibit a square, knob-shaped mastoid region. The region is variable in the extinct noncetacean artiodactylans; these taxa exhibit either the square shape or the wedge shape.
Optimizations indicate that the wedgeshaped mastoid region occurs in basal artiodactylans, is primitive for Artiodactyla , and occurs in outgroups such as perissodactylans and † mesonychians. The Suines and their stem taxa are characterized by a knobshaped mastoid region, and ruminants, with the exception of Tragulus , have the wedgeshaped condition. The unstable position of several extinct taxa make optimization of this character at major nodes in Artiodactyla somewhat ambiguous. It does appear that the wedge condition seen in ruminants is not retained from the ancestral artiodactylan but is instead an independent derivation of this shape.
26. Epitympanic wing: absent (0); present (1). This feature is a flange off the promontorium that extends in both an anterior and lateral direction (in ventromedial view) and often contacts the tegmen tympani if it is extensive. The presence of the epitympanic wing is widespread in noncetacean artiodactylans as well as outgroups such as perissodactylans and † Protungulatum . The epitympanic wing is absent in extant cetaceans.
Optimization indicates that the presence of an epitympanic wing is primitive for artiodactylans. It reverses once to absent in stem taxa to cetaceans but it is ambiguous as to which node first marks the reversal due to a lack of data for fossil taxa that are basal cetaceamorphans.
27. Posteromedial flange of promontorium: absent (0); present (1). This is a flat flange of bone that extends from the medial and posterior edge of the promontorium. In many artiodactylans this flange is continuous with both the epitympanic wing and the caudal tympanic process (e.g., fig. 33). This structure is present in most fossil and living noncetacean artiodactylans, absent in † Protungulatum , and variably present in † meso- nychians. It is absent in extant cetaceans and in the fossils † Dorudon and † Basilosaurus , but it is present in † Pakicetus .
Optimization of the posteromedial flange indicates that its presence is a synapomorphy of Artiodactyla + Perissodactyla . A reversal occurs to the absence of the flange within the stem to Cetacea at least as low on the tree as † Dorudon . The exact node where the absence appears is ambiguous.
28. Promontorium shape (Luo and Gingerich, [1999]): hemi-ellipsoid (0); hemispherical (1). This character, which needs to be examined in anterior, ventromedial, and medial views to be visible, is an attempt to describe how the promontorium of many cetaceans differs from that of many noncetacean artiodactylans and other taxa. Cetaceans have a very ovoid promontorium that bulges ventrally as a distinct hemisphere. Other taxa, however, have a less bulging and more ellipsoid shape to the promontorium.
Luo and Gingerich (1999) also identified similarities in petrosal shape in † mesonychids, † basilosaurids, and other extinct cetaceamorphans to the exclusion of † Pakicetus . My observations indicate, however, that † Pakicetus , for which I have only studied the petrosal in situ (fig. 96), more closely resembles † mesonychians and other noncetacean ungulates in this feature, and that † basilosaurids resemble more highly nested extant cetaceans.
The hemispherical condition is shared by the clade that includes † basilosaurids + extant cetaceans (fig. 1), appearing somewhere between † Pakicetus and † Dorudon (data are ambiguous). The primitive condition for the taxa examined here is to have a hemi-ellipsoid promontorium (e.g., figs. 7–8, 16–17).
In studies of the petrosal anatomy of such taxa as mammaliamorphs, variations in the promontorium shape are often considered to reflect the amount of expansion and torsion of the underlying cochlea (e.g., Wible et al., 1995: 3). When comparing certain cetaceans to noncetacean artiodactylans and other taxa, however, there is considerable variation in the morphology of the promontorium, even though all of these have a coiled cochlea. A separate study comparing size and shape of the underlying cochlea to the external structure of the promontorium could test whether there are any regular relationships to this variation.
29. Caudal tympanic process: mediolaterally narrow bar of bone medial to the stapedial muscle fossa (0); mediolaterally broad bar of bone medial to the stapedial muscle fossa, sometimes with irregular projections or knobs (1). I am employing a broad definition of this structure, somewhat in contrast to Geisler and Sanders (2003). Most of the taxa examined here have a mediolaterally broad type of caudal tympanic process, and this structure may be continuous with the crista interfenestralis or the posteromedial flange off the promontorium. In † Protungulatum and some cetaceans the caudal tympanic process is narrow. In Suines the caudal tympanic process often develops into a knob medially.
Optimizations indicate that the primitive condition for Artiodactyla + Perissodactyla is to have a broad caudal tympanic process; however, this does change to a narrow process at the base of Cetacea , where this state is a synapomorphy.
30. Mastoid plate: absent (0); present (1). This structure on the endocranial surface of the mastoid region is a flat plate of bone that extends distally relative to the ventral surface of the petrosal. In endocranial view this structure, if present, obscures a view of the rest of the mastoid region. When differentially expanded this way, the endocranial side is plate-shaped and distinctly different from the ventral side. Of the sample of taxa examined here, this occurs in extant suines only (fig. 55) and is an ambiguous synapomorphy at the node marking their common ancestor because the condition is not known in the extinct stem taxon † Perchoerus .
31. Basicapsular groove (Wible et al., 1995; Luo and Gingerich, 1999; Geisler and Luo, 1998): present (0); absent (1). The shape of the medial edge of the promontorium varies in the presence, conspicuousness, and position of a groove that runs anteroposteriorly along the medial margin. Luo and Gingerich (1999) and others have reported that this groove is for the inferior petrosal sinus, which according to MacIntyre (1972: 291) connects the cavernous sinus with the internal jugular vein. This groove carries the internal carotid artery when the artery occupies a medial position relative to the promontorium (e.g., Wible, 1986; Wible et al., 2004). In its more elongate forms, this sulcus can extend from the epitympanic wing to the anterior aspect of the mastoid region. In some artiodactylans the edge of the basicapsular groove has a sharp, ruffled border. More dissections of the extant taxa studied here will be important for documenting the exact soft tissue contents of this structure in artiodactylans and relatives.
Most noncetacean artiodactylans have this groove. In some taxa, such as Tragulus napu (figs. 3, 7), however, this groove is absent and the medial edge of the petrosal is sharp with no dorsally or ventrally positioned ridges. This corroborates an observation by Luo and Gingerich (1999) that in taxa where the groove is absent, the promontorium may terminate in a sharp edge as it transitions from the dorsomedial to ventrolateral surfaces. In most living and extinct cetaceans described here, with the exception of Ziphius (fig. 90), the medial petrosal is smooth and bulbous with no conspicuous groove.
The presence of the groove optimizes as primitive for all taxa studied here, a result consistent with Geisler and Luo’s (1998: 196) reconstruction of the primitive morphotype of the common ancestor of artiodactylans. The groove is present in most taxa examined here with the exception of Tragulus napu and † Poebrotherium sp. (figs. 36, 38); this optimizes in each case as an independent loss of the groove. This character transforms to absent in the common ancestor of extant and extinct cetaceans (fig. 1, but this is an ambiguous synapomorphy).
32. Position of basicapsular groove: ventral (0); dorsal (1). Most of the taxa examined here that have a basicapsular groove have it on the dorsal surface of the petrosal. However in some taxa, the groove is in a more ventral position and is visible on the tympanic surface of the bone. Taxa that have the ventral position are Moschus , hippopotamids, Babyroussa, and † Bothriogenys . Each of these taxa derived this independently. The primitive condition for artiodactylans is to have a dorsally positioned groove.
33. Secondary facial foramen position relative to fenestra cochleae and fenestra vestibuli: anterior or lateral to any part of these fenestrae (0); posterior to both fenestrae (1). In two suine species, Sus scrofa (fig. 52) and Hylochoerus meinertzhageni (fig. 57), the secondary facial foramen opens much farther posteriorly than in other species (e.g., figs. 30, 60).
The primitive condition for artiodactylans is to have an anteriorly positioned secondary facial foramen. The posterior position is independently derived in the two suids mentioned above.
34. Cochlear aqueduct position (contains opening of perilymphatic foramen, the end of cochlear canaliculus in human anatomy text books, or cochlear canaliculus; Ekdale et al., 2004; see also Cifelli, 1982): dorso-internal (0); ventral (1). This character distinguishes the very large and dorsally positioned cochlear aqueduct seen in cetaceans from the much smaller, more ventrally positioned structure seen in noncetacean artiodactylans and other mammals examined here. In some cases, particularly with fossils, the preservation may be such that it is hard to identify the exact position of a small cochlear aqueduct; nonetheless, it can be clearly seen in these taxa that they do not exhibit the cetacean condition. As stated by MacIntyre (1972: 290), the cochlear aqueduct often opens into the jugular notch. There is a distinct difference in the state of this character between extant cetaceans and other ungulates. In extant cetaceans the cochlear aqueduct opens dorsally (e.g., fig. 95), close to the internal acoustic meatus, and is also quite large, rivaling the size of foramina inside the internal acoustic meatus. In most taxa examined here, however, this foramen opens ventromedially in the vicinity of the jugular foramen and is very small.
The primitive condition for artiodactylans is to have a ventral or ventromedial position for the cochlear aqueduct. The dorsally positioned foramen is an ambiguous synapomorphy of the common ancestor of † Dorudon atrox and Delphinidae (fig. 1).
CHARACTERS DESCRIBING THE PETROSAL- BULLA ARTICULATION
35. Articulation of anterior crus/tube of ectotympanic with external acoustic meatus portion of petrosal ( Geisler and Luo, 1998; Luo and Gingerich, 1999): absent (0); present
(1). This character describes a contact between the anterior part of the ectotympanic (whether shaped as a ring or a tube) and the petrosal. The contact typically occurs in the region of the petrosal contribution to the external acoustic meatus or the ventrolateral tuberosity (if present). This contact is posterior to the anterior process of the tegmen tympani. When present, this contact is typically relatively small compared to other contacts between the tympanic and the petrosal.
Luo and Gingerich (1999: 58) stated that ‘‘Direct articulation of the anterior crus of the ectotympanic with the tegmen tympani of the petrosal is rare in eutherians’’; however, I have observed contact between the petrosal and the anterior process in many of the ungulates examined here, including both perissodactylans (fig. 113) and artiodactylans (e.g., fig. 19).
The primitive condition for artiodactylans is to have contact between these structures. However, the absent condition appears in a number of places within Artiodactyla , such as at the stem and base of Cetacea , in Ruminantia and on its stem, and in Camelidae .
36. Articulation of posterior crus/tube of ectotympanic and external acoustic meatus part of petrosal ( Luo and Gingerich, 1999): absent (0); present (1). Luo and Gingerich (1999) described the ‘‘ventral exposure of the mastoid region’’ and its relative cover by the posterior process of the tympanic bulla. A contact of some kind is present in almost all taxa examined here.
The primitive condition for artiodactylans (and even artiodactylans and perissodactylans) is to have contact present. This does show homoplasy; however, particularly among camelids + † Poebrotherium , lack of contact is a synapomorphy of this clade.
37. Relative size of articulation of posterior crus/tube of ectotympanic and external acoustic meatus part of petrosal ( Luo and Gingerich, 1999: char. 15): large, entire posterior crus covers ventral exposure of mastoid (0); small, half (or less) of posterior crus articulates with mastoid region, region is ventrally exposed (1). There is significant shape variation among the taxa that have large contacts. For example, Ovis aries (fig. 19) has a thin, elongate but substantial contact whereas other pecorans such as Antilocapra americana (fig. 4) have an elongate but broad contact. This character varies widely across taxa examined here.
A small area of contact is primitive for the taxa studied here. There are transformations to a large contact within artiodactylans but many nodes are ambiguous. An exception is Pecora (fig. 1), a clade for which a large contact is a synapomorphy, and highly nested odontocetes (the common ancestor of Delphinidae and Pontoporia ), which also exhibit large contacts. A large contact is present in hippopotamids but it is ambiguous whether this is a shared derived feature with cetaceans or a convergent appearance.
38. Texture of articulation of posterior crus/ tube of ectotympanic and external acoustic meatus part of petrosal ( Luo and Gingerich, 1999): smooth (0); rough/sutured (1); fused (2). The articulation between the tympanic and the mastoid region varies in texture from smooth to rough. I have also designated a separate state, ‘‘fused’’, for which the texture cannot be assessed. Luo and Marsh (1996: 331) discussed the variation of this structure in odontocetes. They noted that this area is typically a rough surface with ‘‘rugose ridges’’, but in Kogia breviceps (figs. 84–85) and Ziphius cavirostris (fig. 85), this surface is smooth. Primitively for artiodactylans the texture of this area is smooth. The character exhibits extensive homoplasy, and for Pecorans and Cetaceans (including the stem) there is a transformation to the rough condition. Cases of fusion are scattered throughout the tree ( Hippopotamus , Bos , Inia , Balaenoptera ) and these are each independent derivations.
39. Articulation between the anterior tympanic bulla and the petrosal: absent (0); present (1). This contact occurs on the anterior and lateral aspect of the petrosal. In pecorans (e.g., figs. 18–19, 21, 23) the contact is often quite large and meets a swelling on the bulla positioned anterior to the bullar part of the external acoustic meatus. Taxa for which this feature is absent are: † Protoceras celer (fig. 31), † Cainotherium , † Archaeotherium (fig. 73), † Diacodexis pakistanensis , † Agriochoerus , Orycteropus afer , and Camelus dromedarius (fig. 31). All of these taxa, except † Archaeotherium and A. americana , have retained the primitive condition.
Primitively, this contact is present for the common ancestor of artiodactylans and perissodactylans and its presence is widespread throughout Artiodactyla . However, because a number of taxa exhibiting the absent condition are camelids or stem camelids, it is not clear what the basal condition is for Artiodactyla .
40. Fusion of the anterior tympanic bulla with the petrosal: absent (0); present (1). In many taxa in which there is a contact between the bulla and the petrosal, the two bones do not fuse. In all extant cetaceans examined, Hippopotamus amphibius (fig. 43), and Equus caballus (fig. 113), however, the two bones fuse.
Fusion is an ambiguous synapomorphy of crown cetaceans. The primitive condition for artiodactylans is to have an unfused contact between these structures.
41. Medial bulla articulation with pars cochlearis: absent (0); present (1). In many of the taxa examined here, there is contact with the bulla along the full anteromedial border of the petrosal, occasionally interrupted by a foramen. This contact typically involves flanges off the promontorium such as the epitympanic wing or the posteromedial flange. In several cetaceans and their stem taxa, however, this contact is absent.
Presence of a medial bullar articulation is widespread among the taxa studied here and is present in the common ancestor of artiodactylans and in the common ancestor of artiodactylans and perissodactylans. Transformation to absence of contact occurs on the stem to Cetacea at † Dorudon , and possibly even more basal to this (it is ambiguous due to unscored fossil taxa).
42. Posterior bulla articulation with pars cochlearis/pars canalicularis ( Muizon, 1987; Messenger, 1994; Messenger and McGuire. 1999: char. 1494): absent (0); present (1). This contact is most often between the bulla and the posteromedial flange or the caudal tympanic process of the petrosal. It is present in most taxa examined here for which the bulla and petrosal articulation is preserved. It is absent, however, in † Dorudon atrox and † Basilosaurus and in all extant cetaceans except Pontoporia blainvillei . Off the cetacean lineage, this is absent in Orycteropus afer , Camelus , and † Protoceras celer . Contact is present in perissodactylans (fig. 113), but because it is absent in some camelids, the condition of contact is ambiguous for the base of artiodactylans.
Presence of a posterior bullar articulation is widespread among the taxa studied here and is present in the common ancestor of artiodactylans and the common ancestor of artiodactylans and perissodactylans. Transformation to absence of contact occurs on the stem to Cetacea at † Dorudon and possibly even more basal to this (it is ambiguous due to unscored fossil taxa).
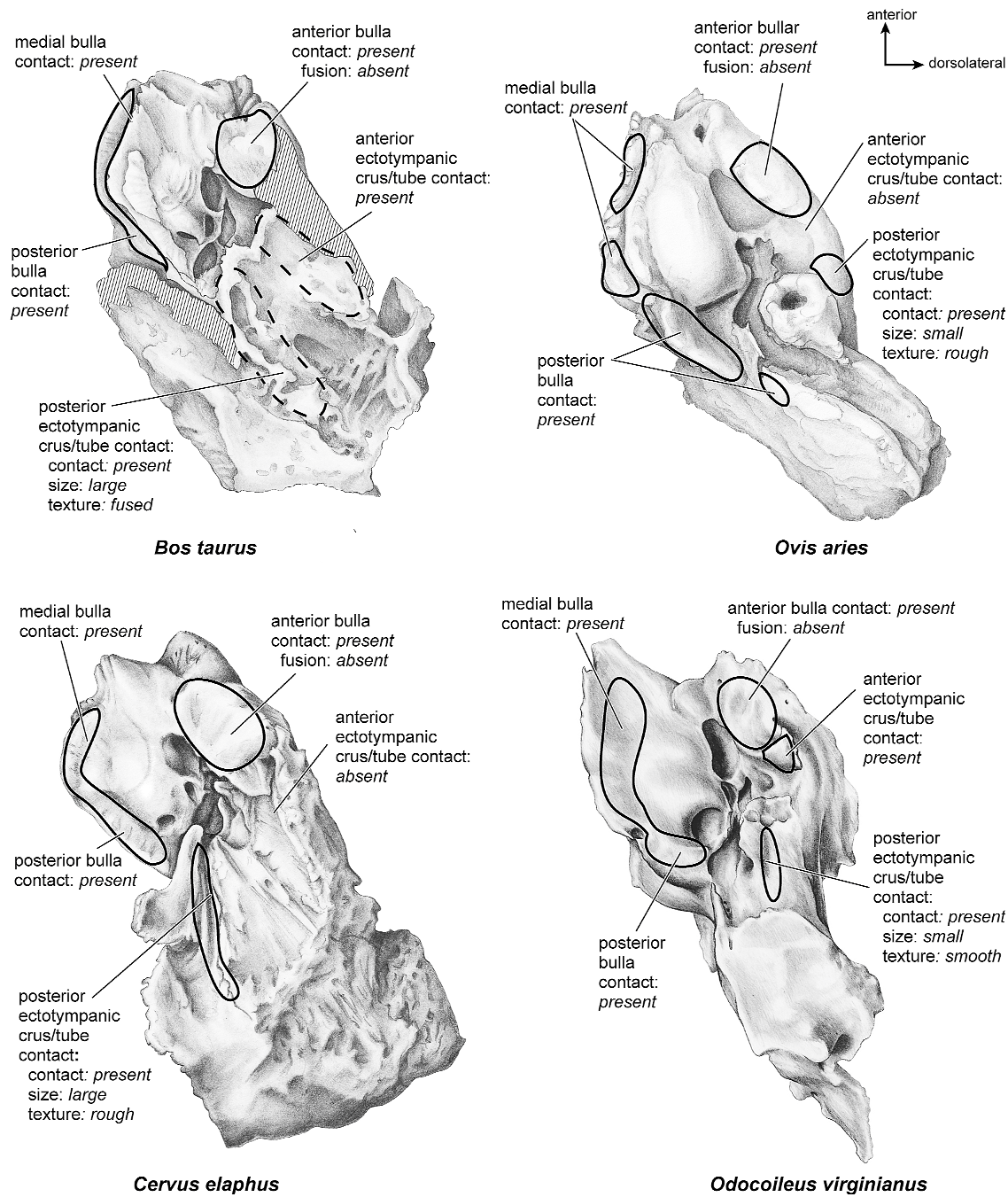
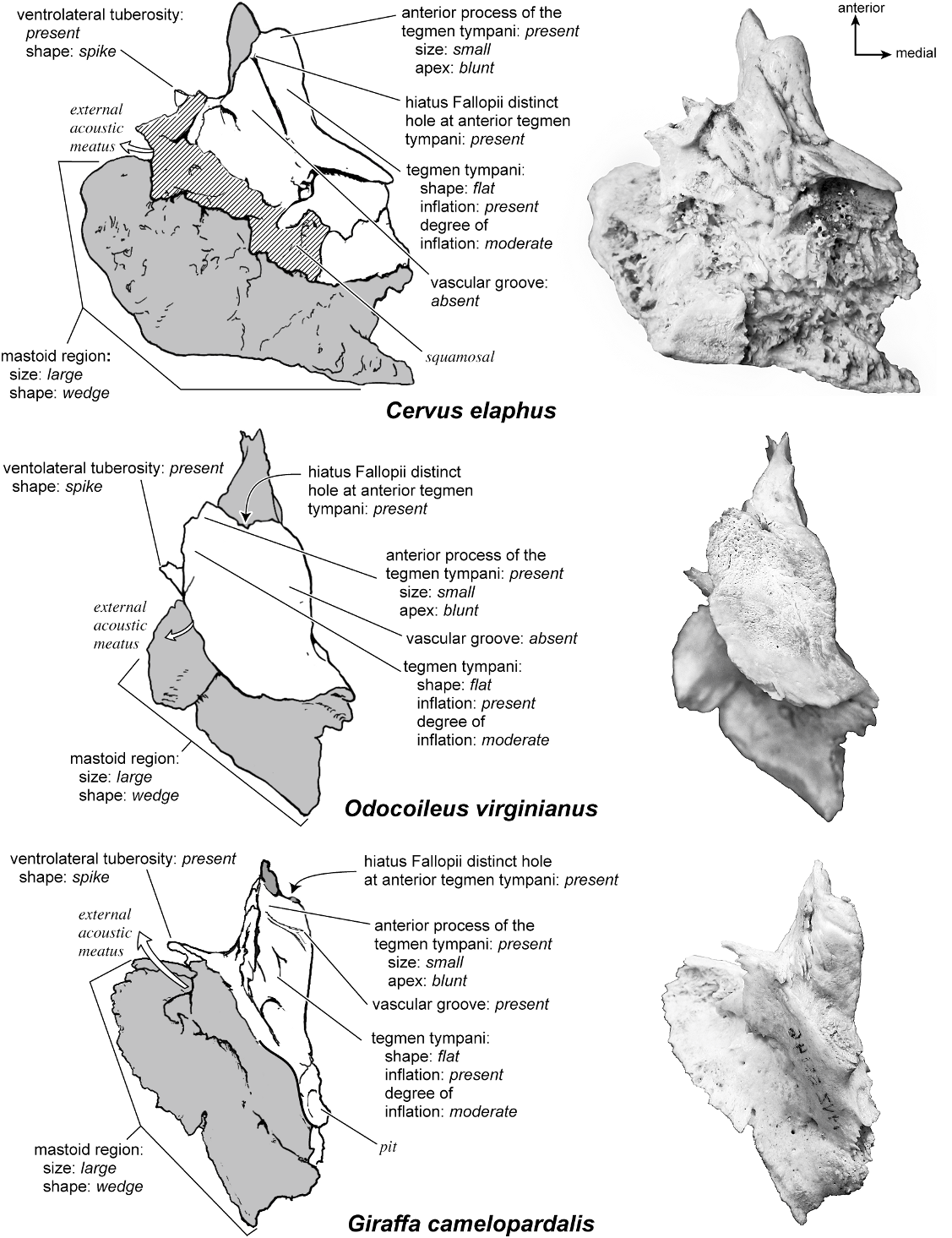

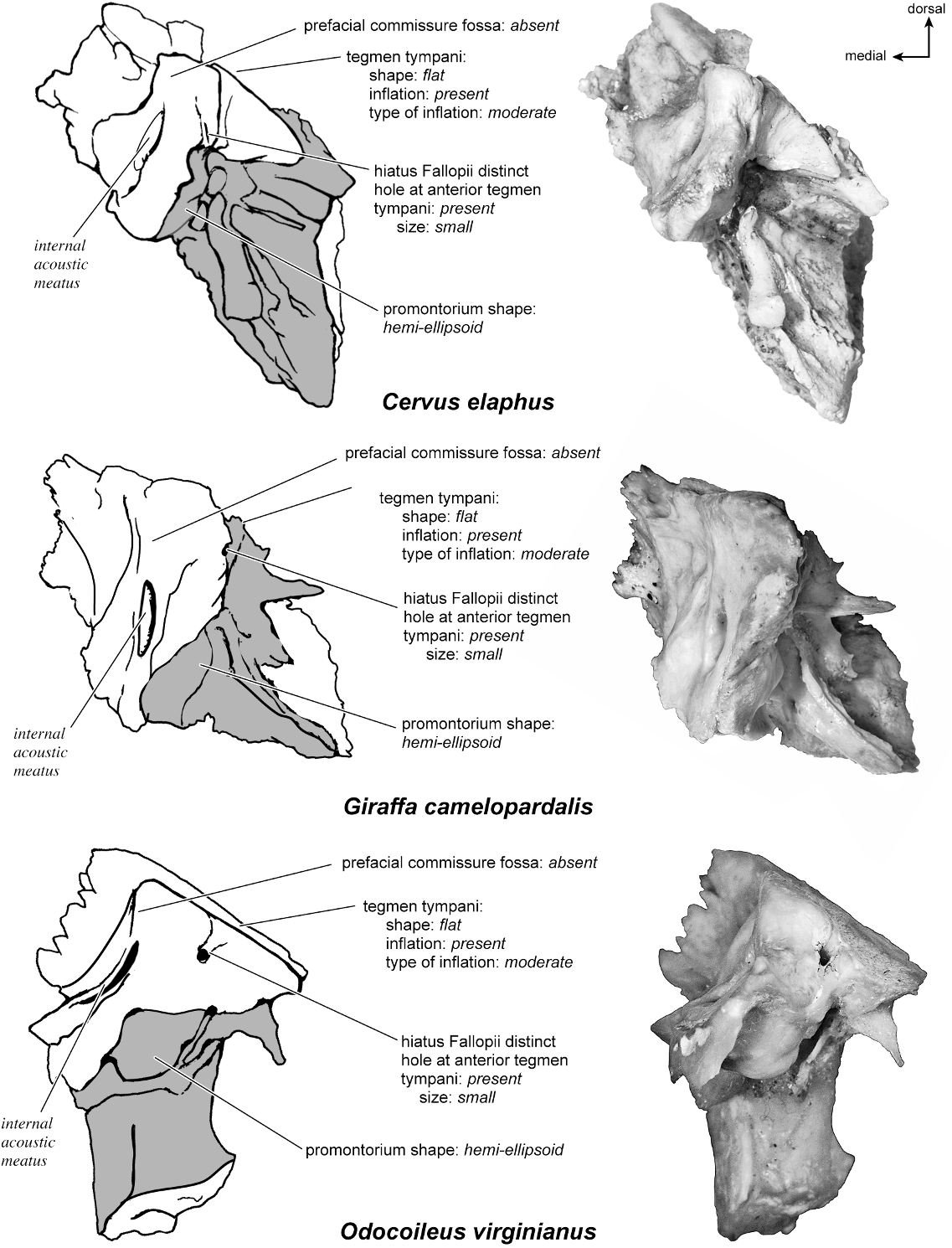

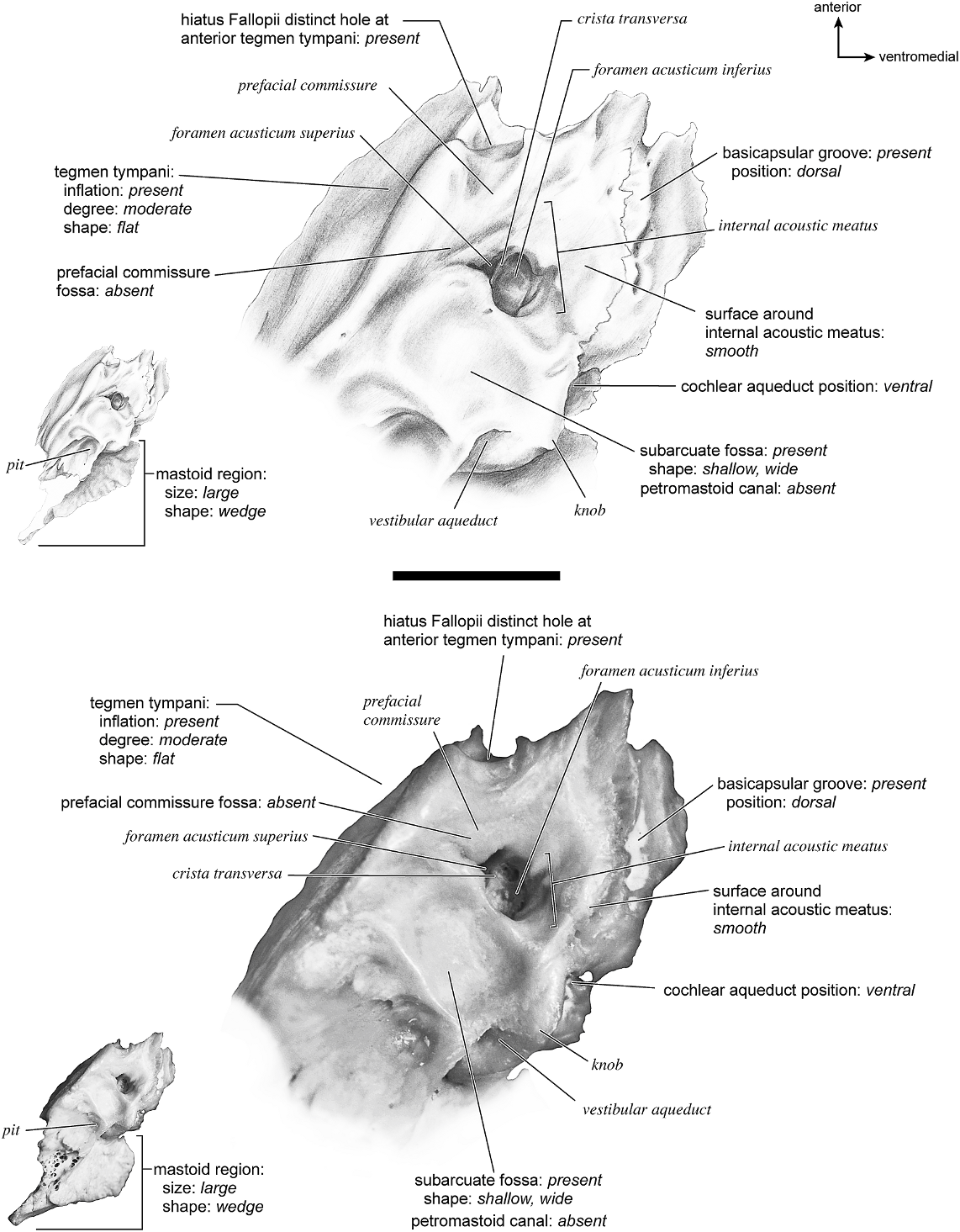

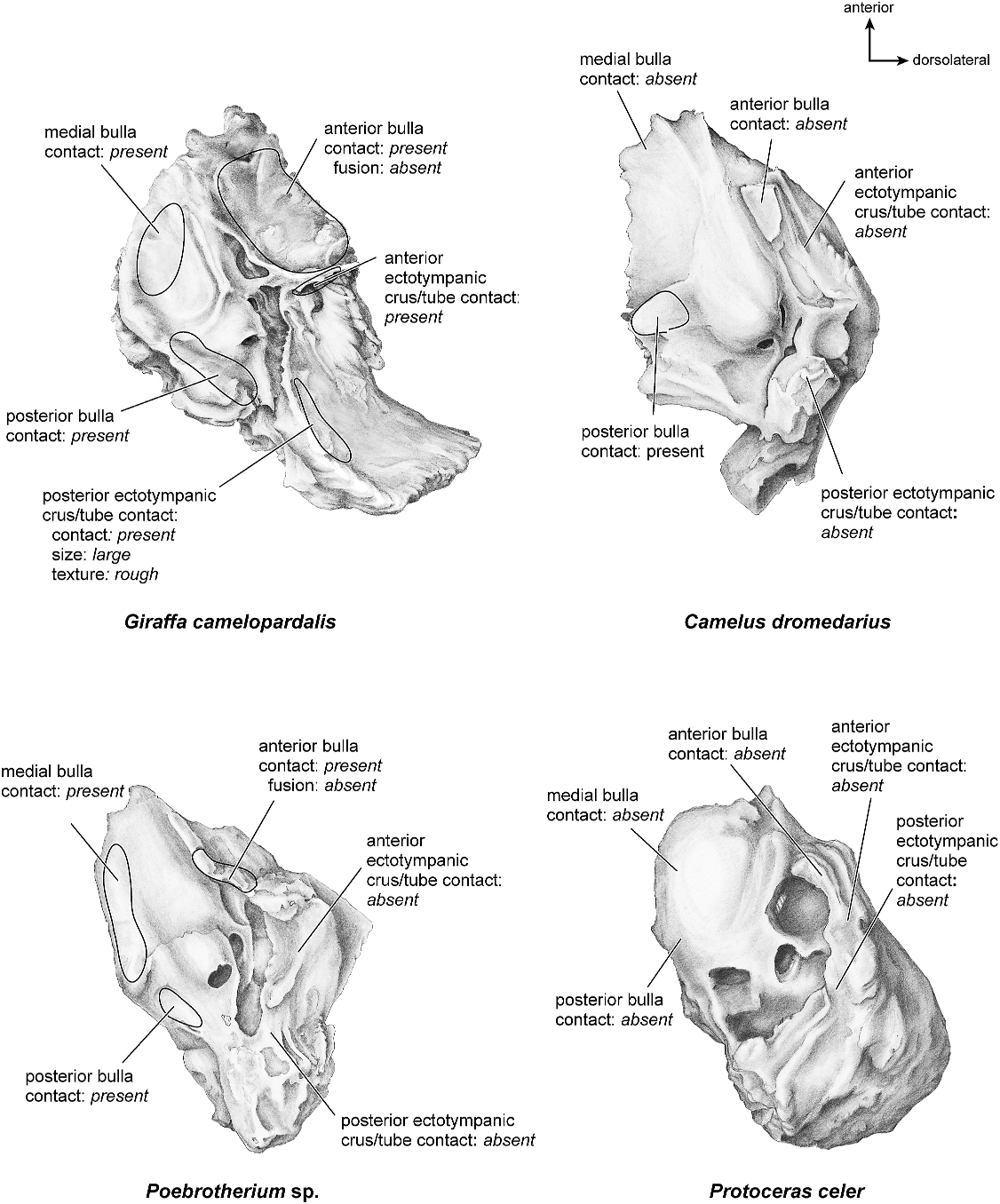
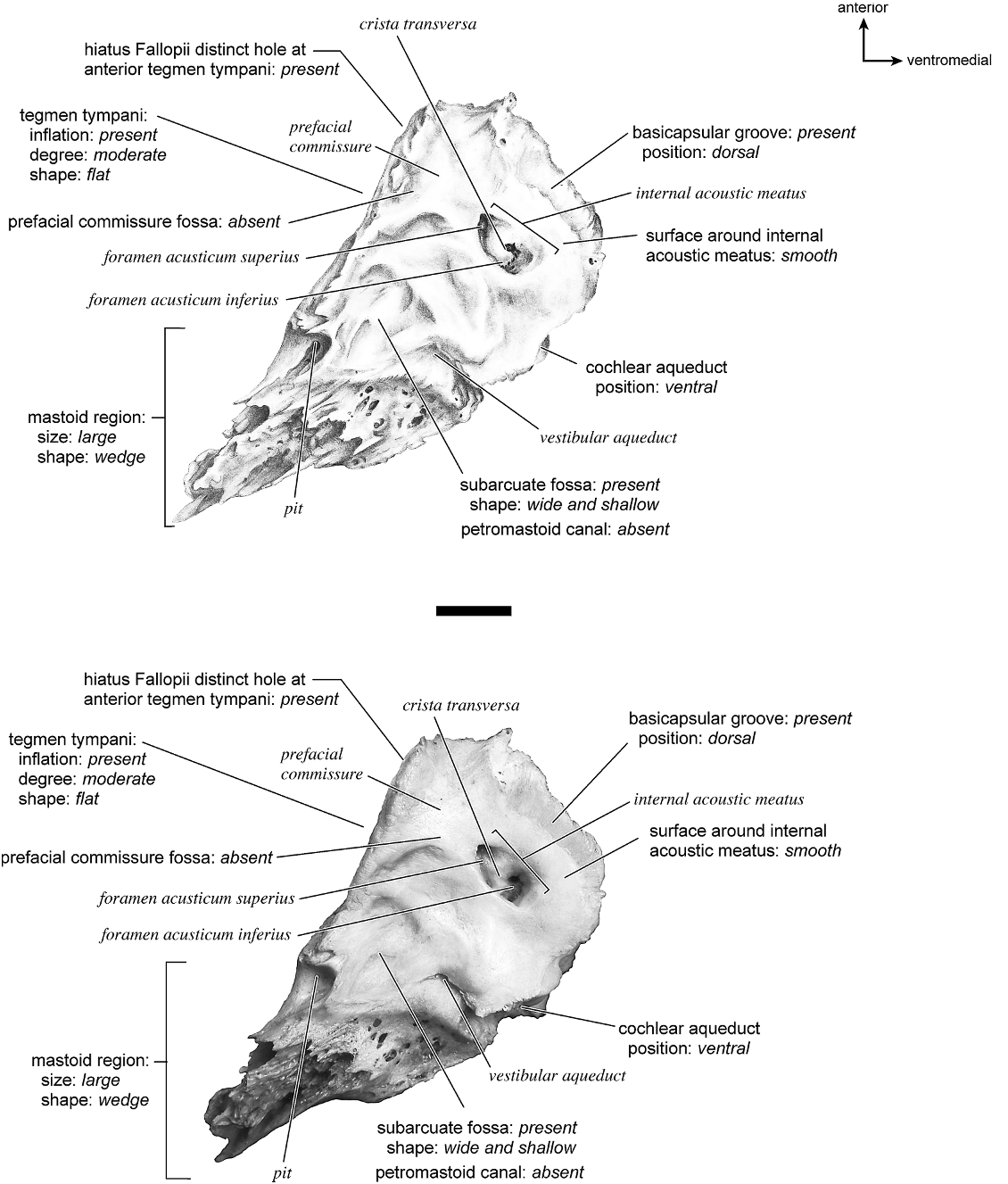

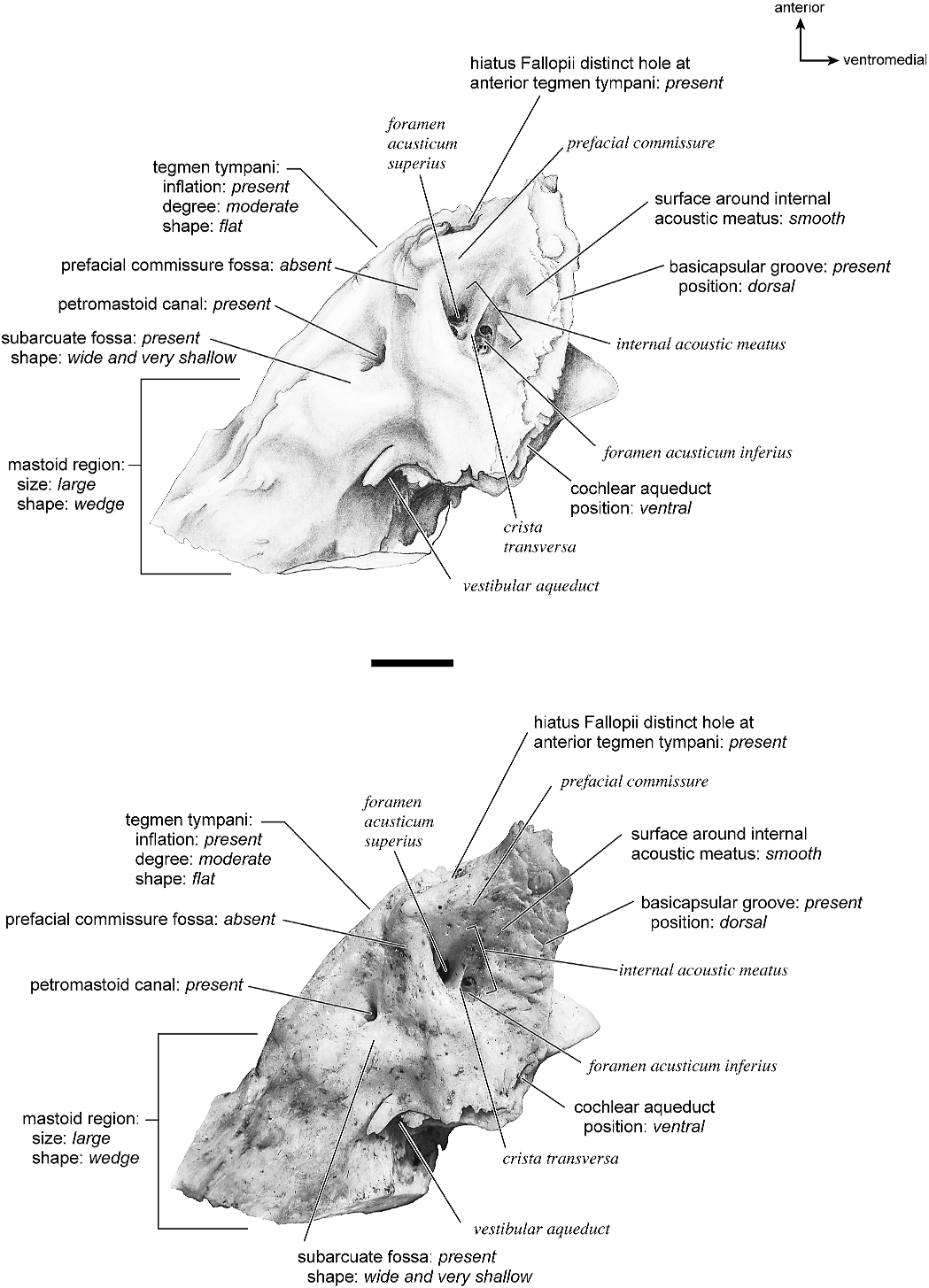
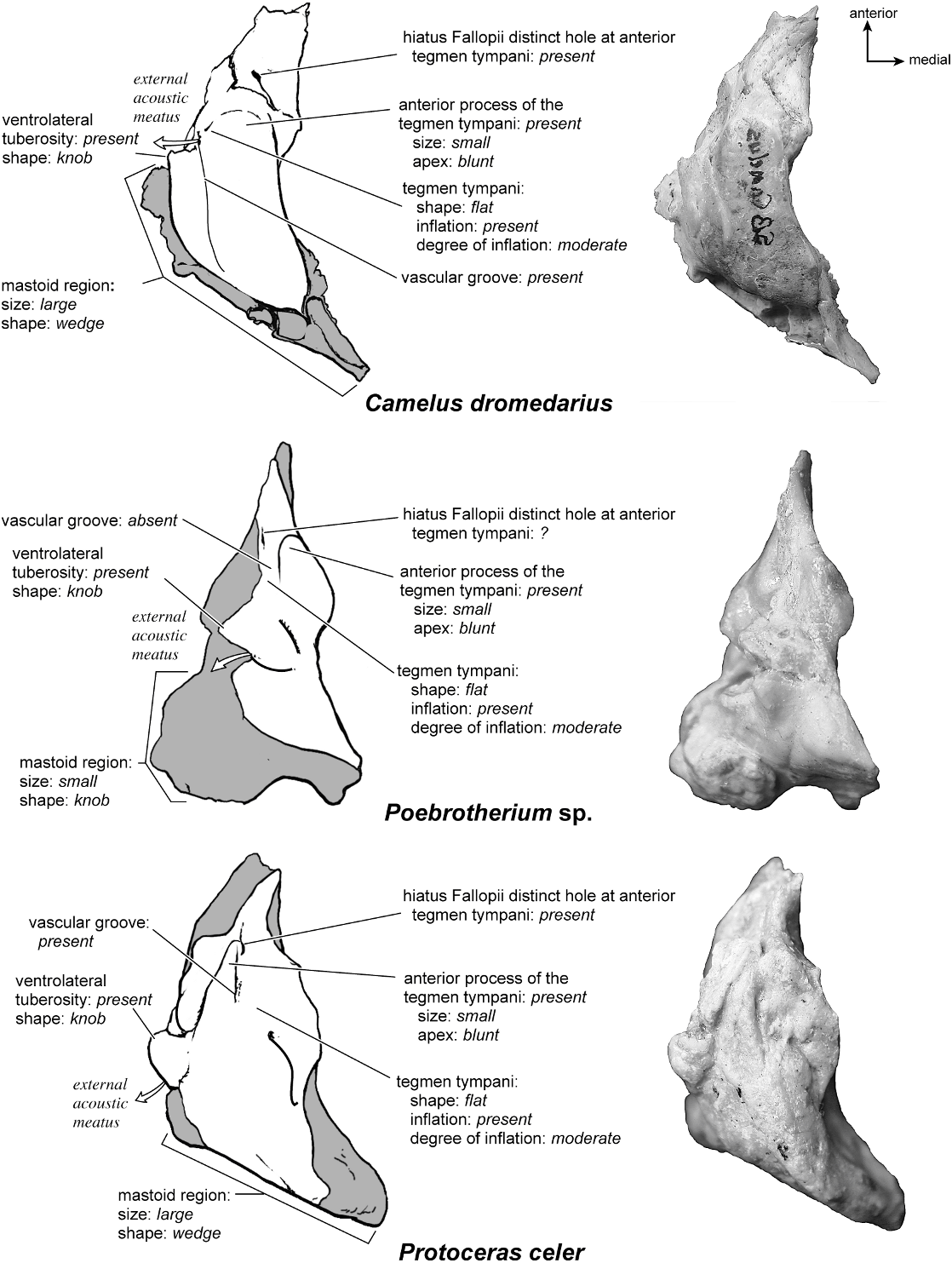

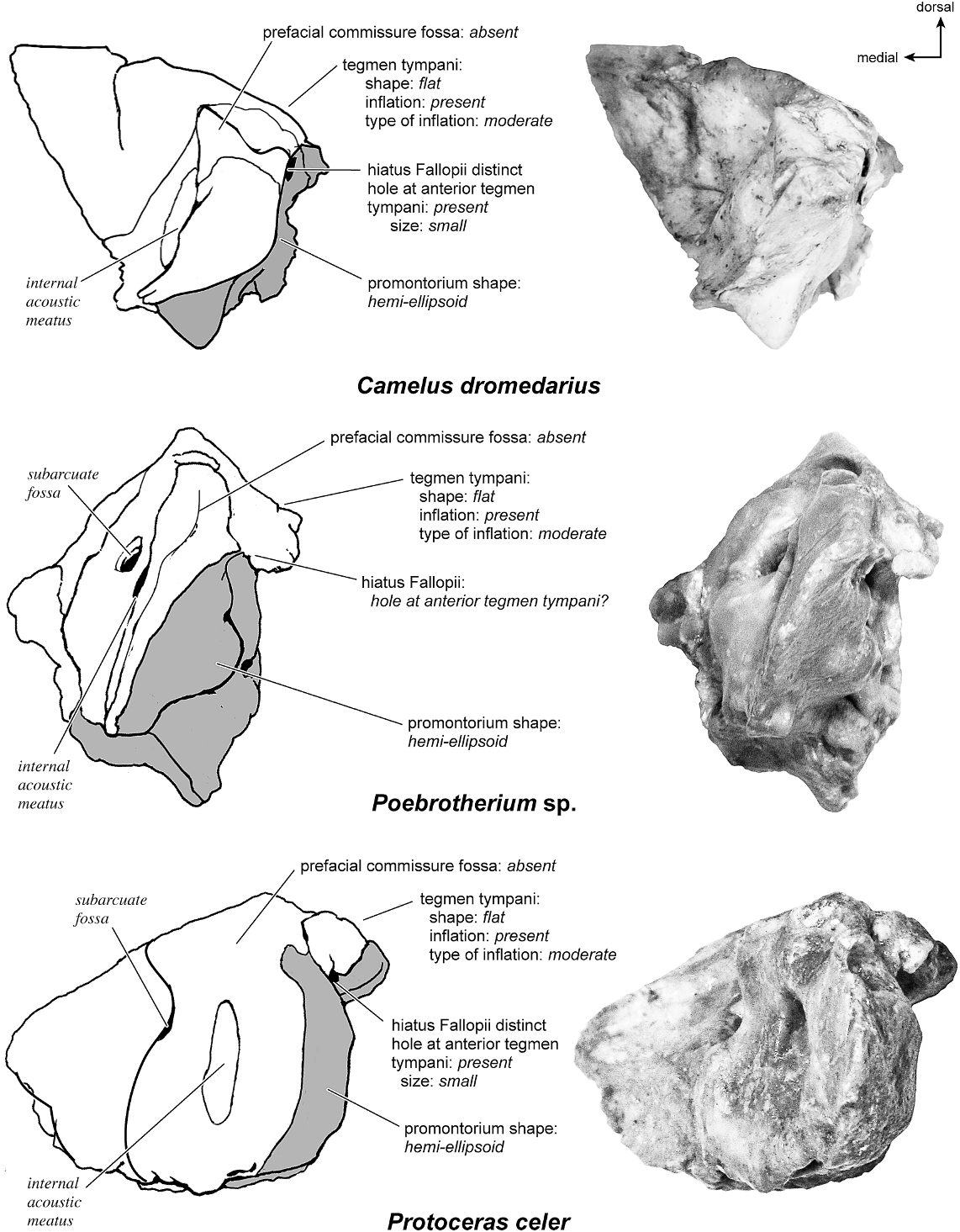
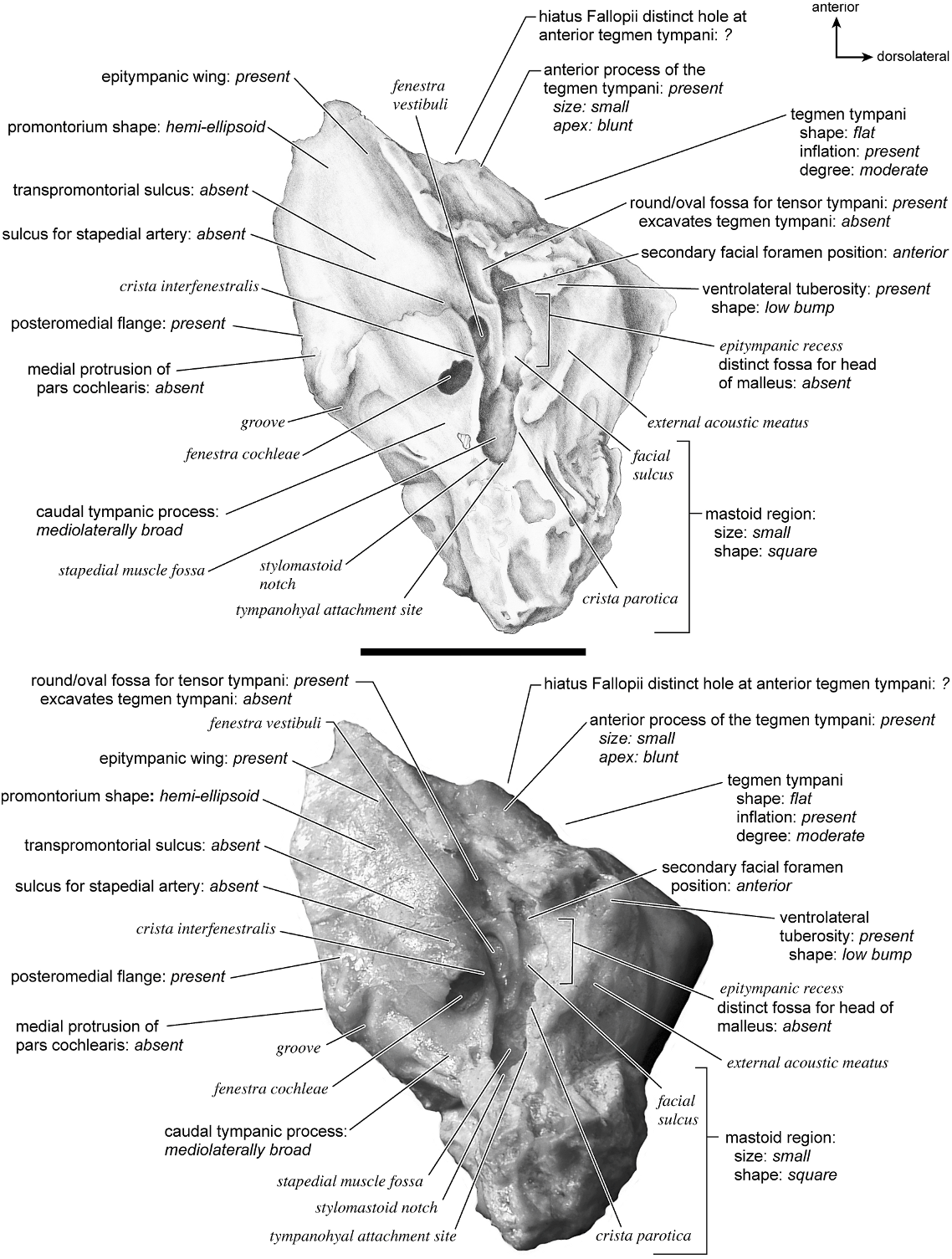
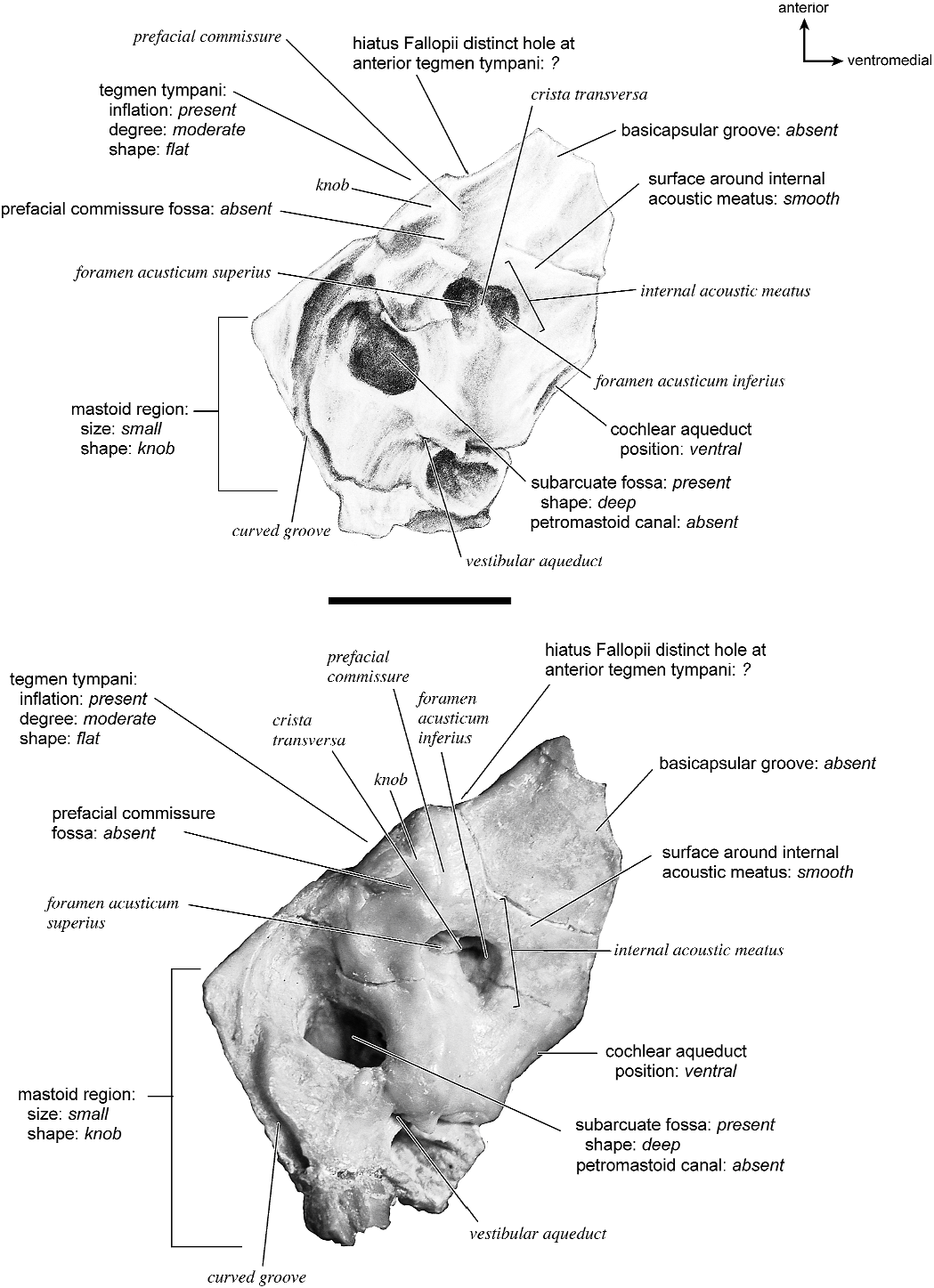
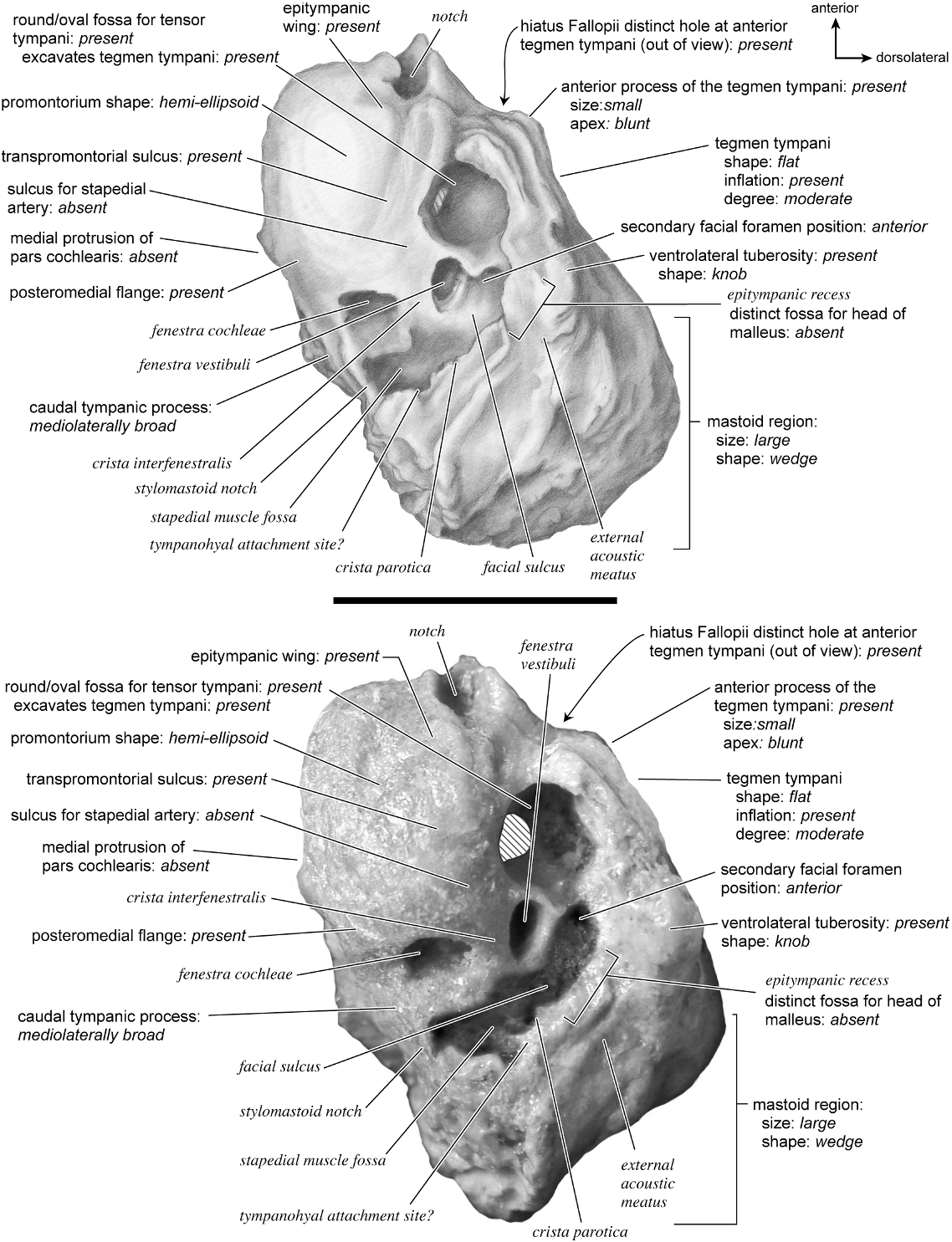


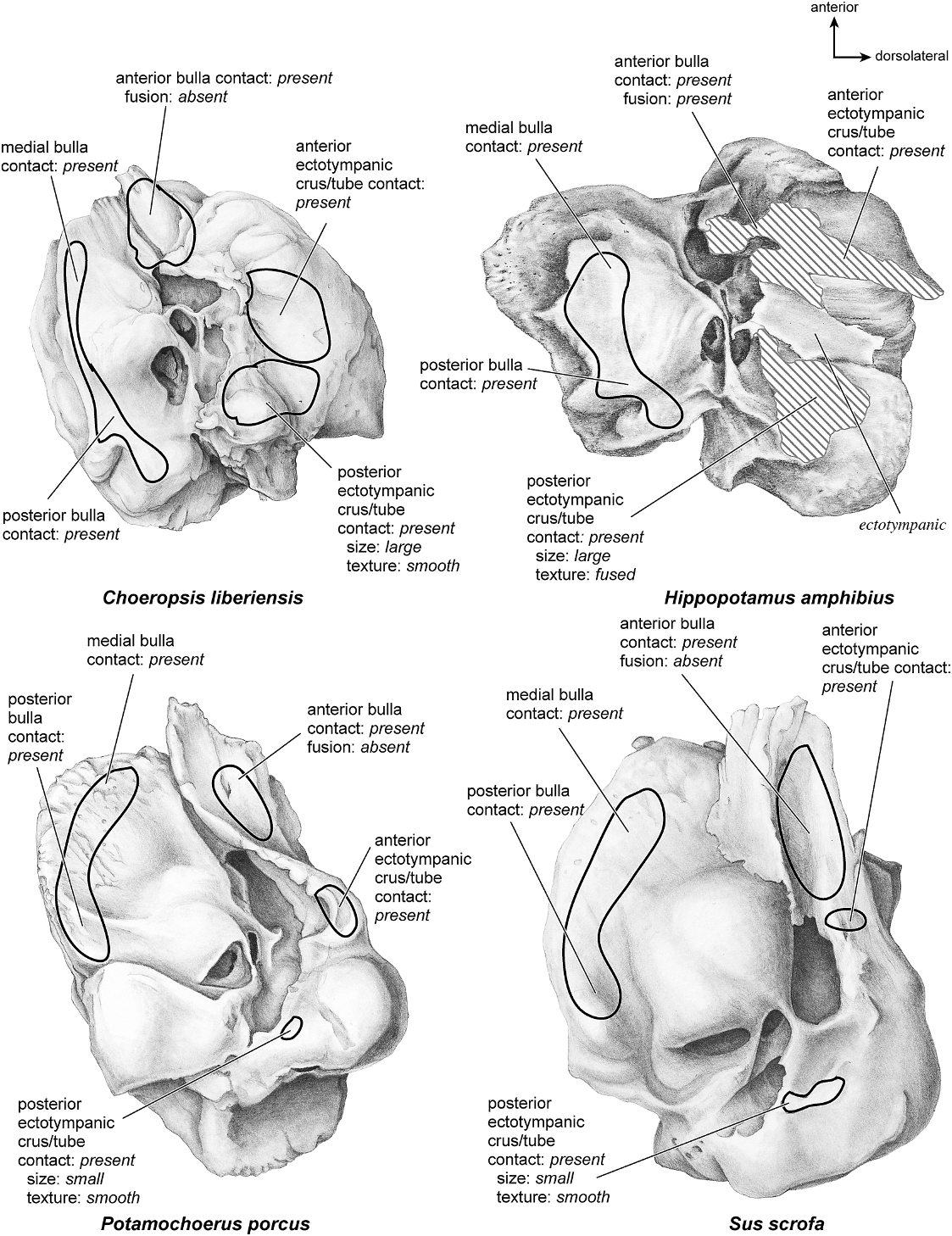


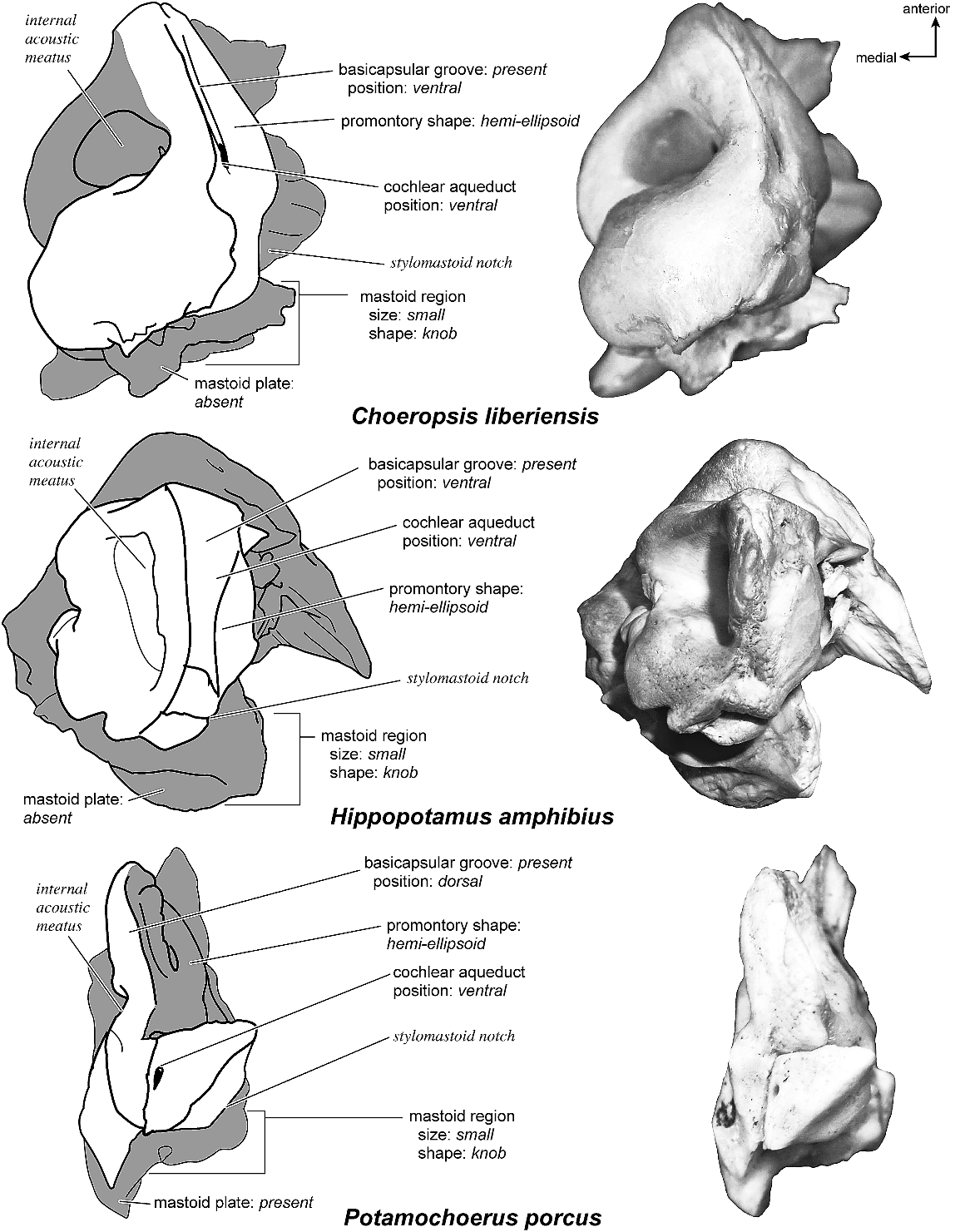

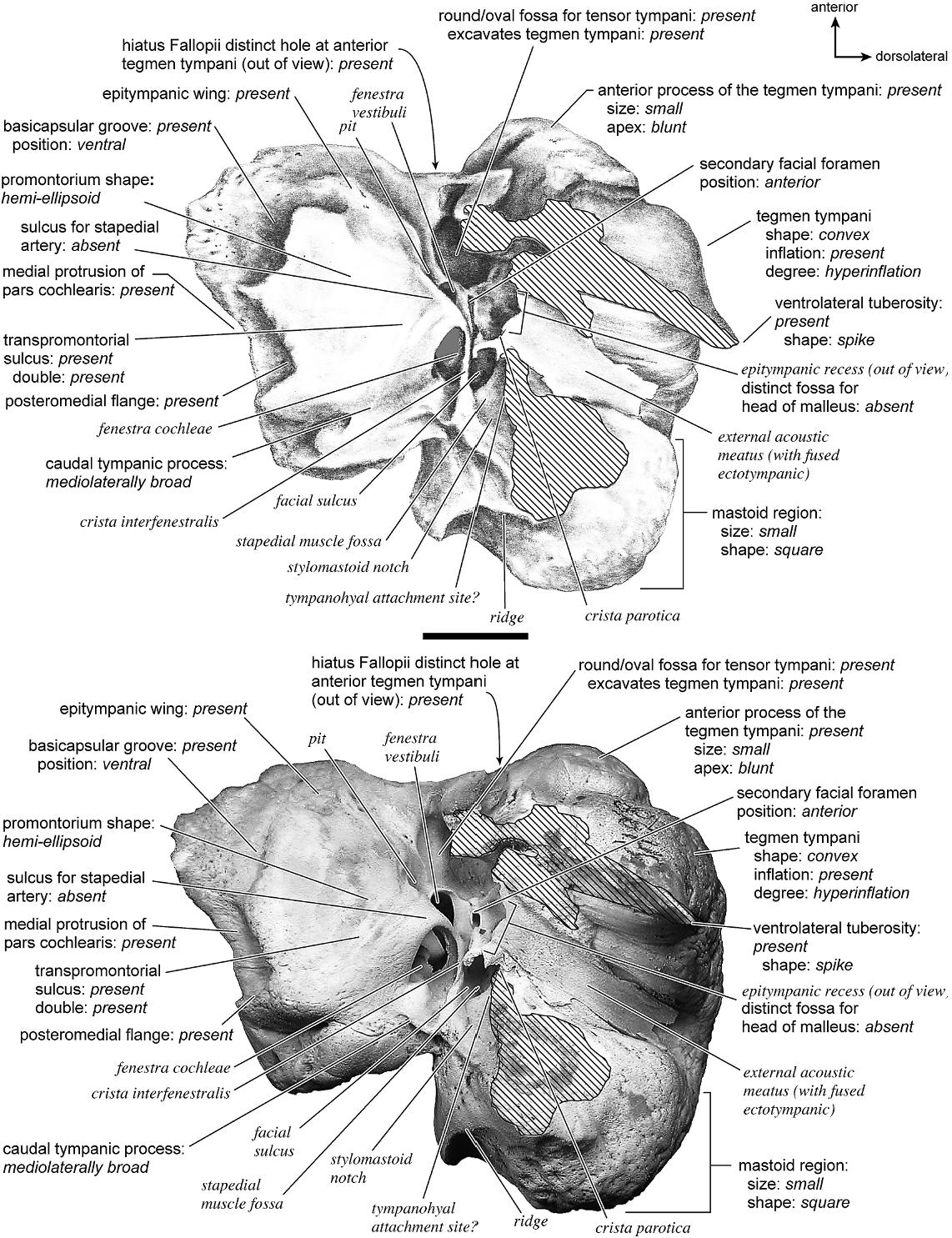

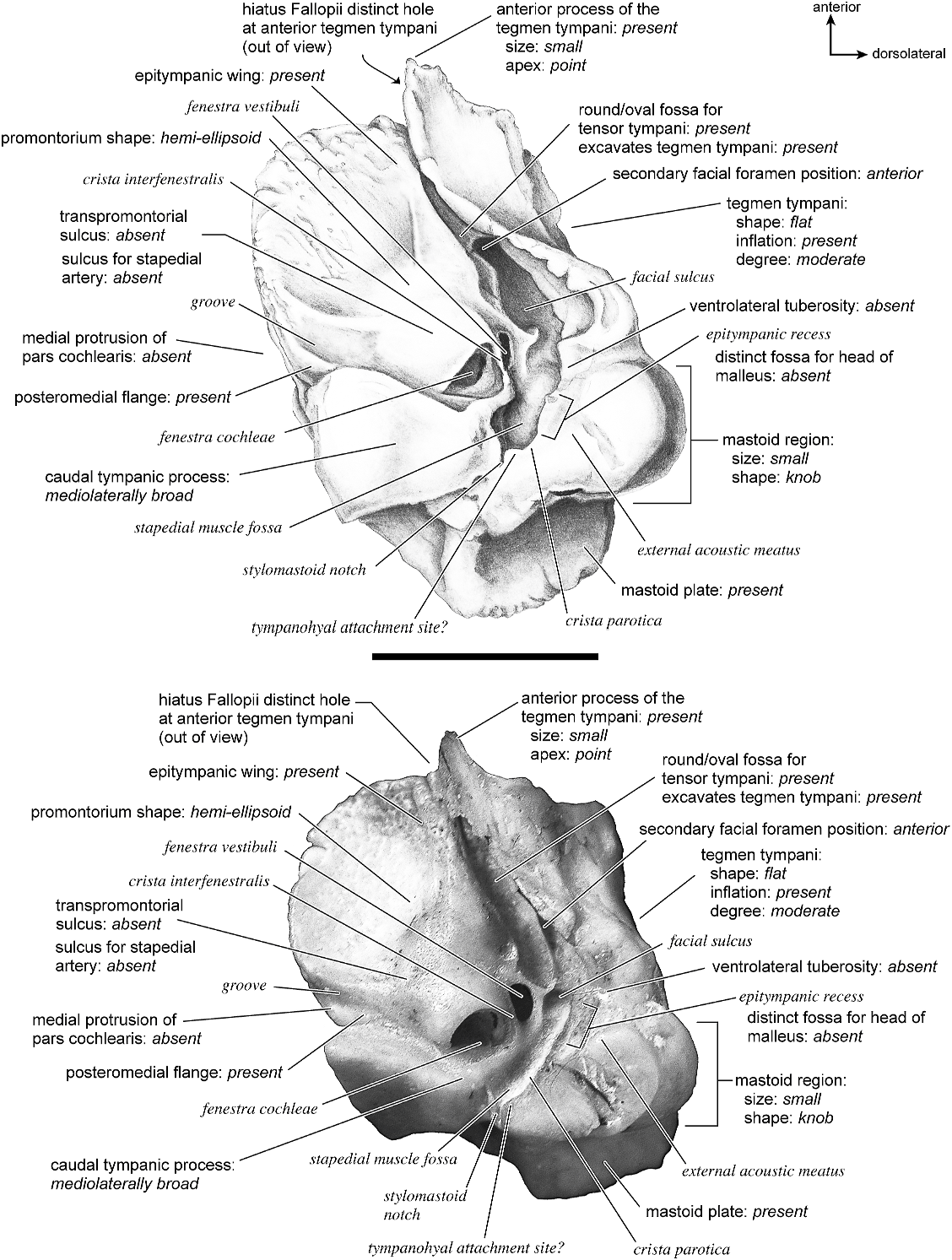
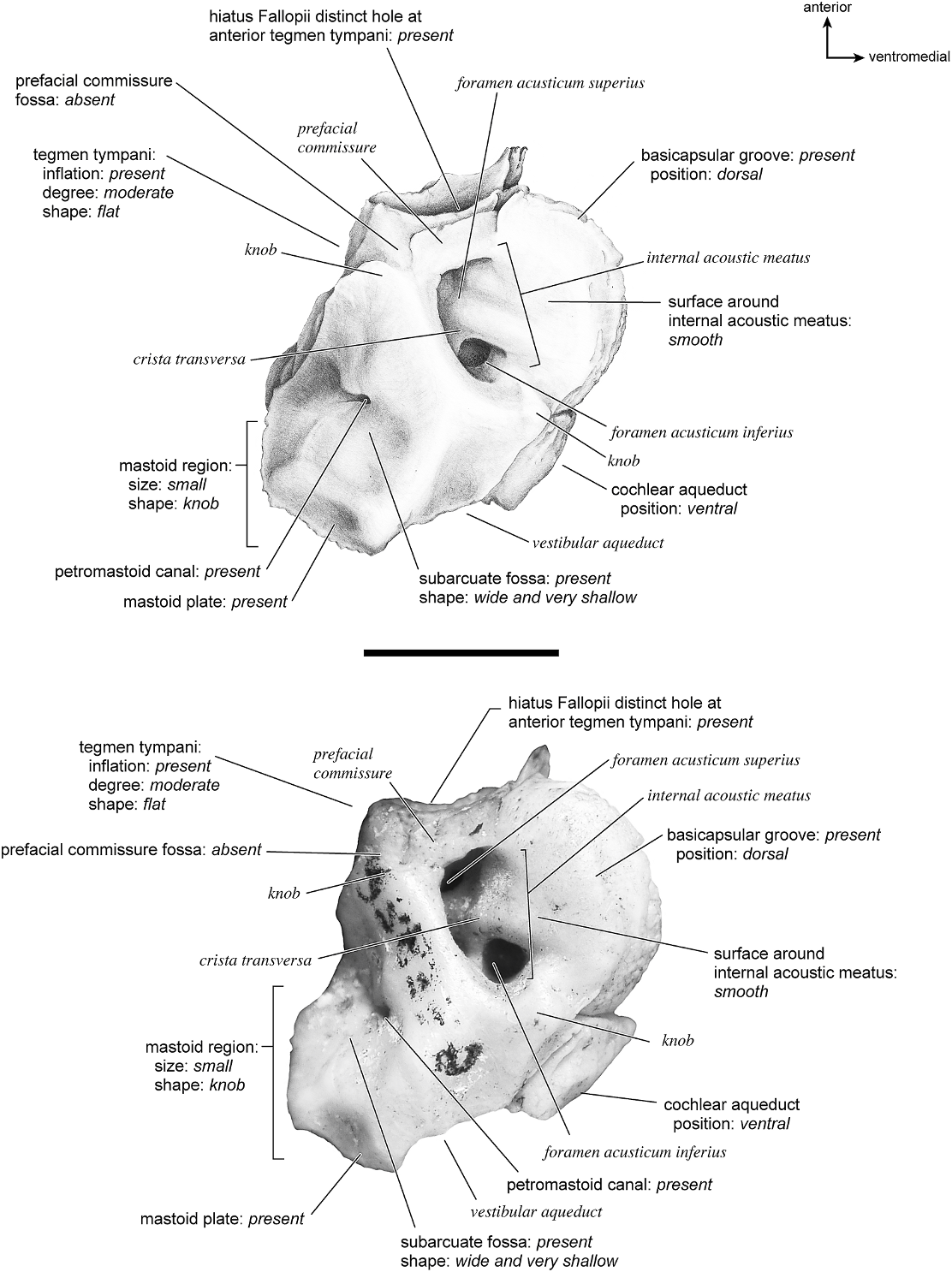


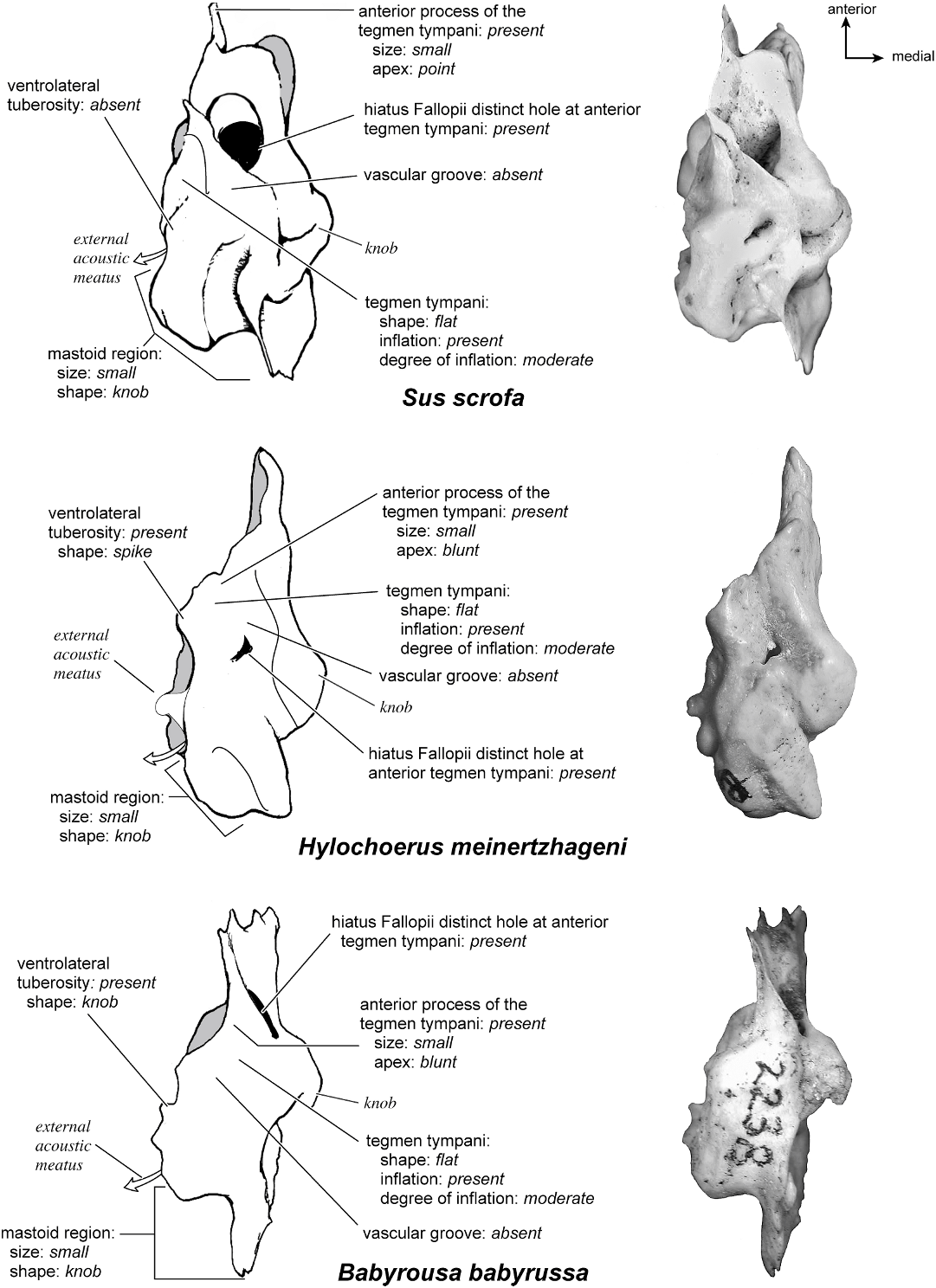
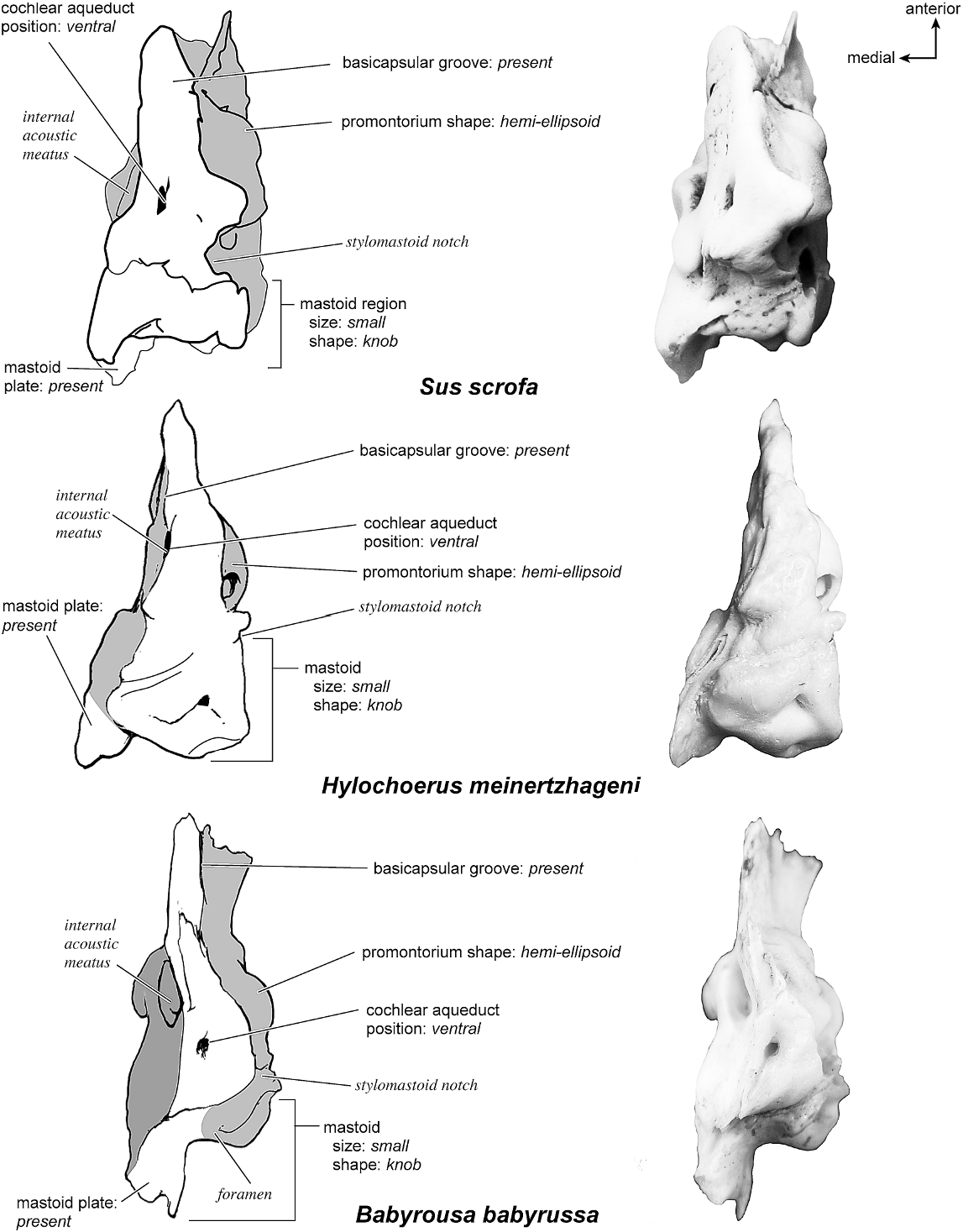



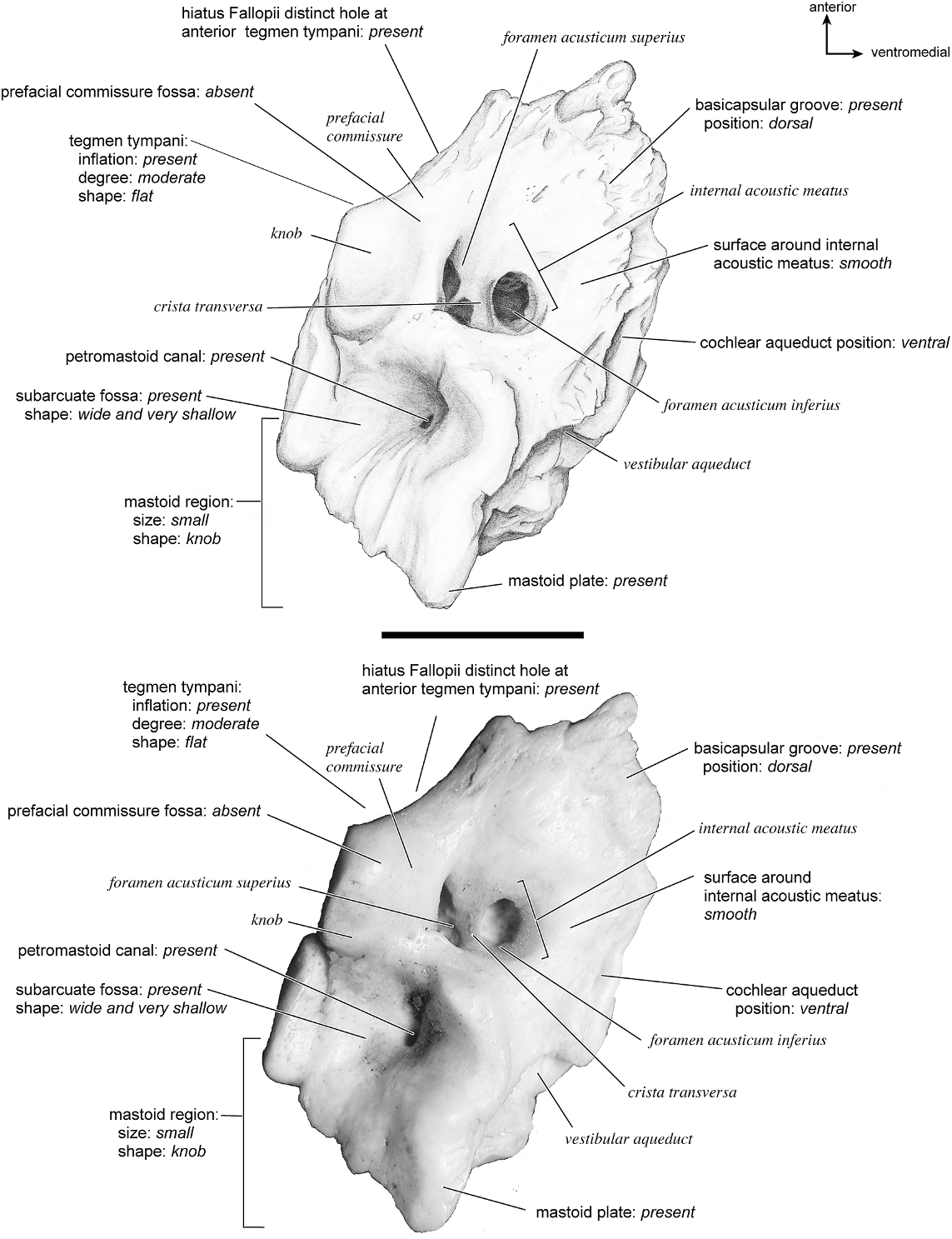
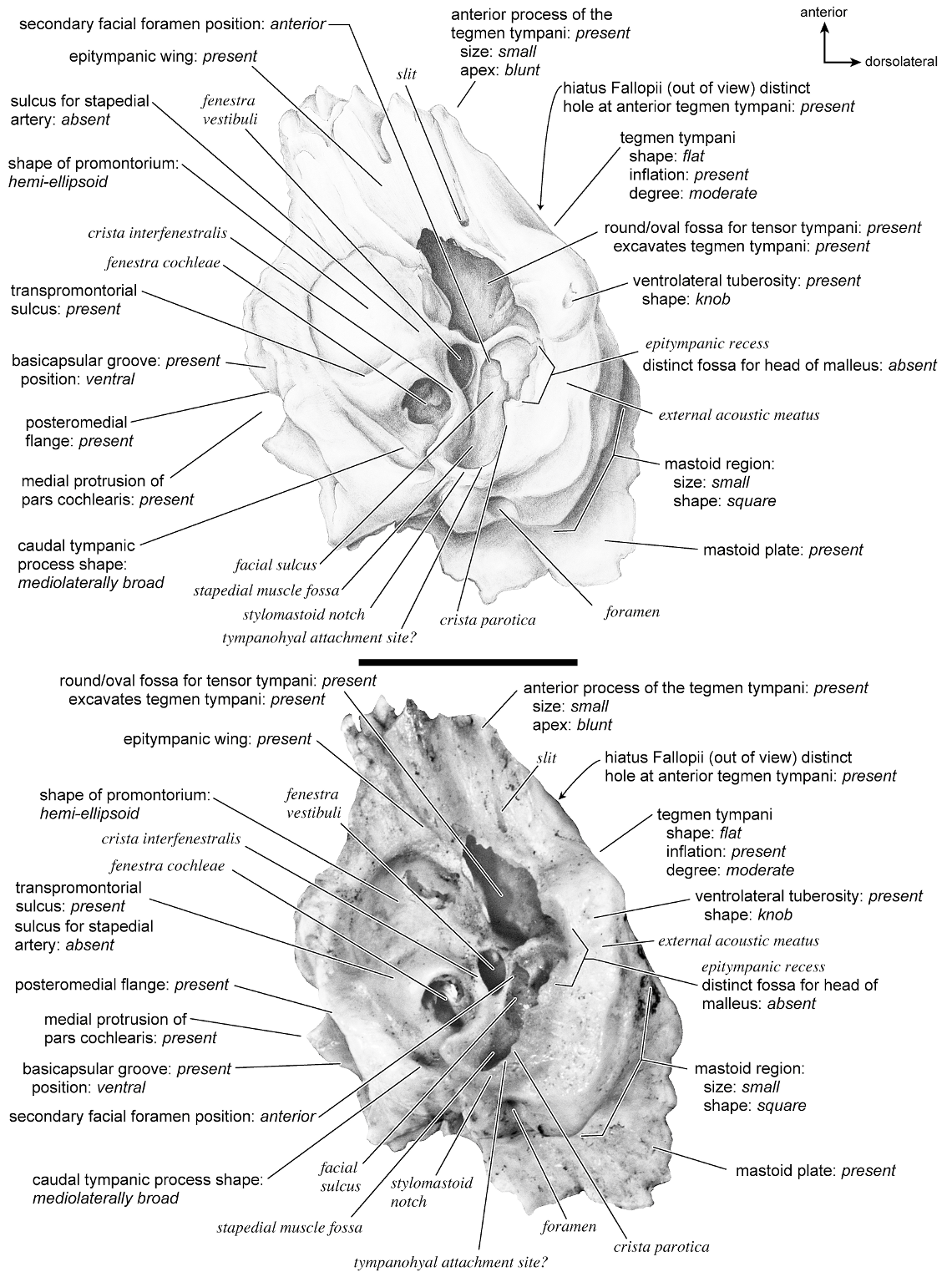
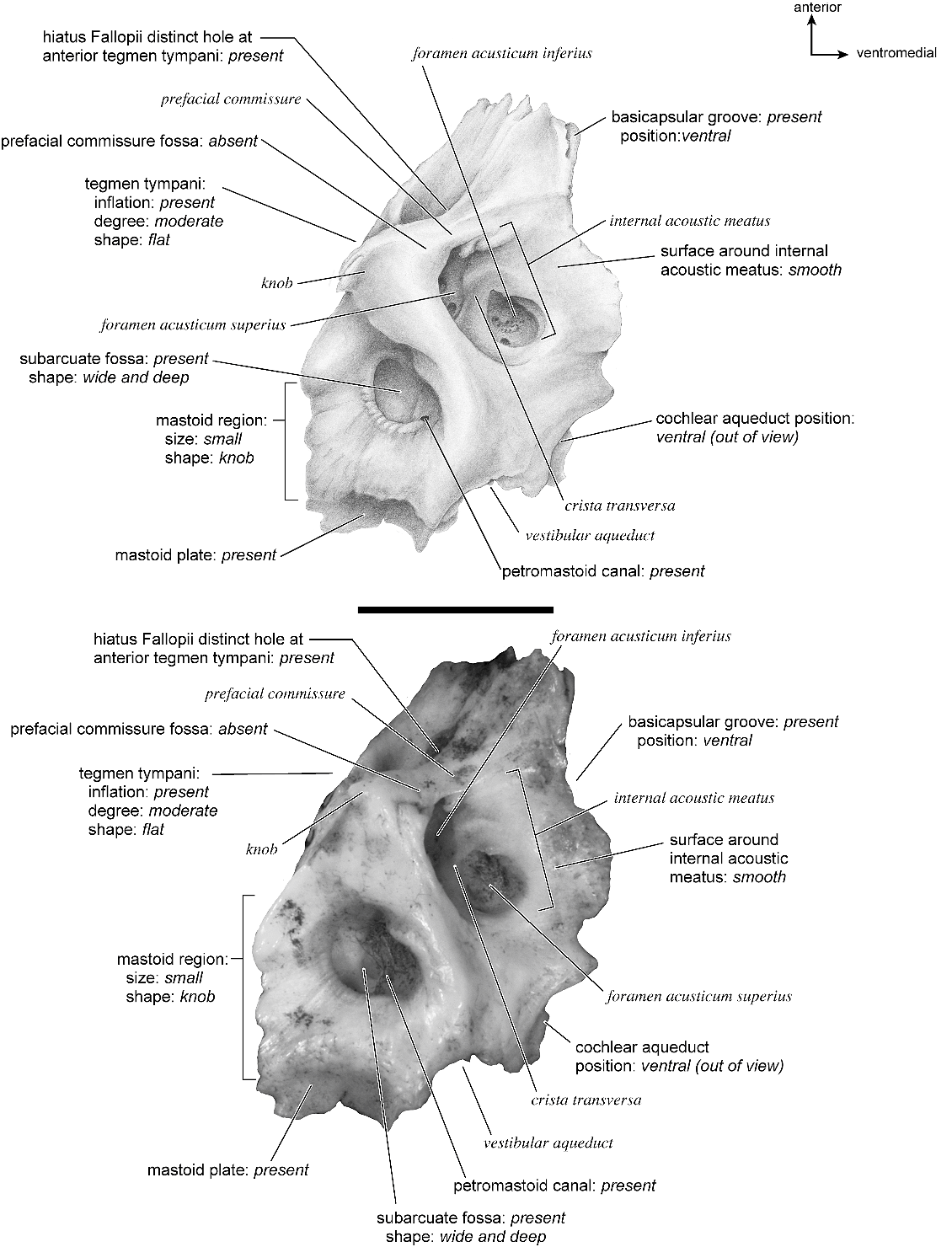
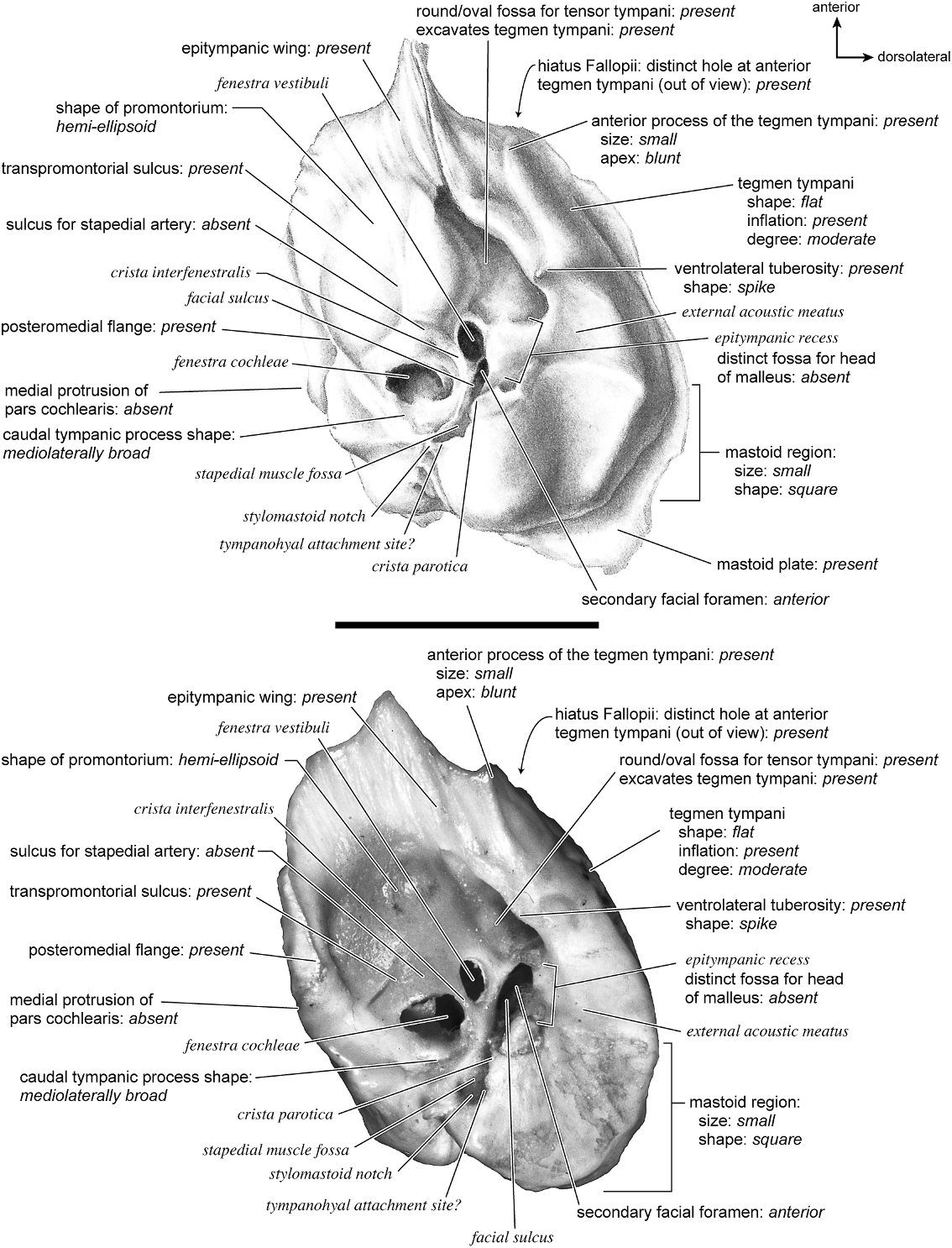
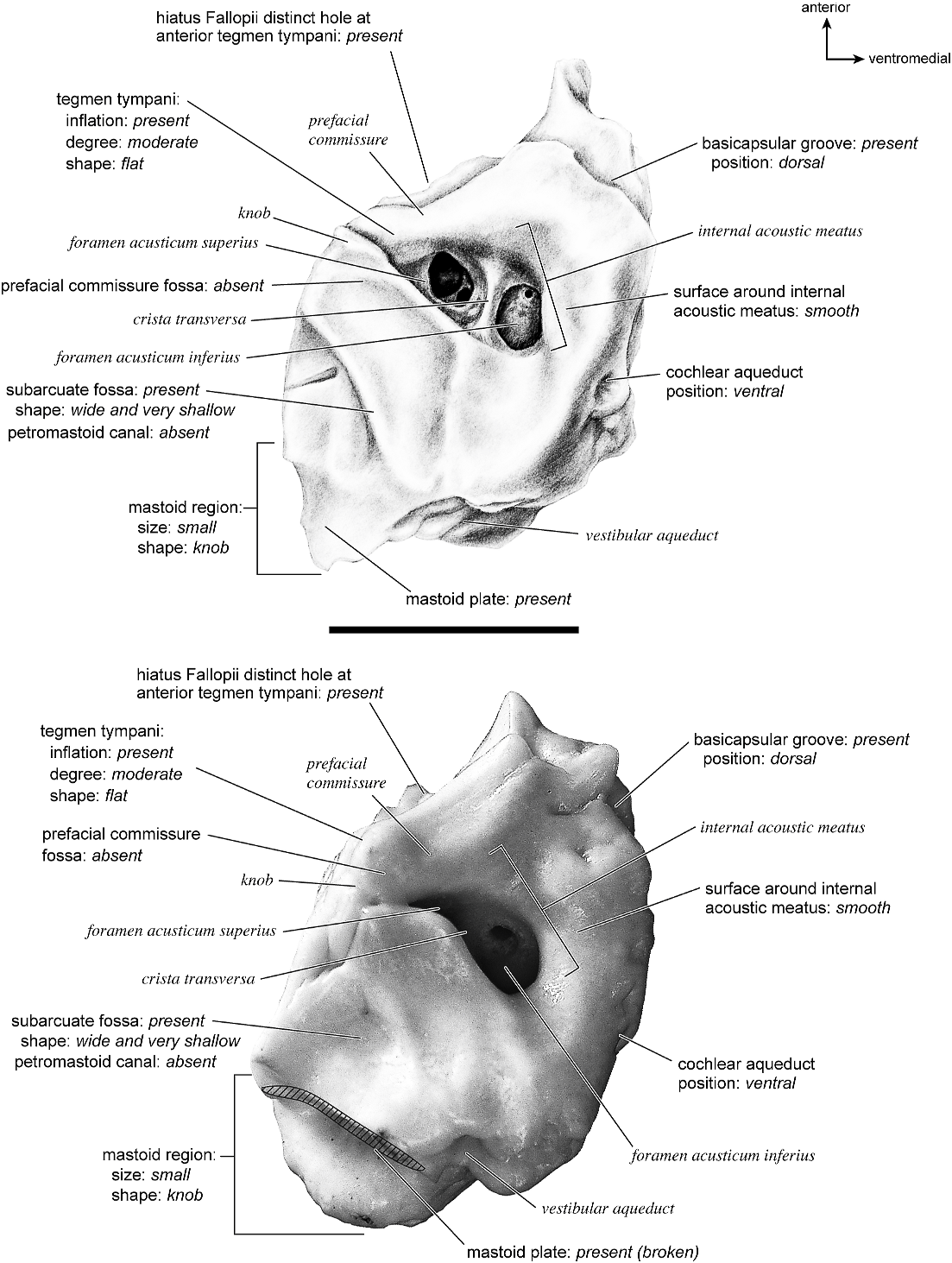
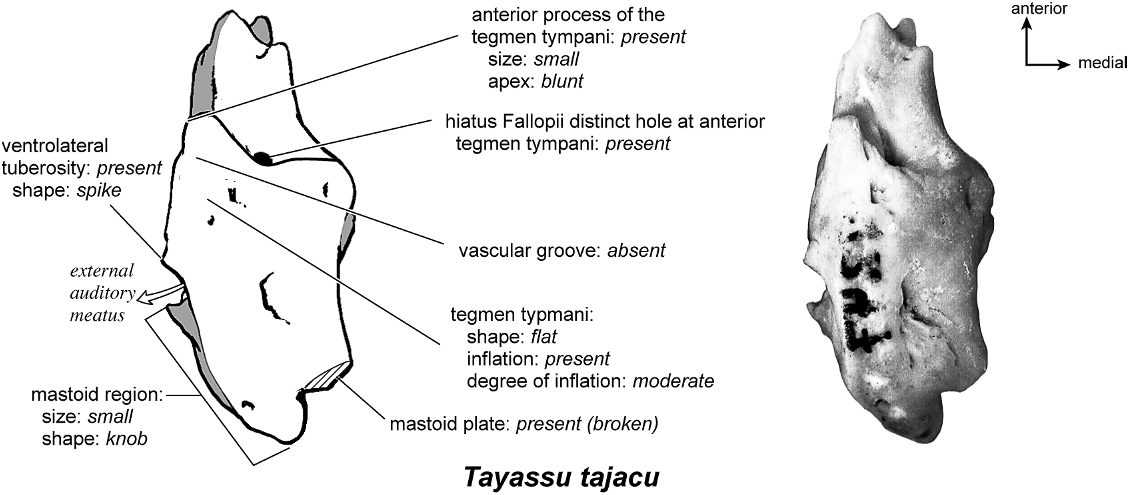
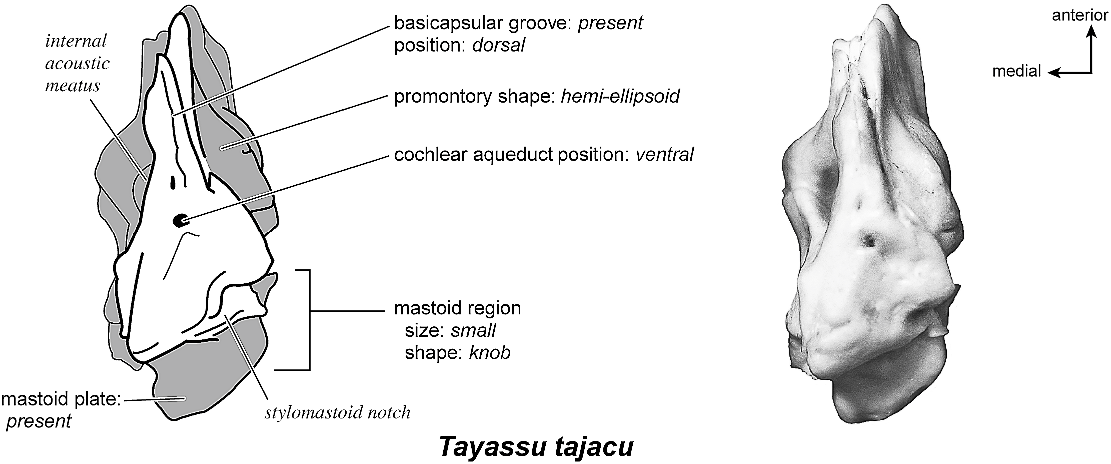
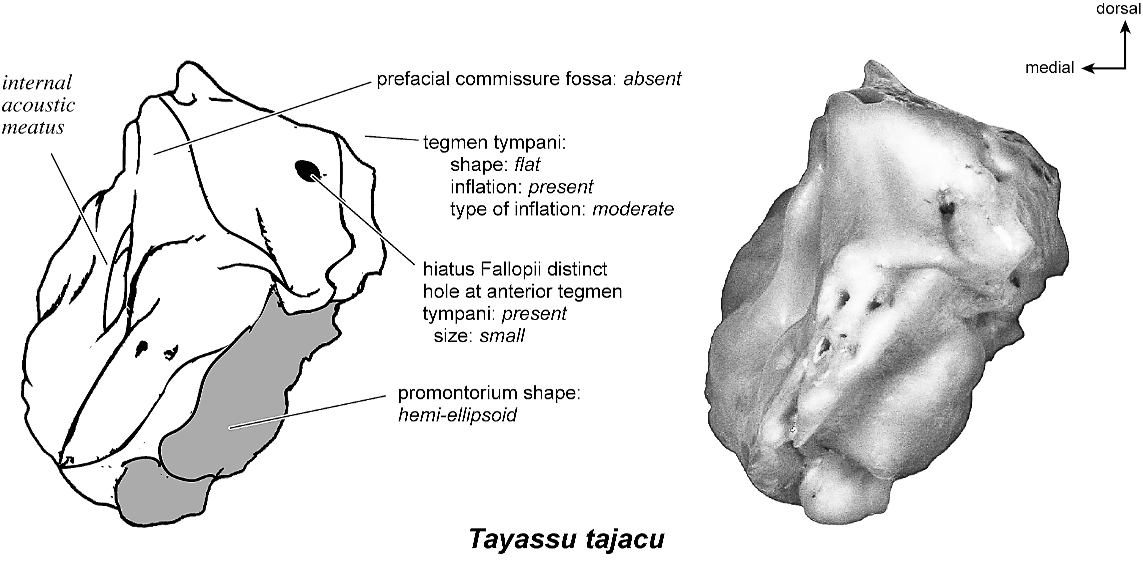

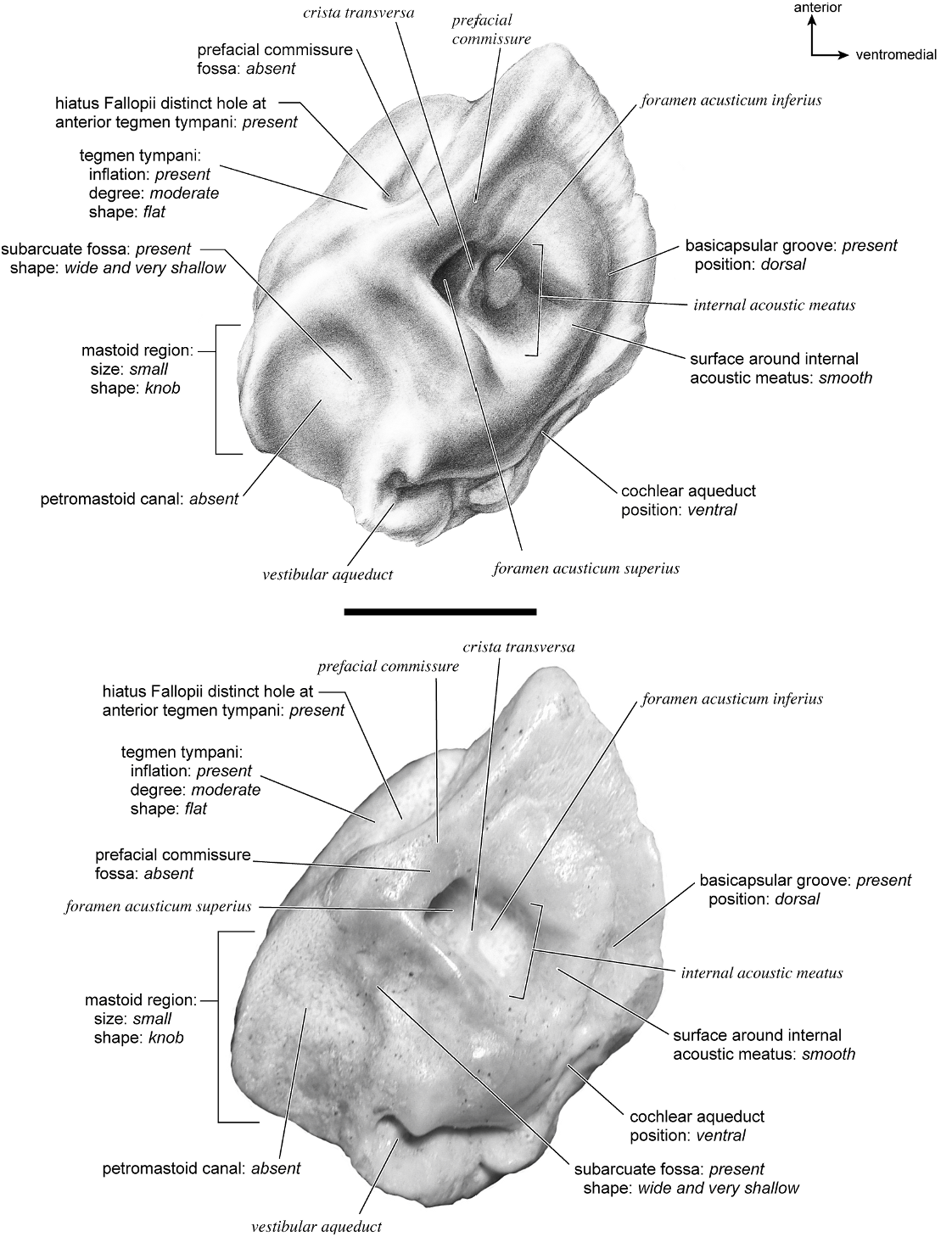

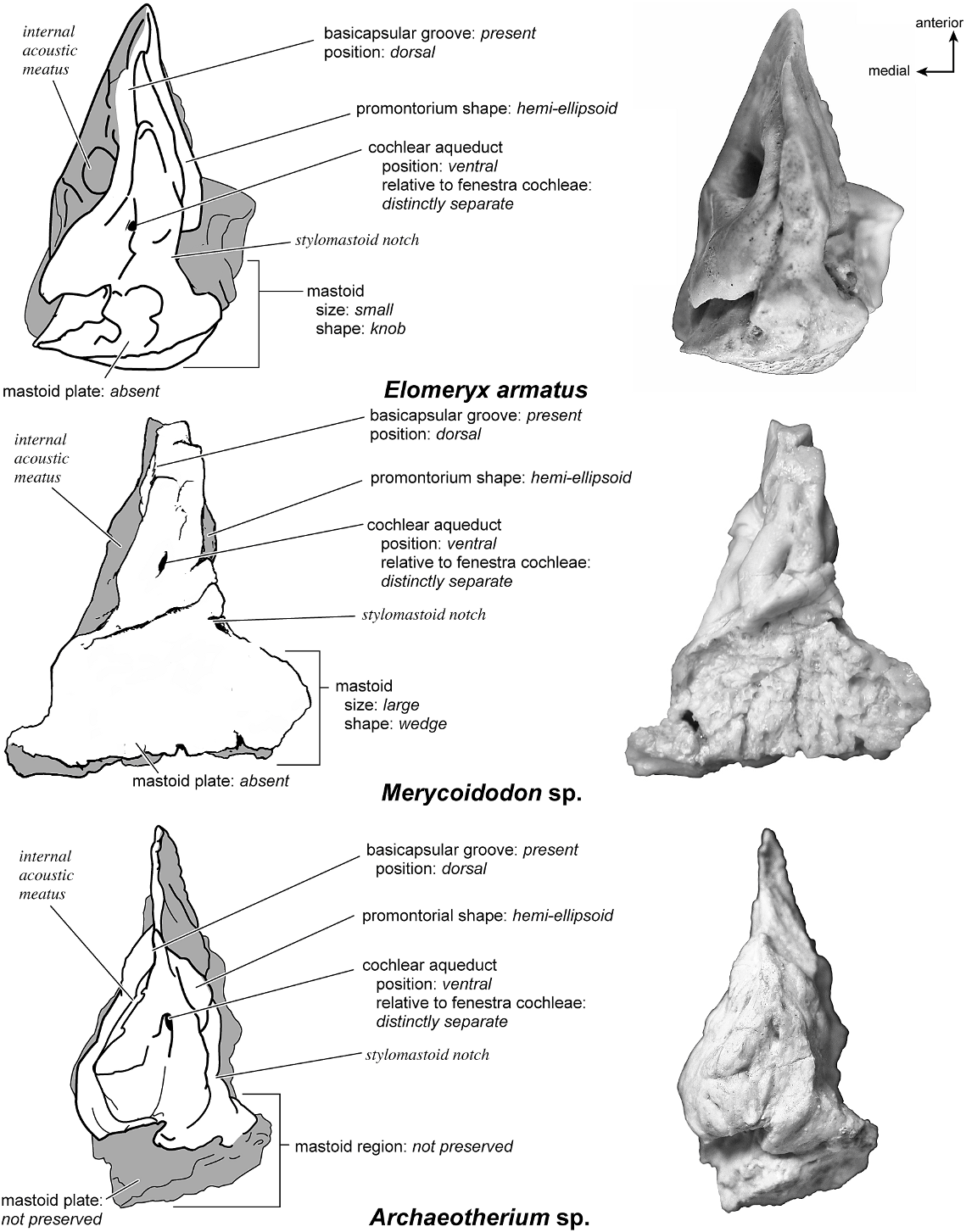

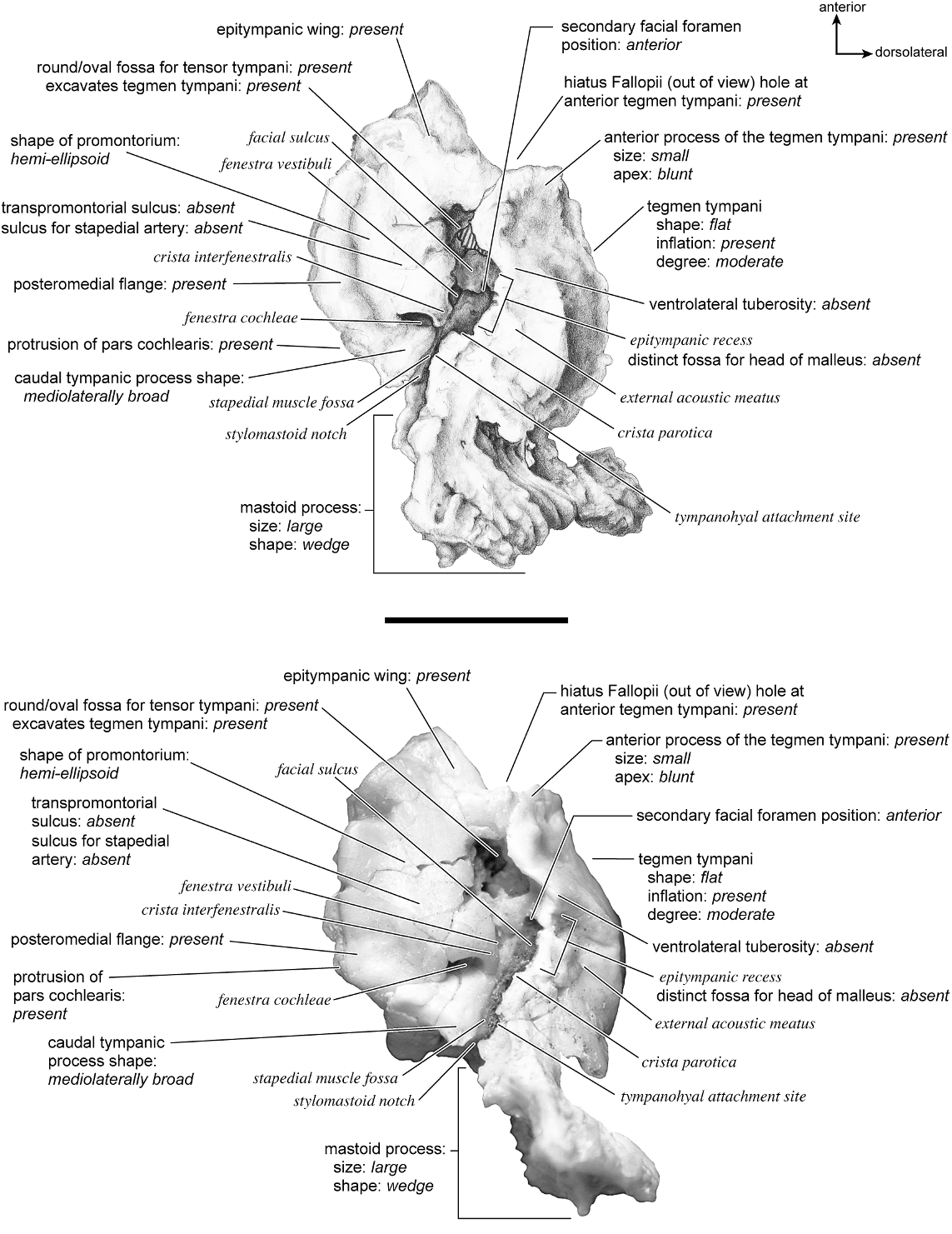




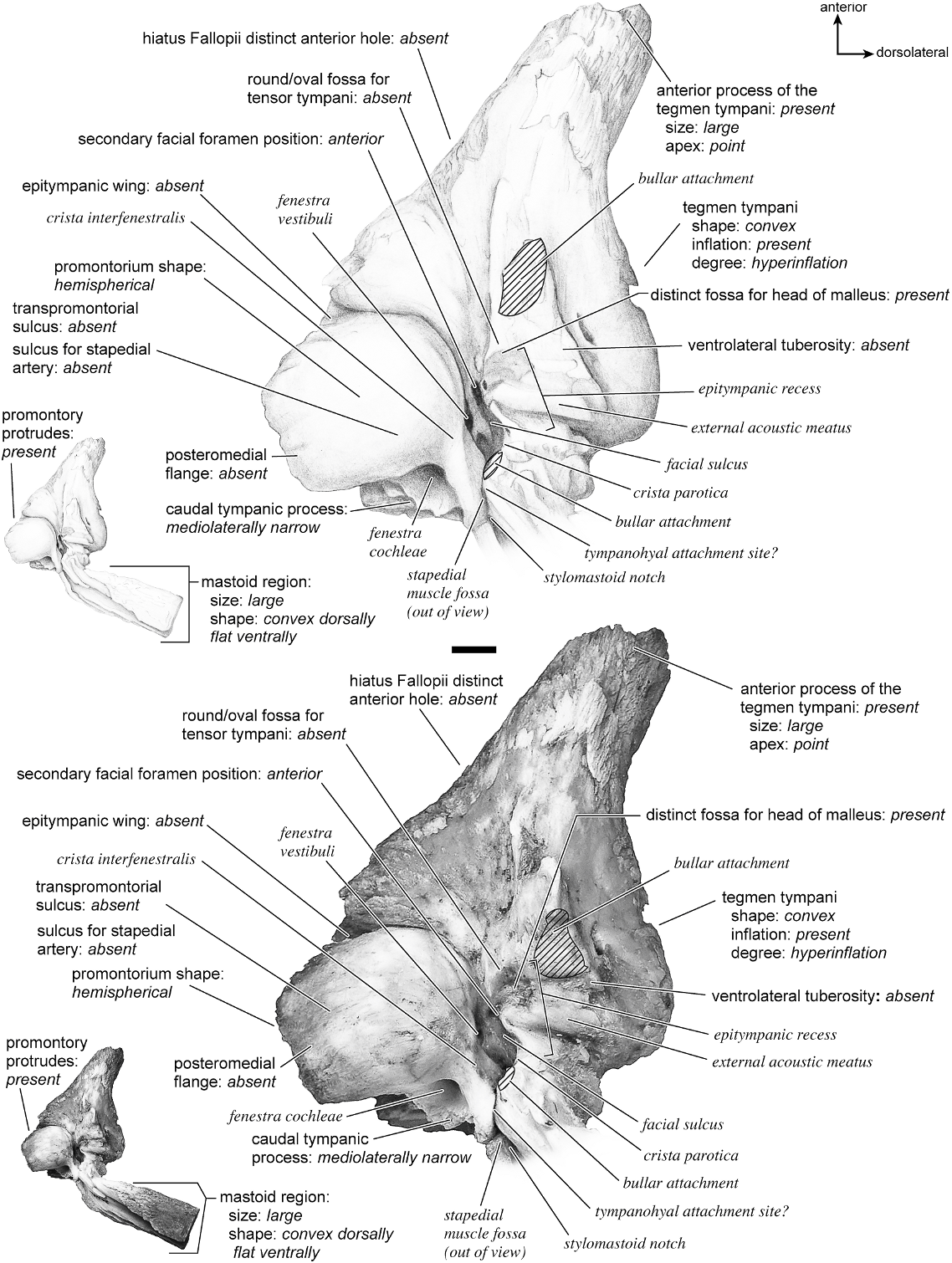

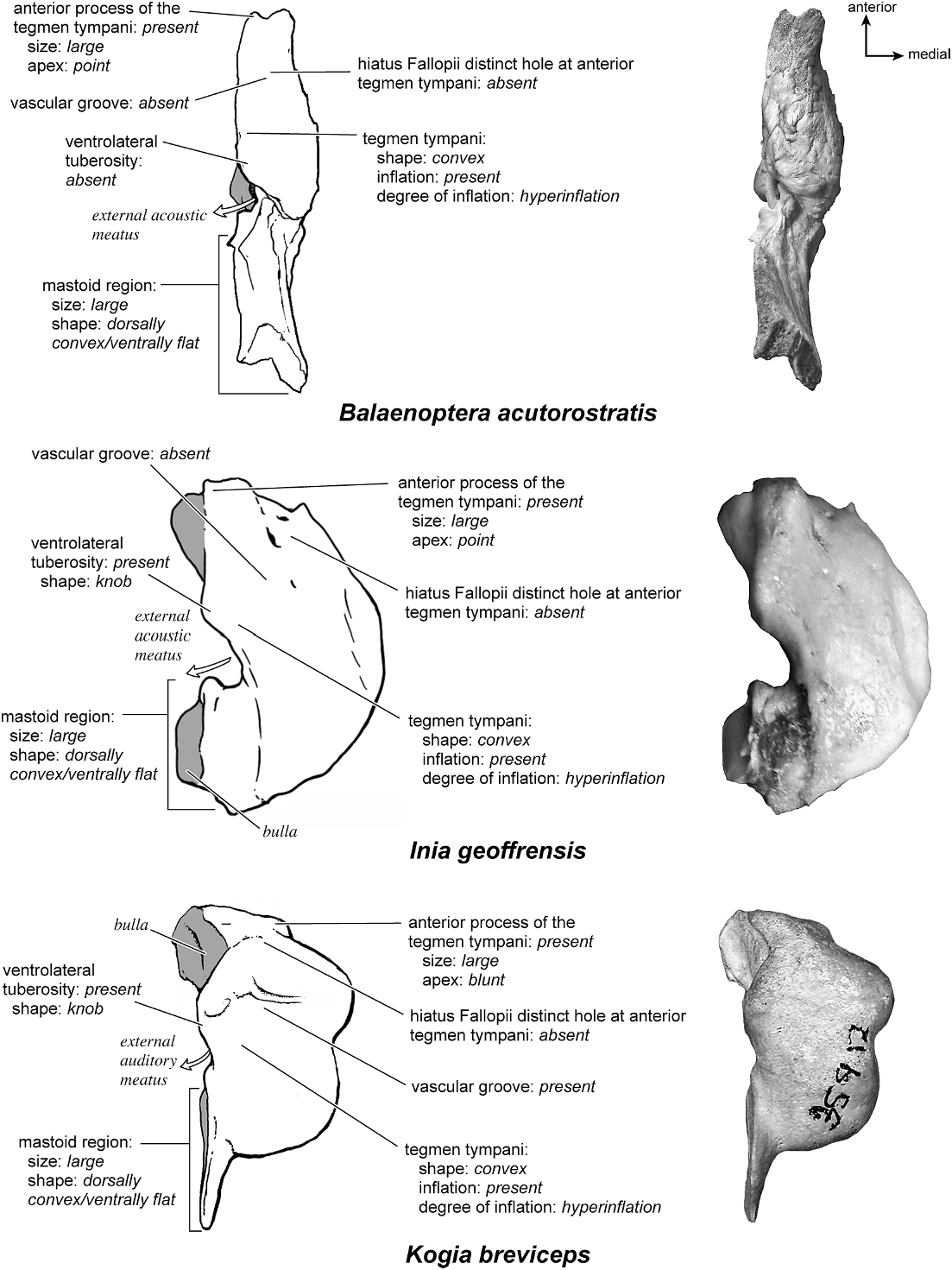


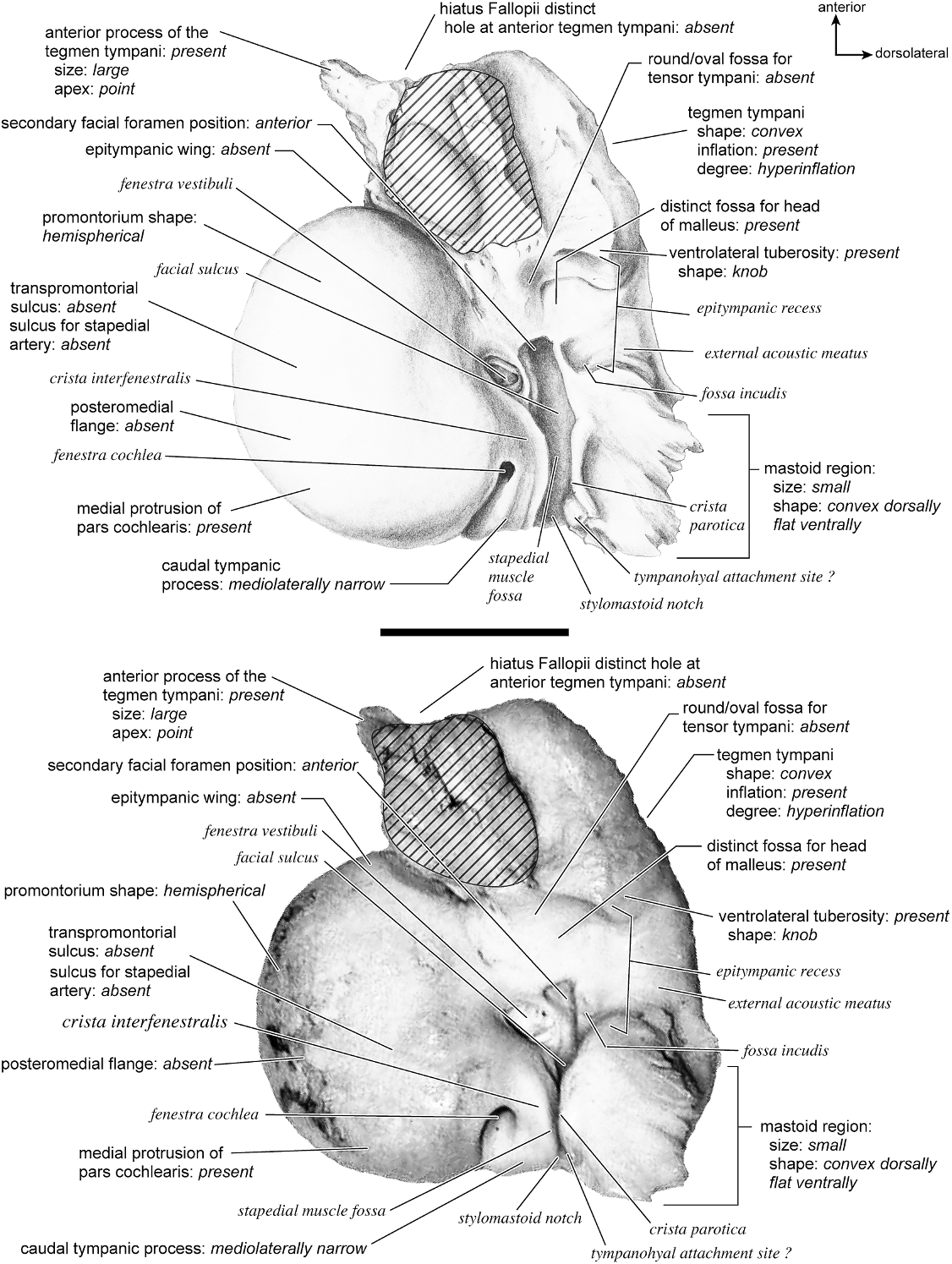
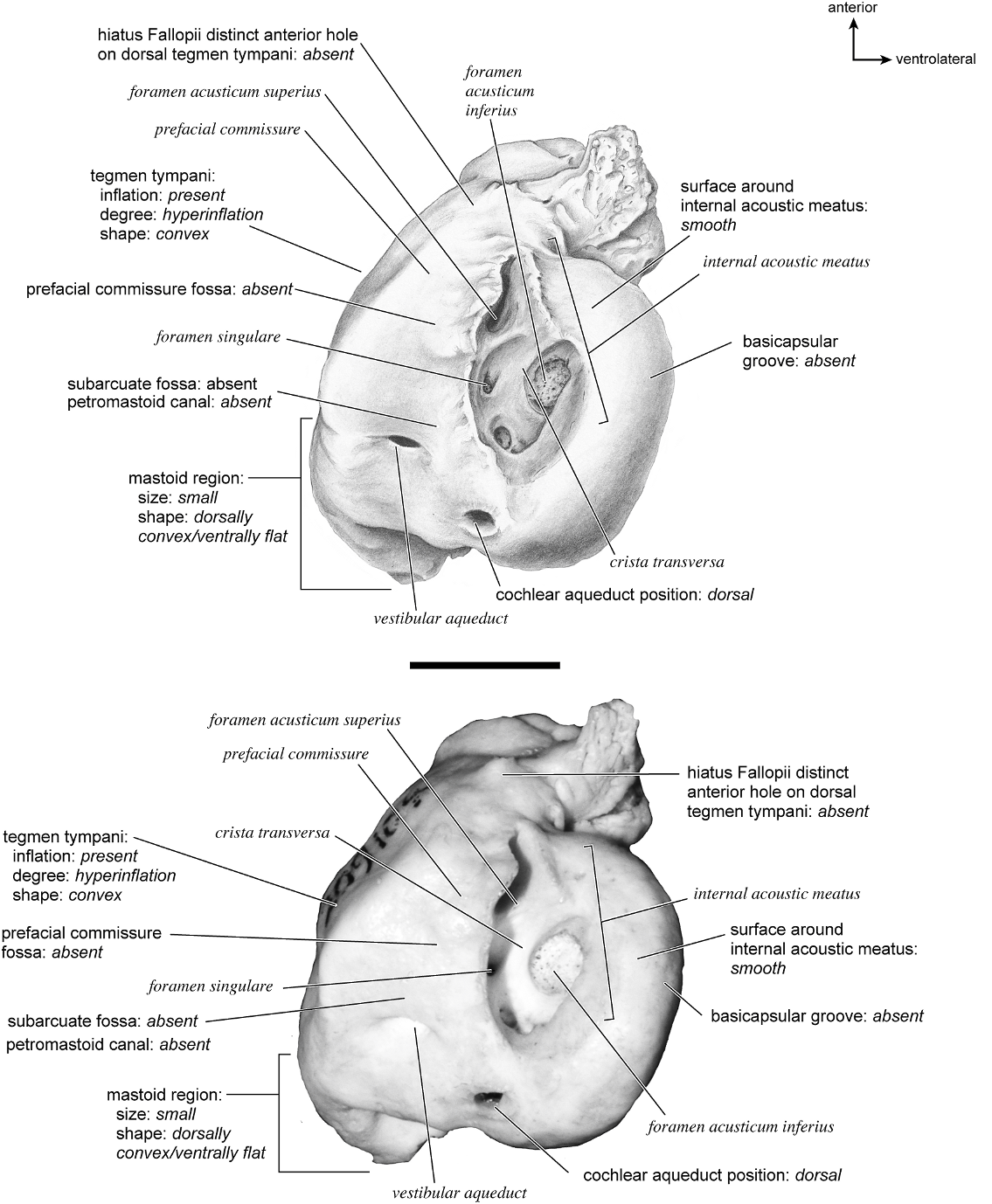
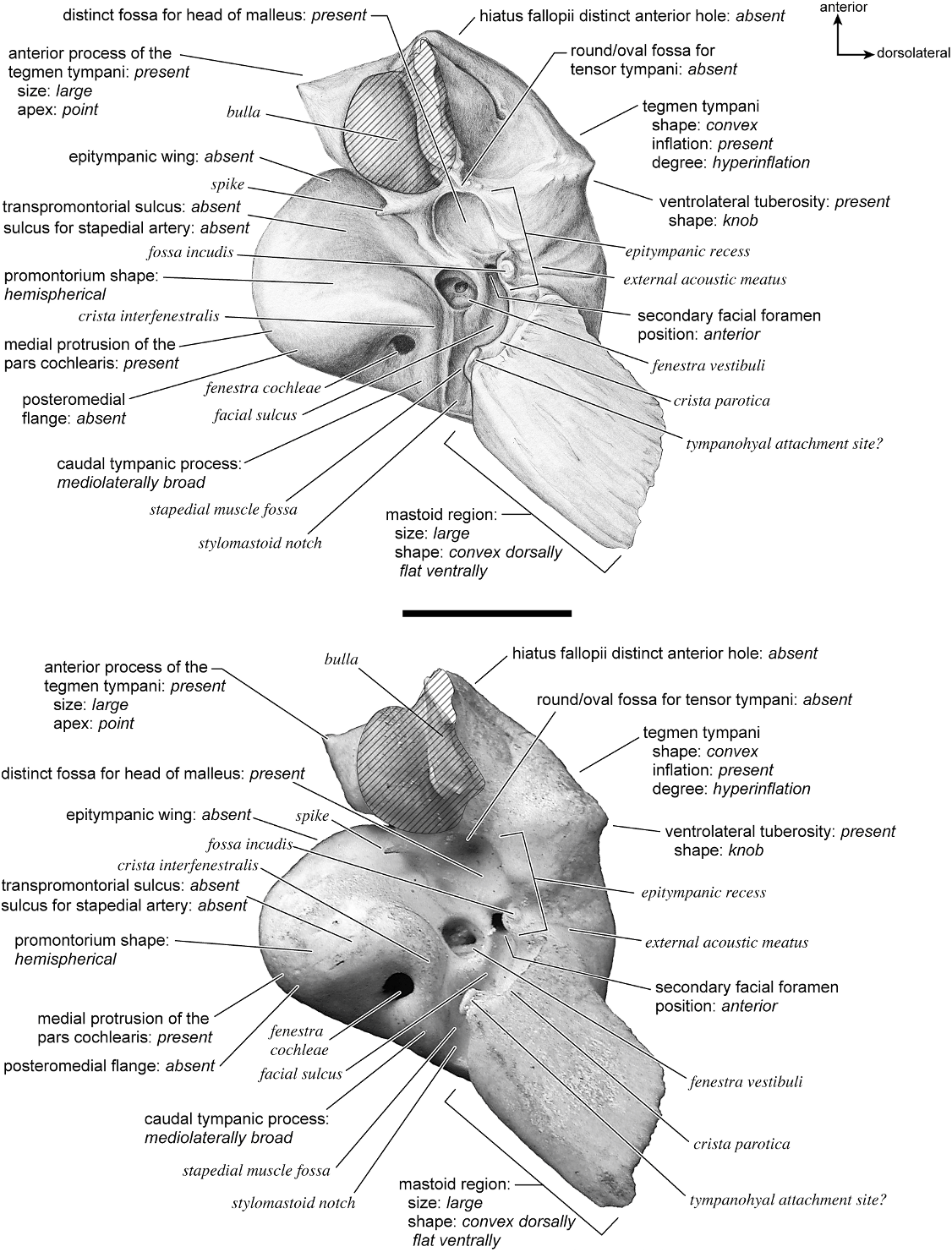
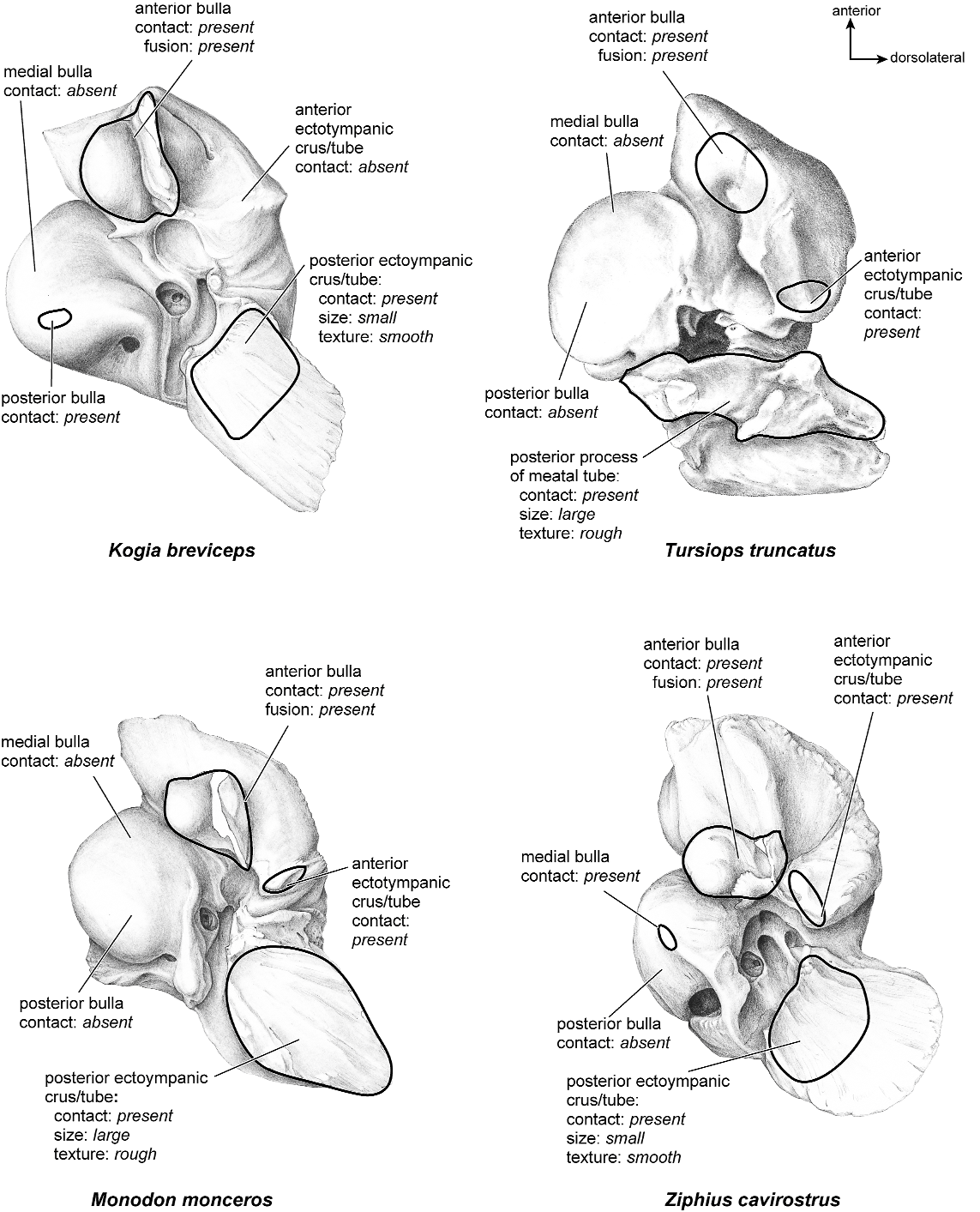
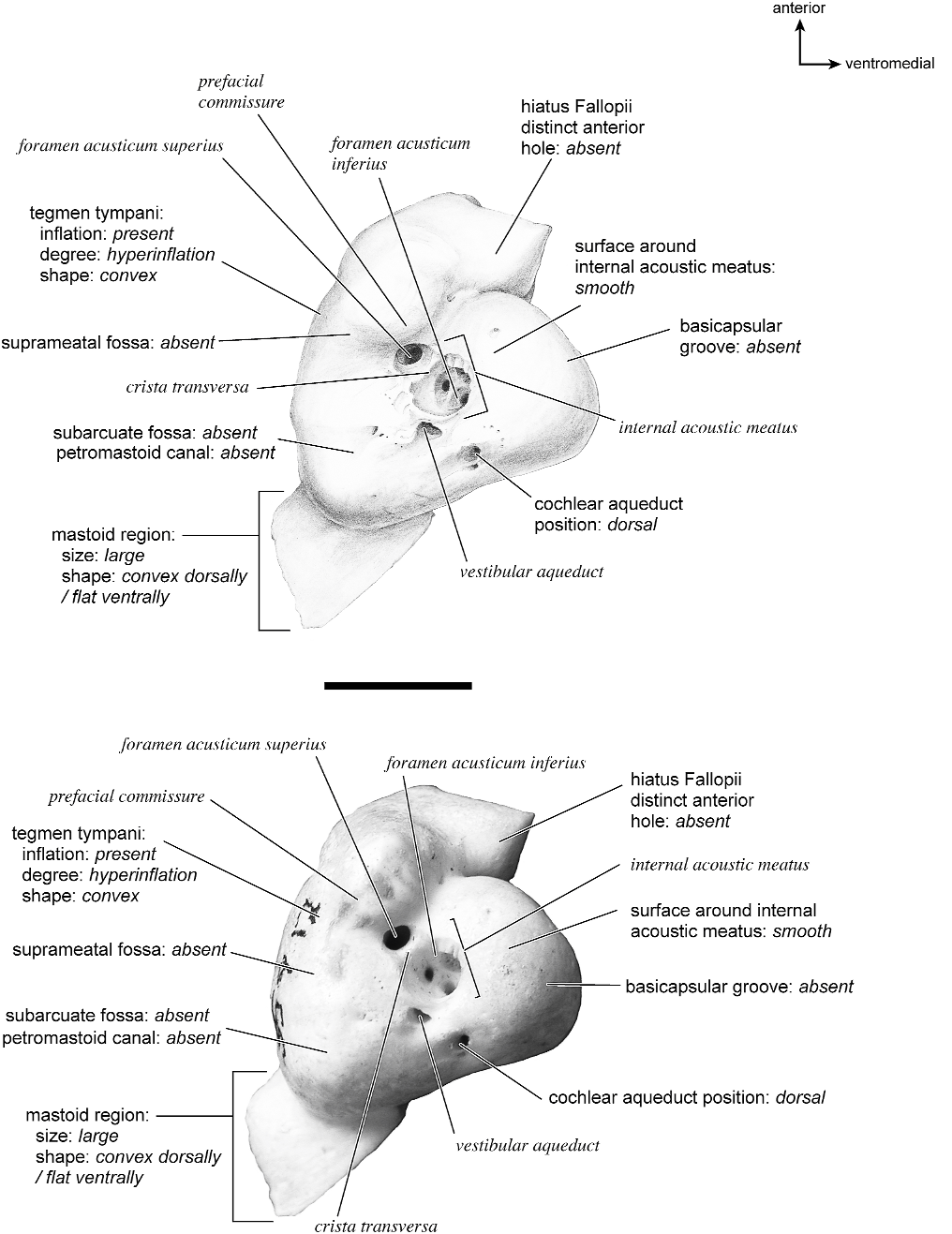
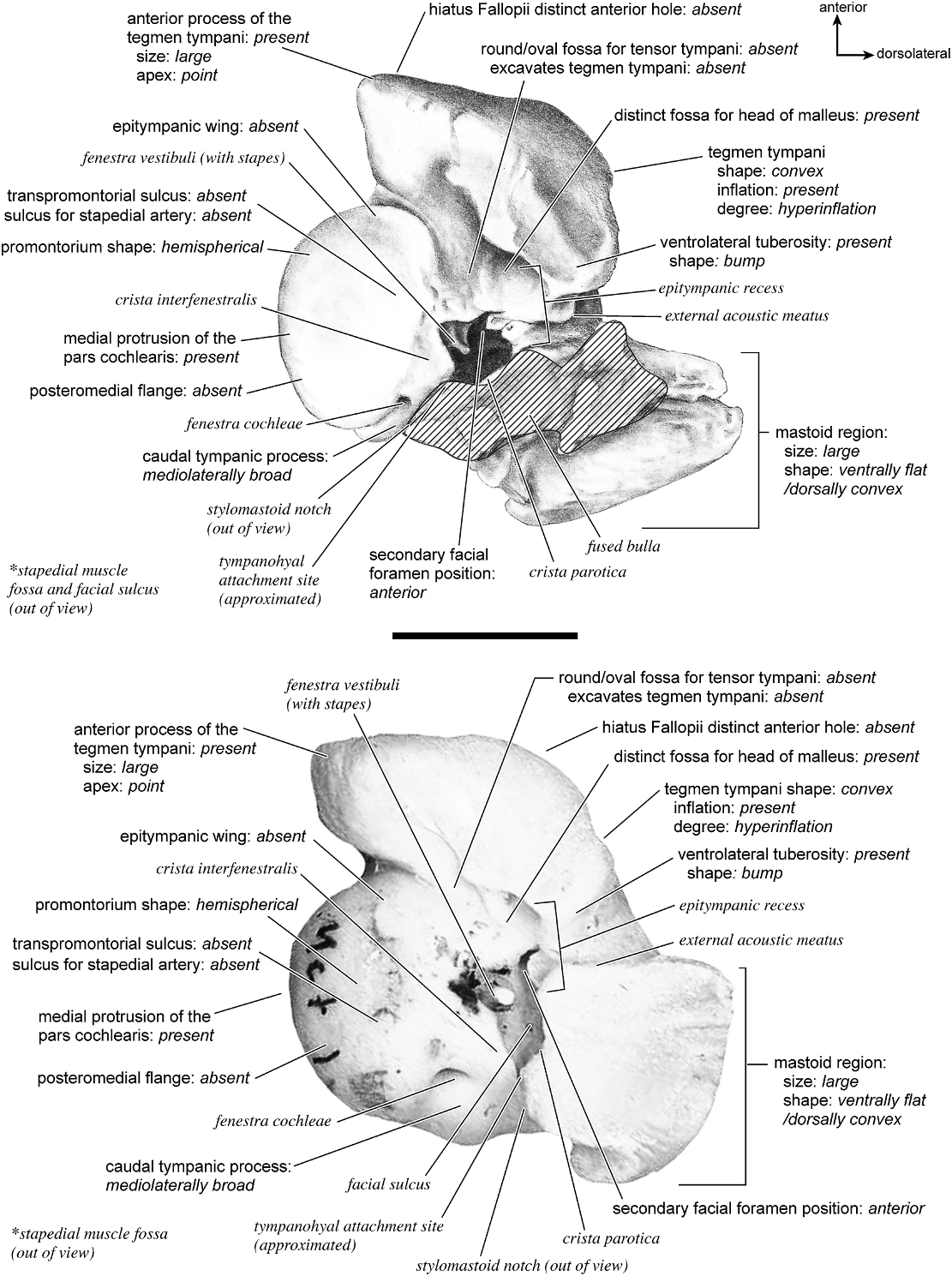

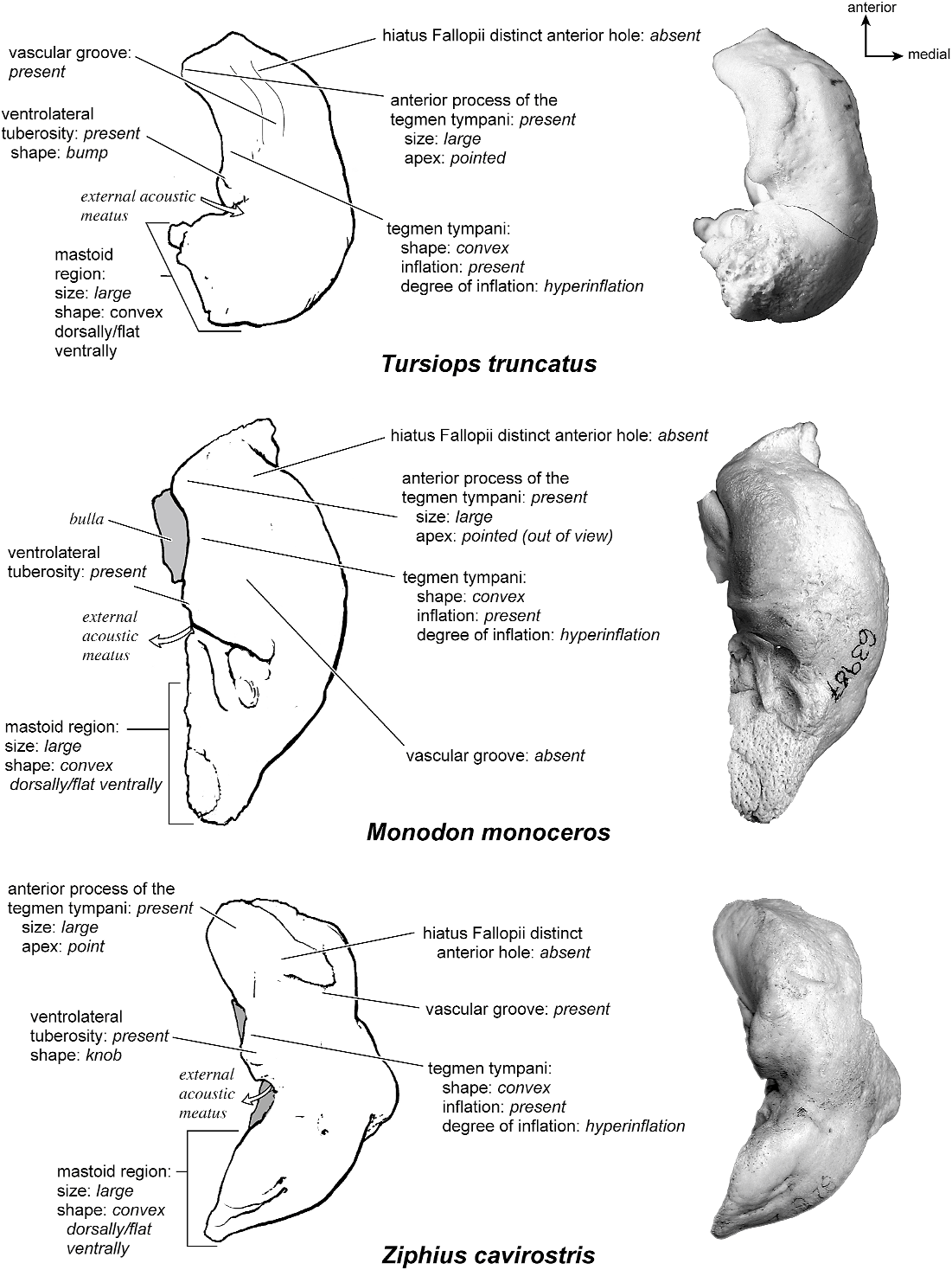


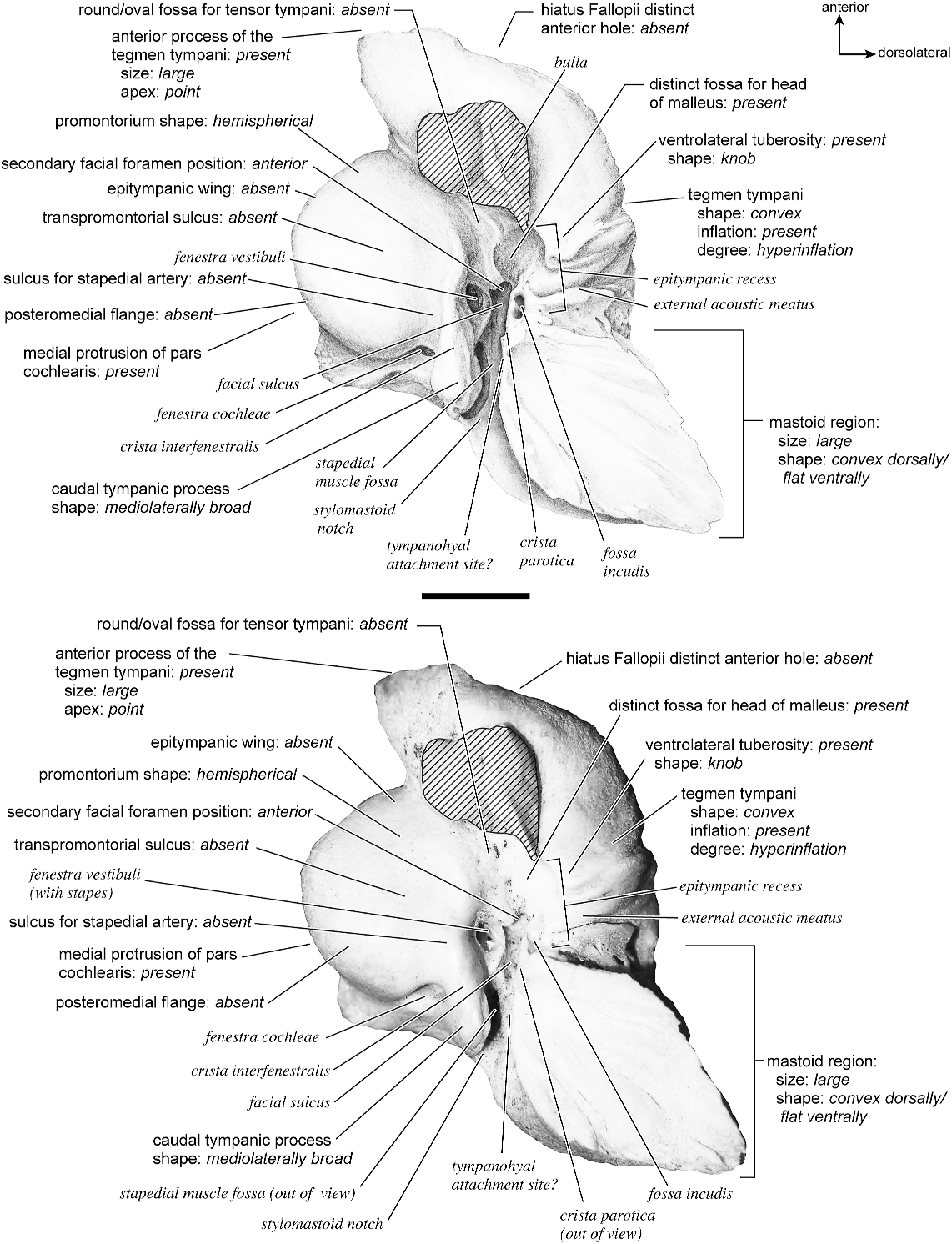
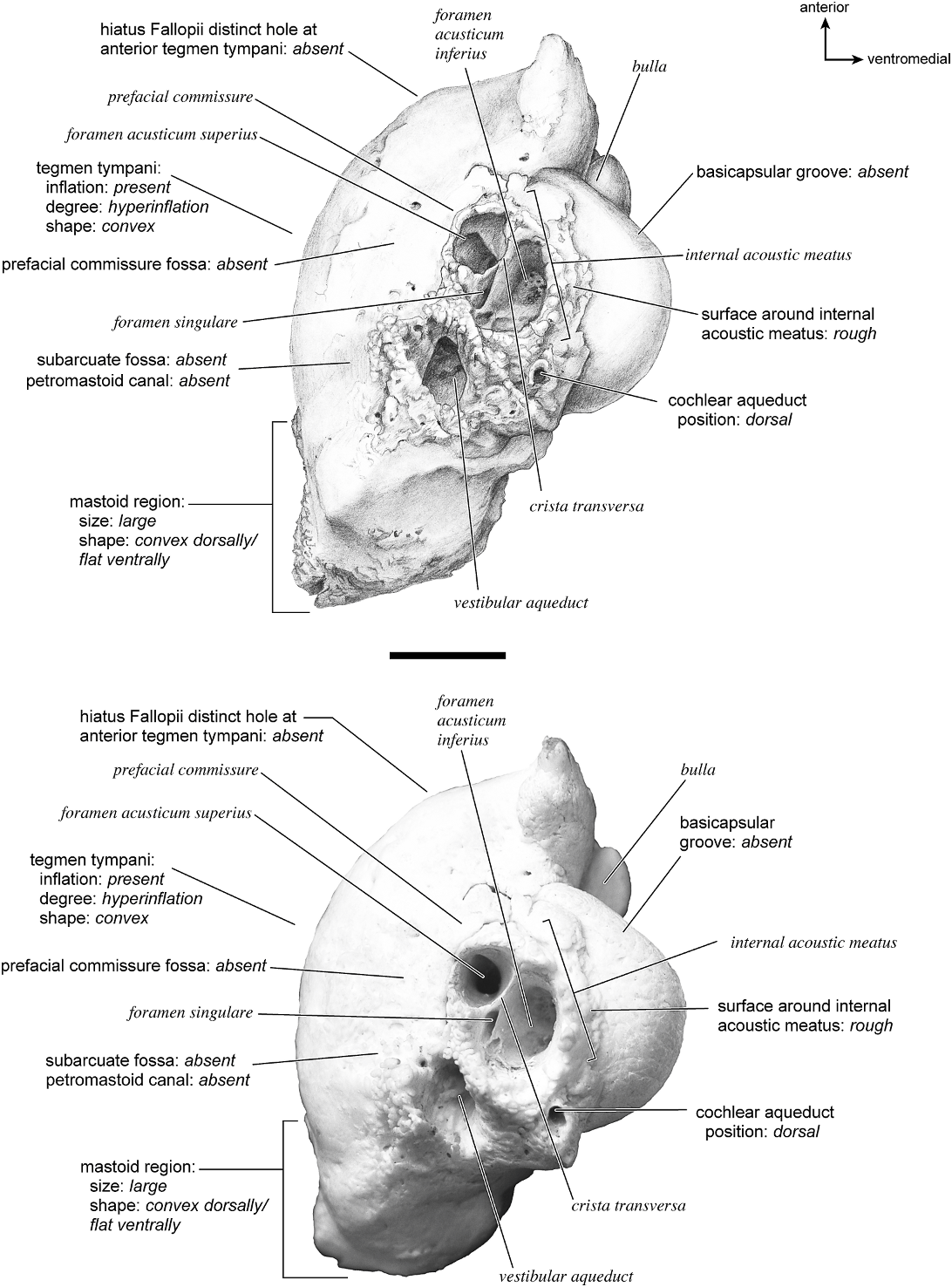


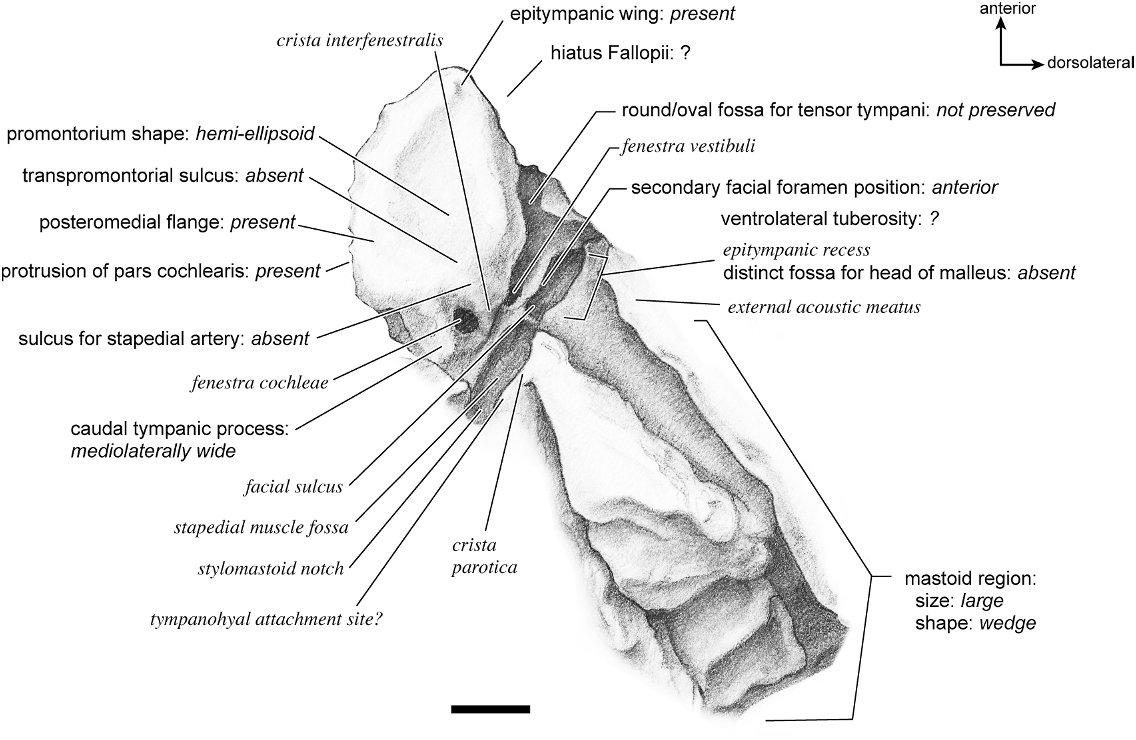
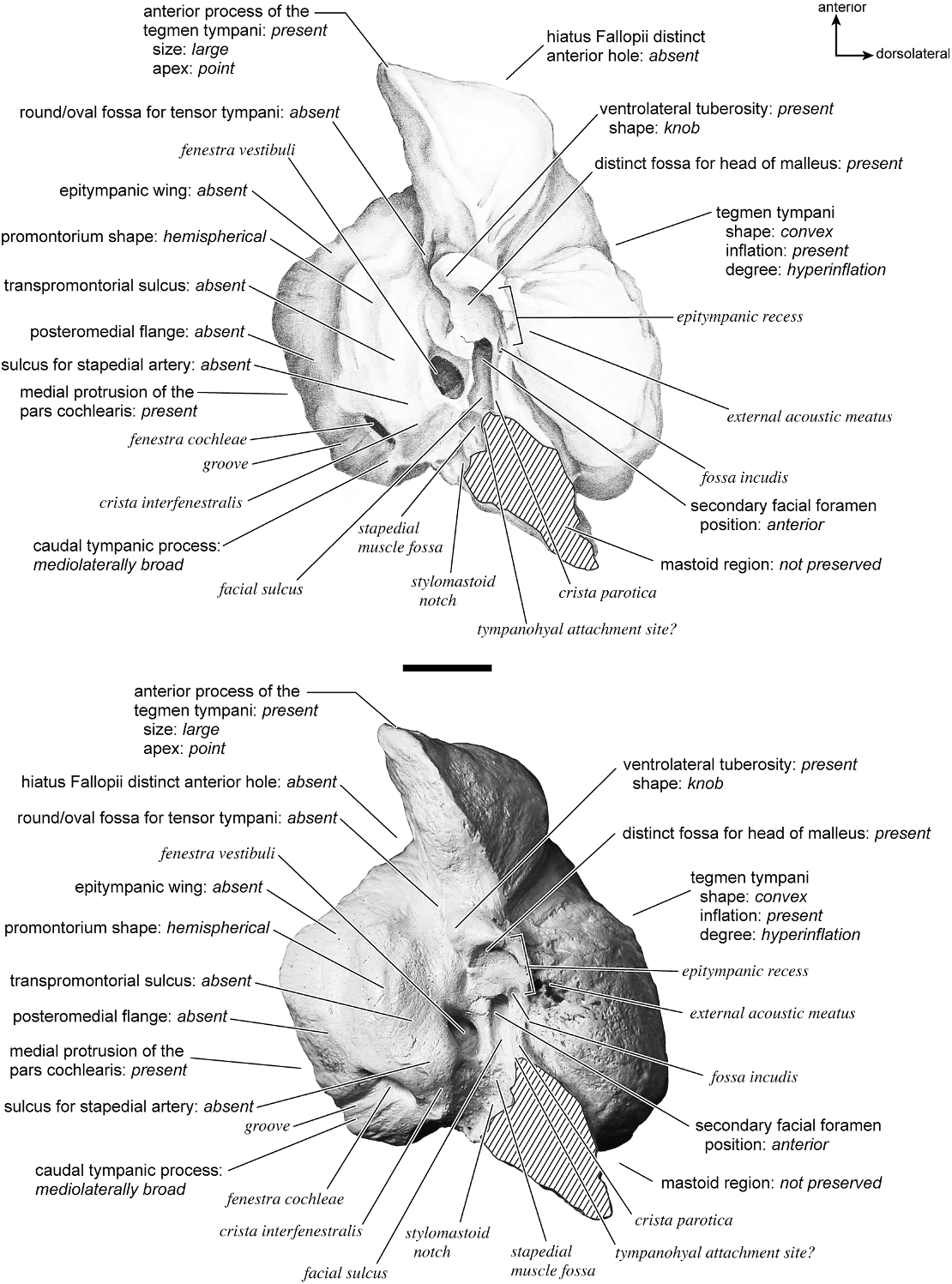
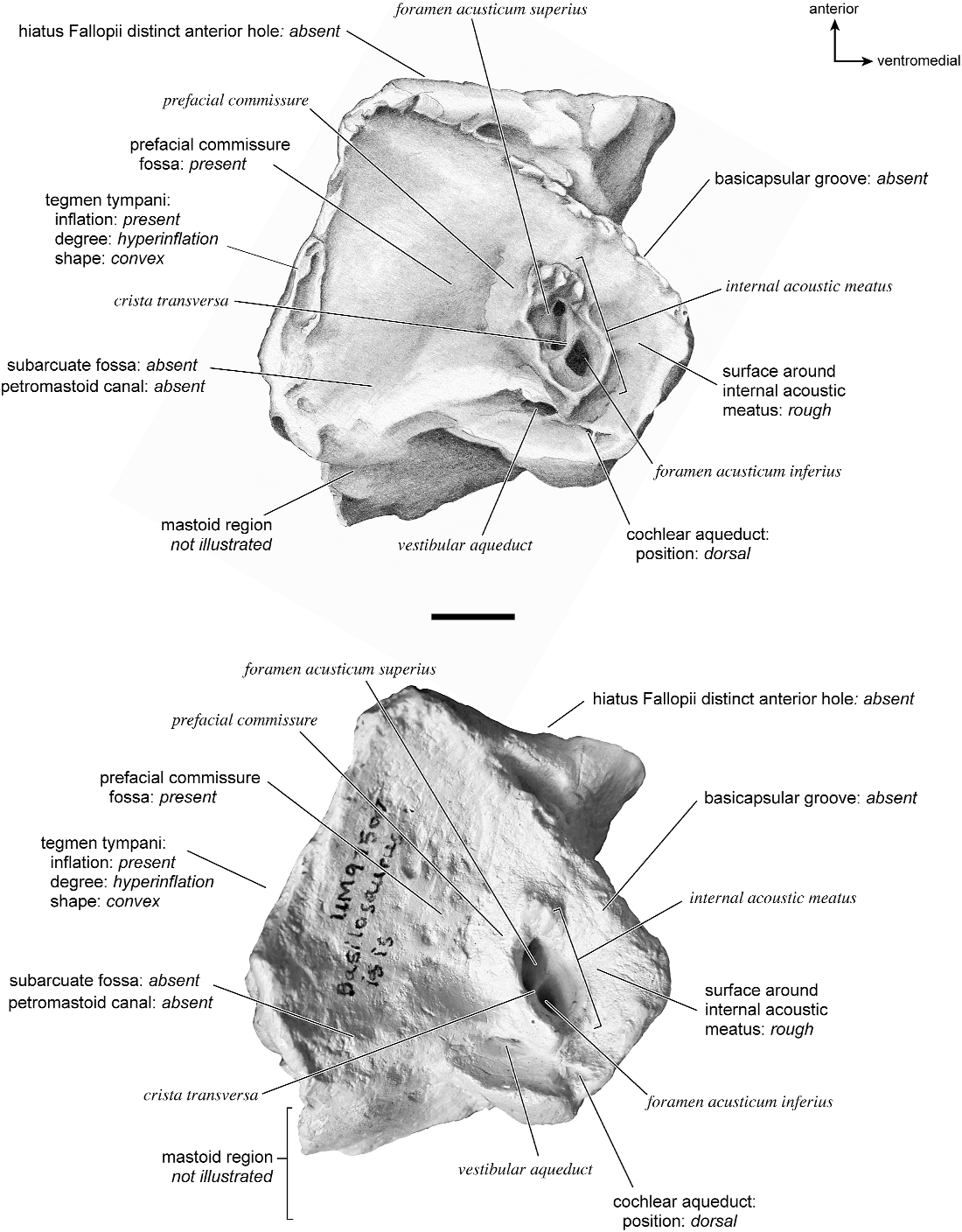
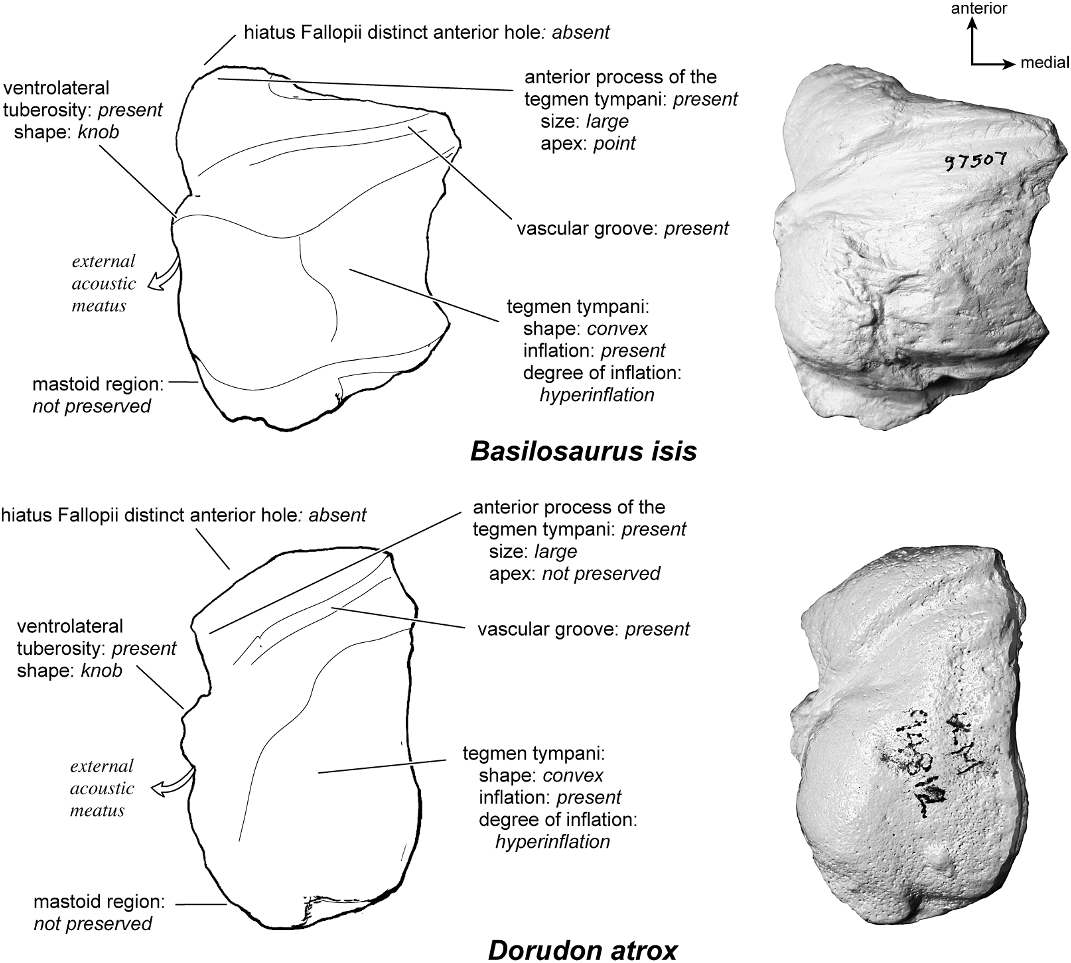


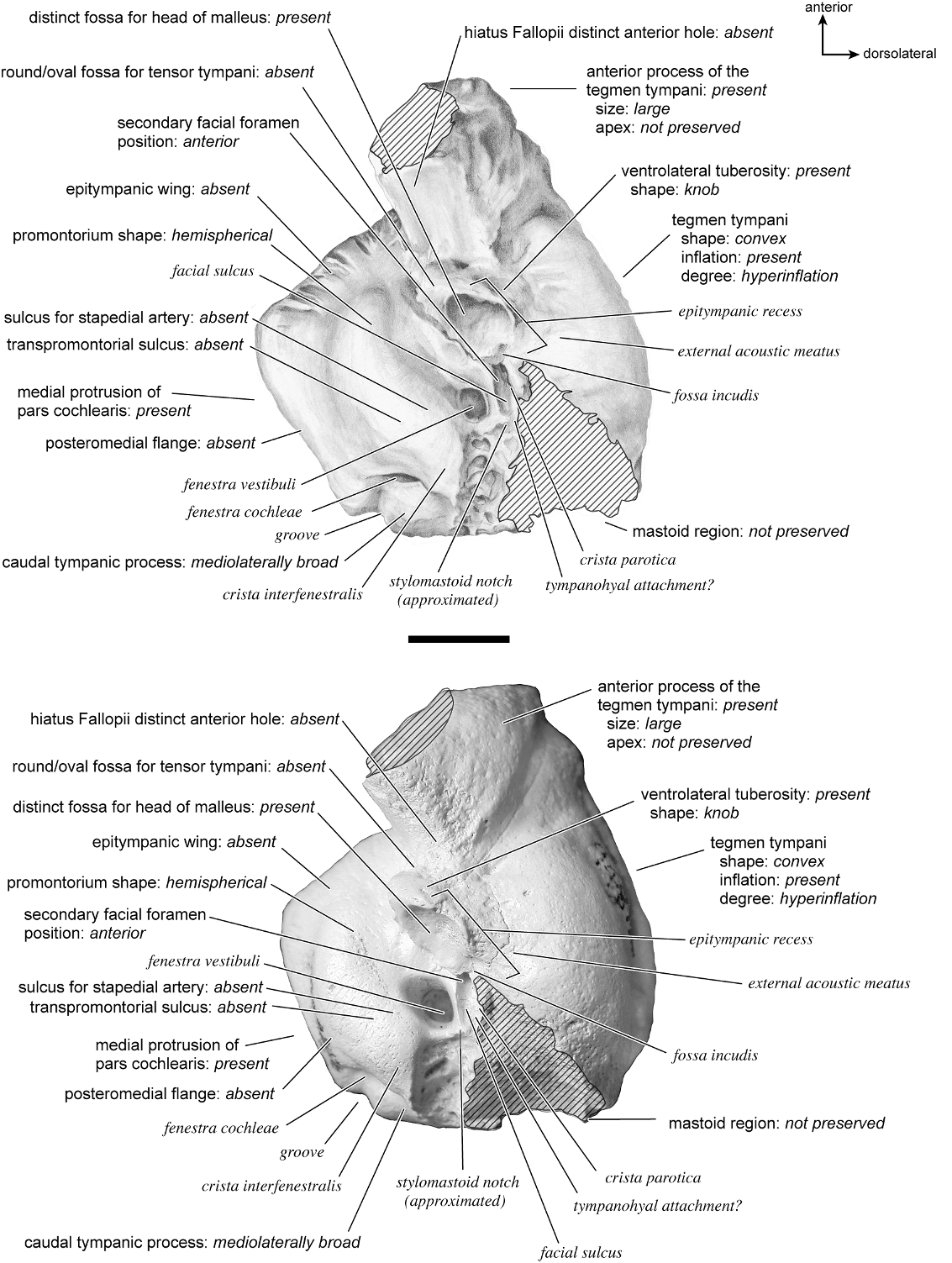


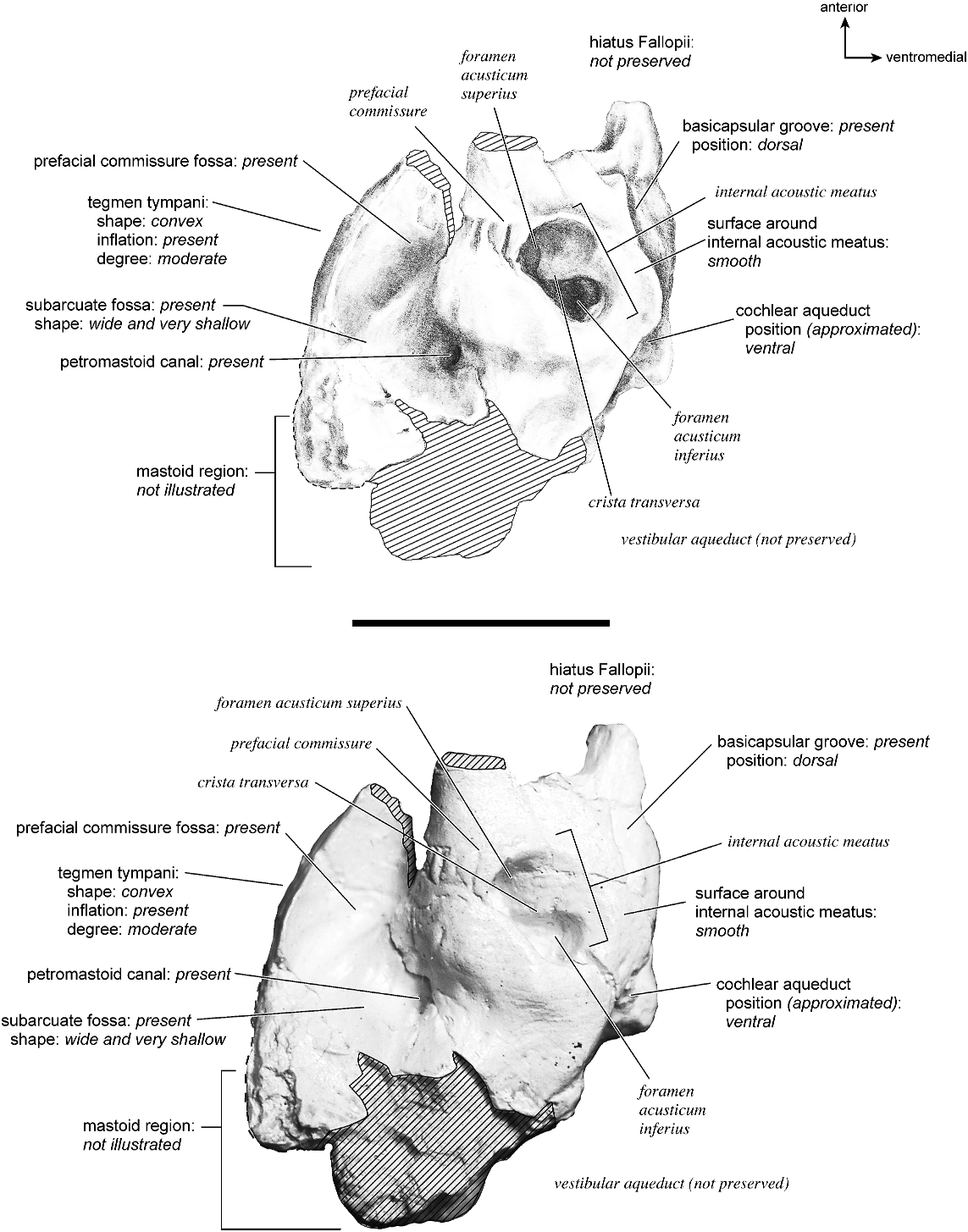


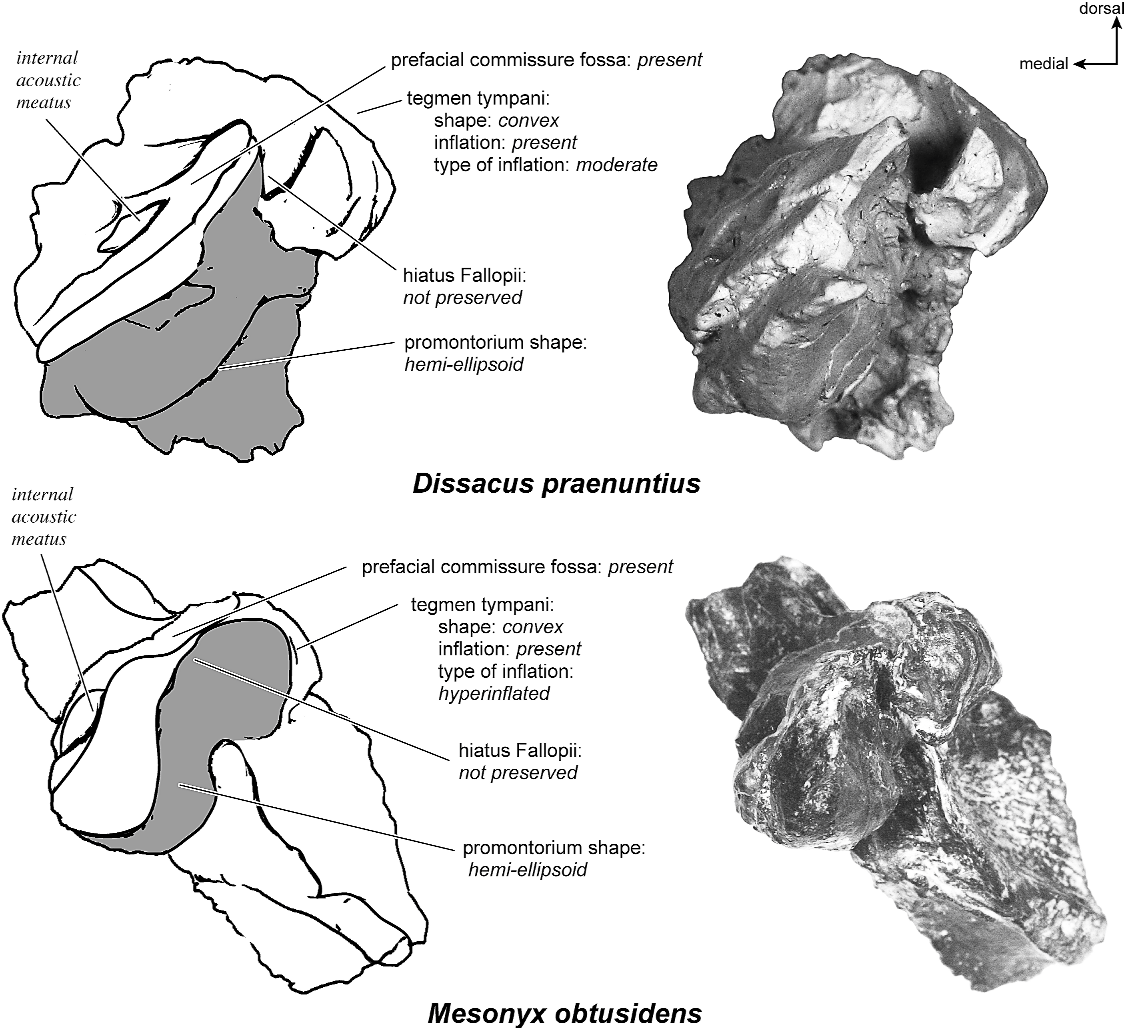
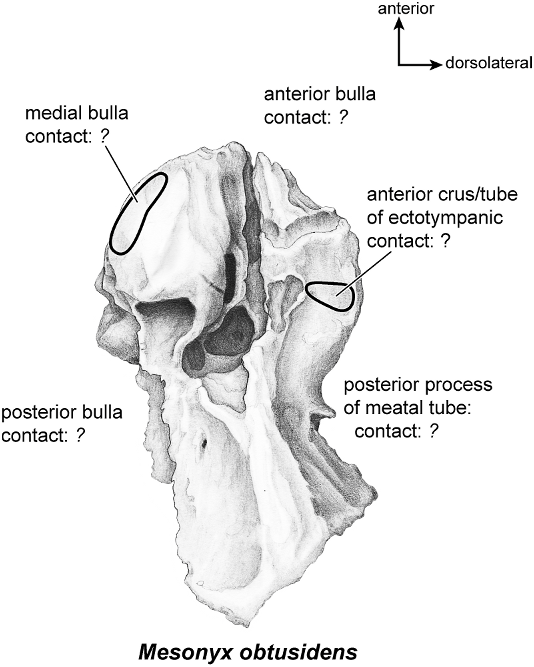
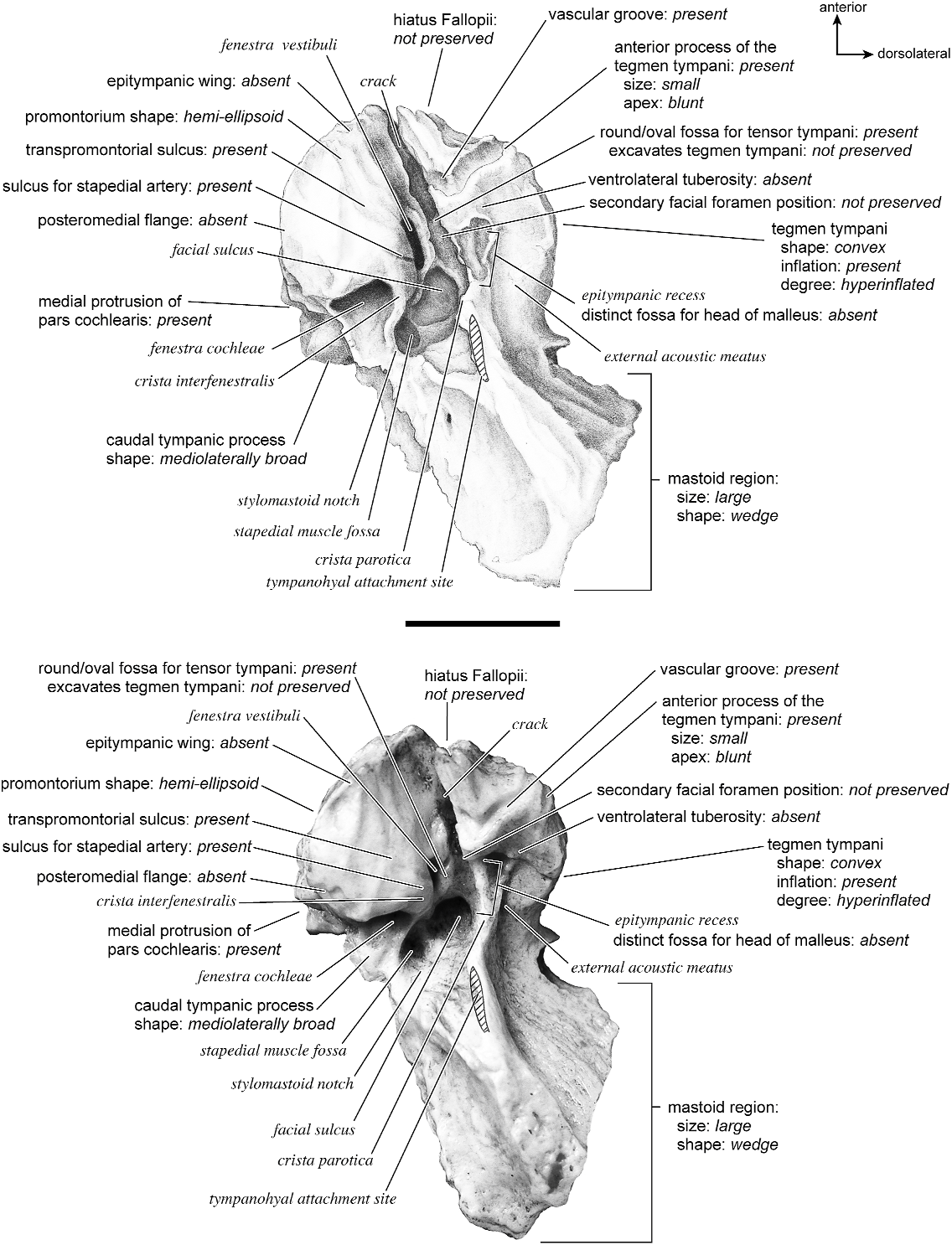


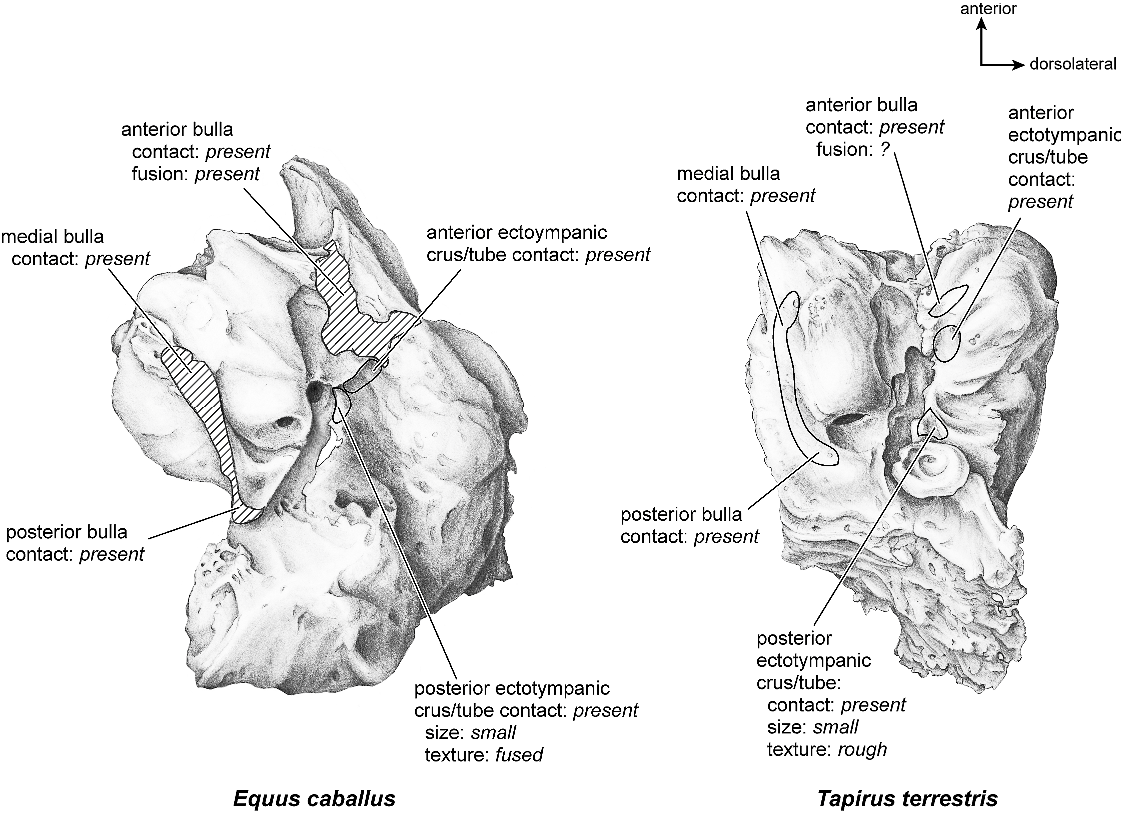

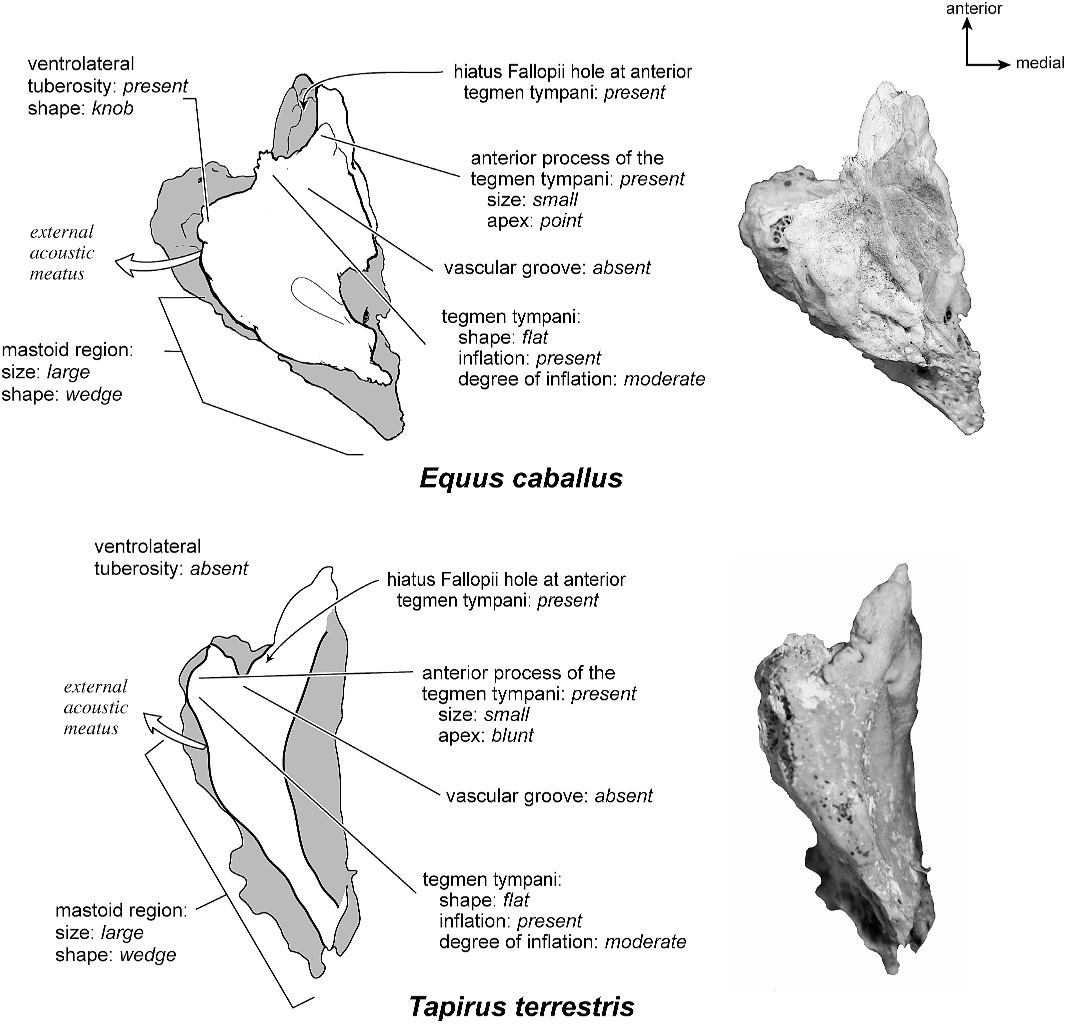
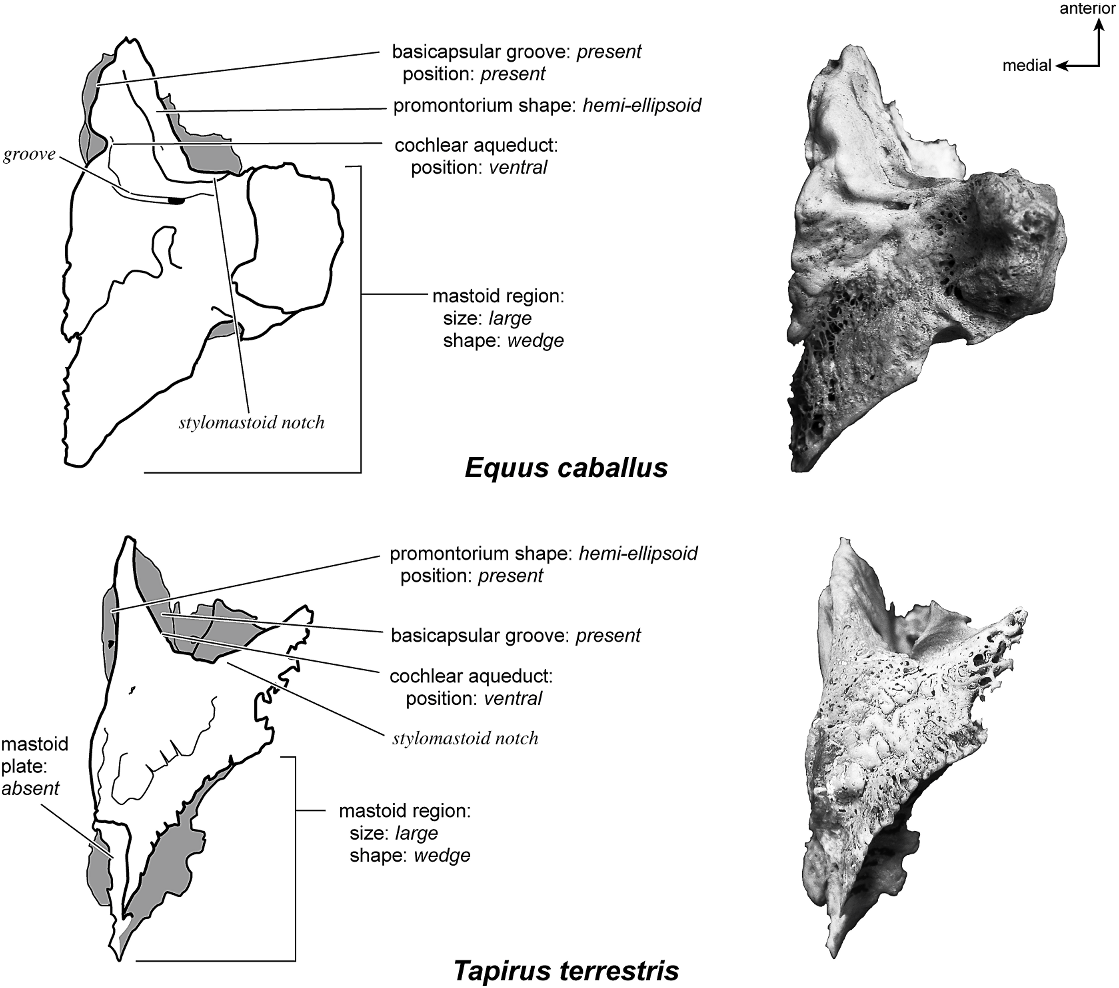


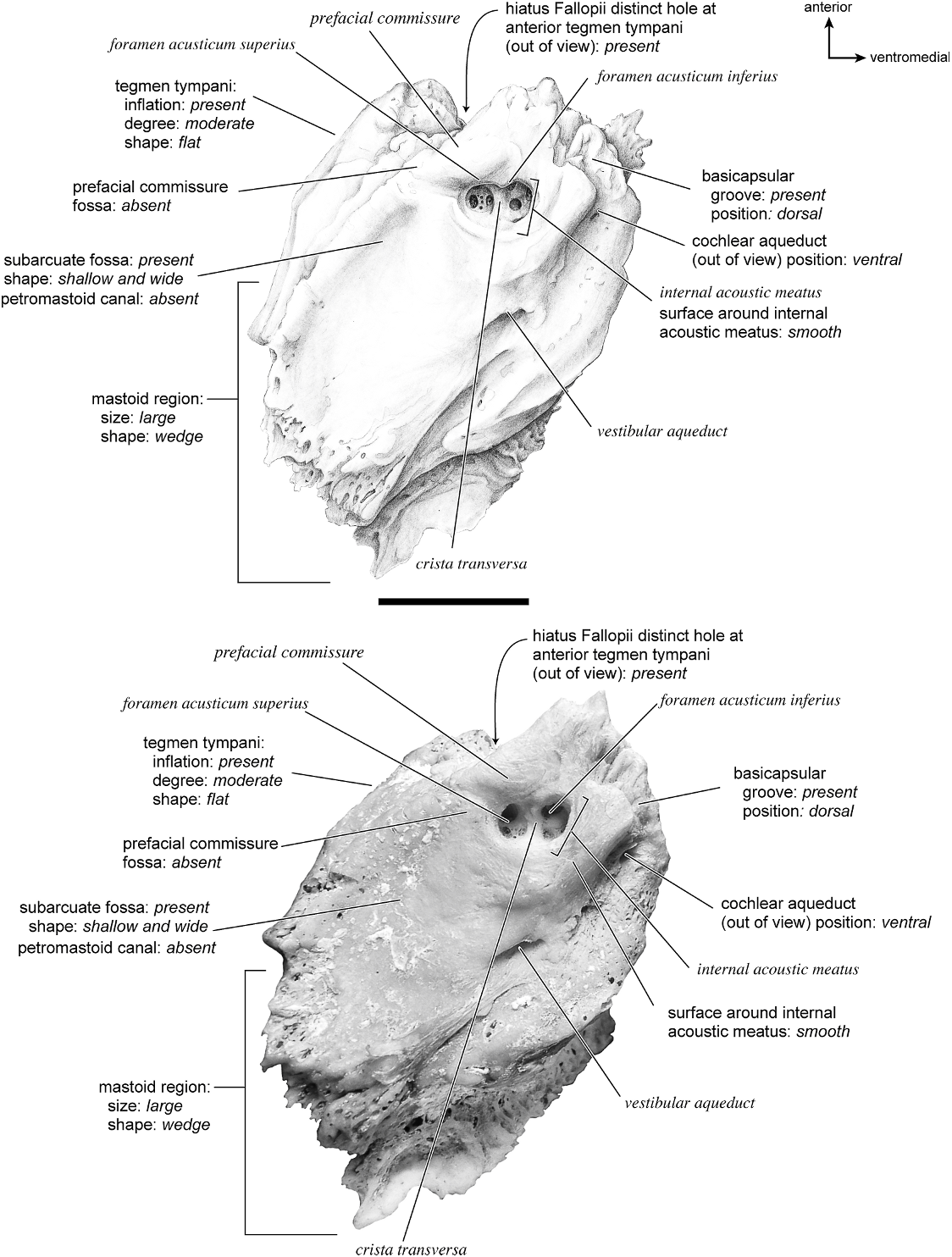
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.