
Trichotrigona camargoiana, Pedro, Silvia R. M. & Cordeiro, Guaraci Duran, 2015
|
publication ID |
https://doi.org/10.11646/zootaxa.3956.3.4 |
|
publication LSID |
lsid:zoobank.org:pub:6A8CA309-D8F4-44B6-BE13-7C53A23E62FB |
|
DOI |
https://doi.org/10.5281/zenodo.6118163 |
|
persistent identifier |
https://treatment.plazi.org/id/874DD440-2D2E-D977-9189-AD4A7EC5D9F2 |
|
treatment provided by |
Plazi |
|
scientific name |
Trichotrigona camargoiana |
| status |
sp. nov. |
Trichotrigona camargoiana View in CoL sp. nov.
Diagnosis. Worker ( Figs. 1, 5, 9, 11 View FIGURES 1 – 11 , 12, 14, 16–21 View FIGURES 12 – 21 ; Tab. I). Front and mesepisterna laterally with plumose setae intermingled with simple; anterior wings bright yellow; setae of tibia III predominantly black; maculation on thorax bright yellow, lateral stripes of mesoscutum from anterior corners to axillae; axillae entirely yellow; tibiae III black.
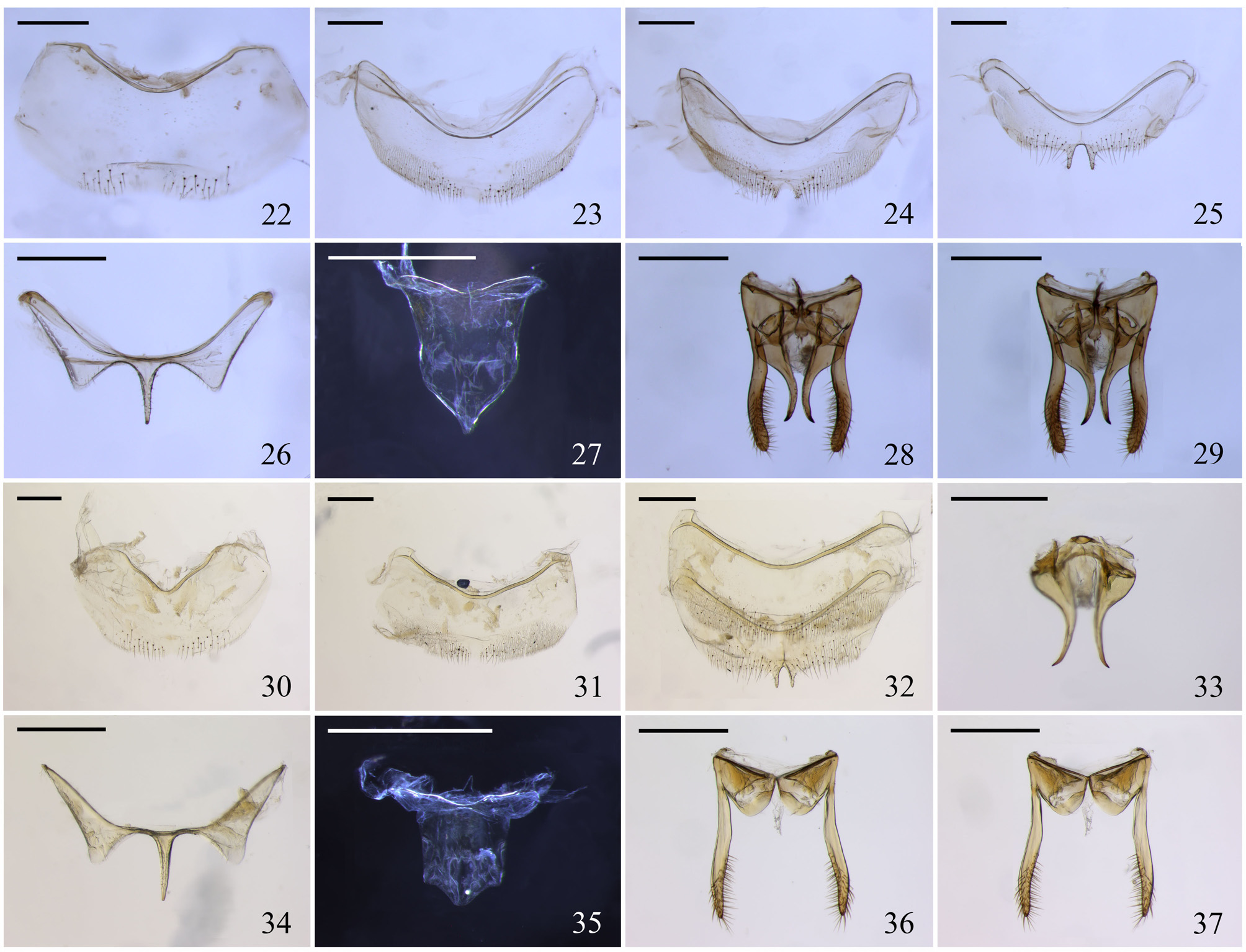
Male ( Figs. 3, 6 View FIGURES 1 – 11 , 22–29 View FIGURES 22 – 37 ). Darker than worker due to blackish metasoma. Face and mesepisterna with abundant plumose setae laterally; labrum slightly convex.
Description. Holotype, worker ( Figs. 1, 5 View FIGURES 1 – 11 ; Tab. I).
Dimensions. Total length, 5.34 mm; forewing length, from apex of costal sclerite to wing tip, 5.19 mm (including tegula, 5.88 mm); maximum head width, 2.12 mm; abdomen width, 2.48 mm.
Integument color. Head and thorax black, contrasting with yellowish abdomen, only TII and apical half of following terga darker yellowish. Antennal scape and mandible yellowish in basal half and ochre-ferruginous in apical half; antennal flagellum fuscous on external surface and bicolored, with basal 2/3 fuscous and apical third reddish on internal surface of each flagellomere; legs ochre-ferruginous, tibia II, femur II and tibia III wholly black except for basal 1/6 and small spot at articulation point with the basitarsus yellow; metepisterna and scutellum ochraceous, last with two brownish medial spots. Wing membrane hyaline, slightly yellowish close to basal veins; alar venation yellowish. Cream-colored maculation occupying lower paraocular area, except for narrow black stripe contiguous with epistomal sulcus, enlarged near antennal socket, with two stripes gradually narrower above, one extending along inner orbit of eye to superior occiput, interrupted at tangent of upper interorbital, other slightly surpassing antennal socket separated by U-shaped black area. Other cream-colored areas include almost whole clypeus except for light brownish irregular medial spot (partially covered by pollen—in paratypes, two bars converging above); one trapezoid in supraclypeal area; almost whole pronotum, except one brownish spot next to medial posterior margin and blackish corners; anterior 1/2 of pronotal lobes; very narrow stripe (about 1/3 of scape diameter) stripe following posterior paraocular margin; posterior 1/2 of tegulae; one stripe with 1/2 to 1/3 diameter of scape at each side of mesoscutum, next to lateral margins and axillae almost completely cream colored.
Pilosity. Well developed and predominantly ochraceous except pale whitish on face and ventrally on thorax and abdomen, and in anterior and black posterior margins of tibia III; mainly with simple setae except shortly plumose on face, sides of thorax (pronotal lobes, anterior corners of mesoscutum, mesepisternum, epimeron, metepisternum and near propodeal spiracles), and apical margin of tibia III, where setae are shortly plumose; external margin of tibia I covered with long, dense, spatulate setae; compound eyes with short pilosity ( 35–40 mm long) well evident; remarkably long and scattered on abdominal terga mainly TV–TVII, more concentrated laterally.
Integument. Smooth and shiny, only with very small sparse piligerous punctures, the space between punctures larger than their diameter.
Form and proportions (measurements in Table I). General shape of the body as described for genus. Head slightly wider than long (2.12: 1.78). Eyes 3x longer than wider (1.32: 0.42) converging slightly below and as long as lower interorbital distance; upper interorbital distance, slightly smaller than the interorbital maximum (1.36: 1.48: 1.32); malar area short, almost linear; clypeus 0.56x shorter than its maximum width (0.68: 1.22) and 0.44x clypeocellar distance (1.56); mandibles as described for the genus; interalveolar distance (between antennal sockets) slightly larger than alveolar diameter (0.26: 0.22), alveolorbital ca. 1.46x first and alveolocellar distance slightly longer than 3x (0.26: 0.22: 0.38: 0.72); interocellar distance 2.8x ocellar diameter (0.50: 0.18) and ocellorbital 1.7x (0.30: 0.18); antennal scape long, surpassing the vertex and slightly shorter than 1/2 length of flagellum plus pedicel (0.90: 2.08). Flagellomeres as described for genus. Tibia III club-shaped, triangular with posterior angle largely rounded, 2.5x longer than wide; basitarsus III with parallel sides and posterior angle obtuse, 2.8x longer than wide (0.90: 0.32) and as long as the maximum width of tibia III. Abdomen approximately ogived, a little wider than thorax (2.48: 2.36) on the base and pointed at the apex.
Male ( Figs. 3, 6 View FIGURES 1 – 11 ; Tab. I). Dimensions. Total length, 5.19 mm; forewing length, from apex of costal sclerite to wing tip, 5.27 mm (including tegula, 5.90 mm); maximum head width, 2.20 mm; abdomen width, 2.55 mm.
Integument color. Like worker except scutellum and legs darker and yellow markings on face and thorax less vivid.
Pilosity. Plumose on face and sides of mesepisterna, as in worker, only slightly longer and denser; paler on thorax, mainly scutellum. Outer surface of tibia I with simple setae, not spatulate, as in male of T. extranea . Alar microtrichiae shorter and less dense than in worker, anterior wings fuscous on apical half and pale yellowish on base, not as yellow as worker. On abdominal terga, finer, sparser and less abundant. Posterodistal 1/3 external surface on tibia III with simple setae, denser, longer and finer, with intermixed shorter plumose setae.
Integument. Smooth and shiny, as in worker.
Form and proportions (measurements in Table I). General shape of the body as described for worker. Head 1.28x wider than long (2.20: 1.72) and 1.45x wider than clypeocellar distance (1.52). Eyes 2.09x longer than wide (1.38: 0.66) converging below and 1.2x longer than lower interorbital distance (1.14); upper interorbital distance slightly smaller than interorbital maximum (1.28: 1.34: 1.14); malar area short, almost linear; clypeus 0.61x shorter than maximum width (0.67: 1.10) and 0.44x clypeocellar distance (1.52); mandible long and slender, slightly longer than half clypeocellar distance (0.88: 1.52), with small indentation near inner angle, not forming a proper denticle; labrum normal, slightly convex; interocellar distance 1.92x longer than ocellorbital and ocellus diameter (0.46: 0.24: 0.24); antennal scape shorter than in worker, almost as long as alveolocellar distance (0.65: 0.68) and ca. 1/4 length flagellum plus pedicel (0.65: 2.63); first flagellomere short, 1.75x as long as wide; interocellar distance 1.92x ocellus diameter. Scutellum 1.96x wider than long. General shape of tibia III as in worker, 2.54x longer than wide, with the posterodistal quarter slightly concave (a little less than in T. extranea ), similar to shallow corbicula, swollen in anterior 1/3; basitarsus III rectangular as in worker, slightly shorter and wider, ca. 2x longer than wide. Shape of abdomen similar to worker. Genitalia and pregenital sterna as in the figures 22–29.
Type material. Holotype, worker, from "Candeias do Jamari - RO ca. 3,5 km W, margem esquerda rio Candeias", " BRASIL 17.IV.2011 08°47'S, 63°44'W S. Pedro & G. Cordeiro leg,", "905c-006 RPSP ninho Coleção Camargo", 12 paratypes—two males (one pupa) and 10 workers (1 fixed in anhydrous alcohol)—with same labels, all in RPSP, and 9 paratypes (5 dried from alcohol, and 4 kept fixed in alcohol) from the same nest with the date " 09.IX.2011 " and "G.D.Cordeiro leg. (N19)", three donated to AMNH (905c-005), SEMC (905c-003), and DZUP (905c-002), the other six in RPSP.
Etymology. The species here described is named honoring the late Prof. Dr. João Maria Franco de Camargo ( 1941-2009), who discovered Trichotrigona and described the first species, in addition to two other genera, 88 new stingless bee species and one Xylocopa Latreille, 1802 (Apidae) . His contributions to the taxonomy, biogeography and biology of the stingless bees ( Pedro, 2009) have substantially advanced the knowledge of this tropical, social and honey making bee group.
Geographic distribution and habitat. Trichotrigona camargoiana sp. nov. is known only from the type locality, Candeias do Jamari, Rondônia, Brazil ( Fig. 47 View FIGURE 47 ), between the right margin of Madeira and left margin of the Candeias River, in a region with strong anthropogenic influence (buildings, roads, crops and just 15 km E of the capital, Porto Velho), surrounded by patches of restored forest about 10 meters high. A few kilometers from the meliponary, lowland native forest still covers the banks of the Candeias (about 19 km W) and Madeira rivers (about 0 2 km E).
The meliponary, where the nest of T. camargoiana sp. nov. was found, was installed in a large clearing (ca. 700 m ²) of upland ("terra-firme", not subject to periodical inundation) close to other clearings surrounded by regenerating forest. The closest lowlands, preserving the native vegetation and subject to periodical flooding, were about 1 km NE from the margins of a tributary of the Candeias River.
TABLE I. Measurements (mm) of Trichotrigona extranea (Daraá river, Amazonas, Brazil) and T. camargoiana sp. nov. (Candeias do Jamari, Rondônia, Brazil).* measurements of the holotype of T. extranea were retaken (using the same procedures and same stereomicroscope as used for T. camargoiana ) in order to be directly comparable with the ones of T. camargoiana sp. nov. ** from Camargo & Pedro 2007.
Bionomics and nesting. The nest was established on August 26th, 2010, in a small wooden hive of about 20 x 10 x 10 cm, in a meliponary that housed mainly nests of Melipona ( Michmelia) seminigra abunensis Cockerell, 1912 and Frieseomelitta trichocerata Moure, 1990 , besides species of Plebeia Schwarz, 1938 , Scaptotrigona Moure, 1942 , Nannotrigona melanocera ( Schwarz, 1938) , and other stingless bees. According to information from the beekeeper, the hive was previously occupied by other Meliponini but was empty when the new nest was established. Orchids ( Orchidaceae ), avocado ( Persea americana Mill. , Lauraceae ), “Urucum” or Achiote ( Bixa orellana L., Bixaceae ), Byrsonima (Malpighiaceae) and several species of flowering plants were cultivated nearby.
Nest characteristics. The description presented here is based on few data, obtained during four days visiting the meliponary. The intention was to keep the nest preserved for future studies obtain detailed information on food gathering behavior, and to collect more specimens, including the queen. Unfortunately, the colony died a few months later and no additional studies were performed. No pieces of the nest or specimens were preserved.
The nest entrance was constituted only by a small orifice of ca. 3.5–4.0 cm in diameter, without a projecting tube, just a short ring of sticky, dark reddish resin ( Fig. 38 View FIGURES 38 – 46 , detail). Dark resin was spread mainly above and lateral to the ring, in an area of about 3–6 mm long ( Fig. 38 View FIGURES 38 – 46 ), like a frame. The cavity, inside the wooden box, had a thin protective layer of cerumen, probably, but it is difficult to say if it was deposited by T. camargoiana sp. nov. or a previous occupant.
A large deposit of dark reddish and yellowish resin was found adjacent to the wall containing the nest entrance ( Fig. 42 View FIGURES 38 – 46 ). This mass of sticky (more recently deposited) and solid (older) resin was crossed by a gallery connecting the exterior of the nest with the brood chamber inside the box. Deposits like this were found also in the nests of T. extranea ( Camargo & Pedro 2007a). One male was photographed carrying resin on tibia III ( Fig. 42 View FIGURES 38 – 46 , detail).
The brood chamber was composed of a cluster with about 350 cells and cocoons; the older cocoons (with defecating and postdefecating larvae or pupae) were attached by short connectives of cerumen to the top of the box and continued joined after the box had been opened ( Fig. 39, 43 View FIGURES 38 – 46 ). The cluster of cells and cocoons had no involucrum surrounding it, as in T. extranea nests ( Camargo & Pedro 2007a). The brood cells were arranged in irregular clusters, although some cells seemed to be aligned to compose a comb ( Figs. 39, 43 View FIGURES 38 – 46 ). The same pattern was found in one of the three nests of T. extranea (807c, Camargo & Pedro 2007a) and is probably related to the shape and diameter of the cavity available for the brood chamber. Long pillars and connectives were absent; only short connectives were found joining cells to the hive walls ( Figs. 39, 41, 43 View FIGURES 38 – 46 ).
The cylindrical cocoons, made with drier dark yellowish cerumen (darker at bottom), were about 5.8 mm high by 3.78 mm wide, while the cells, lighter and smoother, were a little more spherical ( Figs. 39, 40, 41, 43, 44 View FIGURES 38 – 46 ). Two cells were under construction ( Fig. 44 View FIGURES 38 – 46 ) and still without provisions. Open cells displayed liquid provisions and with eggs positioned vertically ( Fig. 40 View FIGURES 38 – 46 ).
Storage pots were not identified. Considering the uniformity, disposition and shape of the brood cells, there was no evidence of pollen or nectar pots.
Nest population. One physiogastric queen was observed ( Fig. 31 View FIGURES 22 – 37 ), however the stage of damage to the forewings, in order to estimate queen age, could not be determined. Adult males also were seen inside the nest ( Fig. 42 View FIGURES 38 – 46 , detail). The nest population could be subjectively estimated at ca. 200 adult bees, following the calculation proposed by Roubik (1979) which considers the number of cells and cocoons (total brood), a field estimate of total adult bees and the proportions of brood and adult bees found by Camargo & Pedro (2007a) for the older nests of T. extranea (806c and 807c). The larva of T. camargoiana sp. nov. has high and well evident dorsolateral tubercles ( Figs. 45, 46 View FIGURES 38 – 46 ), much more prominent than in larvae of Partamona Schwarz, 1939 , as described by Michener (1953).
Behavior. The workers of T. camargoiana exhibit a very docile behavior. Even when the cover of the wooden hive box was removed the workers exhibited no aggressive behavior. A guard was always present and occupied almost the whole entrance orifice, most of the time with the head upside down ( Fig. 38 View FIGURES 38 – 46 , detail). She stood back only when a forager was arriving or when a bee was departing. The aperture is sized just for one bee and is closed with cerumen in the evening, then being re-opened at daylight. Apparently the same guard was seen ejecting a load of waste with its mandibles.
The flight of foragers was slow, and despite this it was not possible to follow them for long. Only four or five foragers left the nest per hour during the morning. Some collecting with an entomological net was carried out around the meliponary, but no T. camargoiana sp. nov. was caught or seen visiting any of the nearby plants. The bees were not found collecting resin on nearby tree trunks. The beekeeper said that he has never seen the bees on flowers. Of the bees collected, none of them carried pollen loads on their corbiculae or even on the mesepisternal setae. One of them had a mass of pollen between the mandibles and below the labrum. The dissection of another worker revealed no pollen carried internally in the crop. Odor baits (cineole, vanillin, benzyl acetate, methyl salicilate, eugenol and benzyl benzoate) were hung in the vicinity but none were visited by T. camargoiana sp. nov.
The nest entrances of Frieseomelitta trichocerata were observed but no T. camargoiana sp. nov. was seen attempting to enter the nests or even near them.
Larval provisions collected from one cell had the following pollen types, in decreasing order by frequency: Didymopanax (Araliaceae) , Mimosa (Mimosoideae) , Arecaceae (Palmae) , Hyptis (Labiatae) and Anacardiaceae . In less quantity, there also were Poaceae (Gramineae) , Alchornea / Aparisthmium (Euphorbiaceae) , Urticaceae , Cyperaceae , Asteraceae , Melastomataceae and Myrtaceae .
Remarks. Considering the morphological similarity as well as the nest characteristics of Trichotrigona camargoiana sp. nov. and T. extranea , it is reasonable to infer they share the same food gathering behavior. Perhaps, a meaningful difference is presence of plumose setae on the face and mesepisterna of T. camargoiana sp. nov., considering that bees with parasitic or cleptobiotic behavior generally have no or few plumose setae (e.g., Lestrimelitta ). Nonetheless, both species present the same level of reduction in pollen-collecting apparatus, added to the facts that storage pots were not found in the nest of T. camargoiana sp. nov. and none of the bees collected had loads of pollen on the corbiculae or pollen grains on the mesepisternal plumose setae. One of the workers had a mass of pollen in the hypostomal region between the mandibles, which may imply that they can carry pollen in the crop and use it directly for cell provisioning. There is apparently no pollen storage within nests of this genus. The suspicion that Trichotrigona may steal provisions from nests of Frieseomelitta remains without strong support, except for the fact that all nests of Trichotrigona were found very close to nests of Frieseomelitta . Three nests of T. extranea occupied the same tree cavity as nests of Frieseomelitta [ F. paupera ( Provancher, 1888) and Frieseomelitta sp., Camargo & Pedro 2007a] but with no direct connection between them, being separated by a compact batumen ( Camargo & Pedro 2007a). The nest of T. camargoiana was less than seven meters away from 10-15 colonies of Frieseomelitta trichocerata . It is interesting to note that both genera are phylogenetically closely related ( Camargo & Pedro 2003; Rasmussen & Cameron 2010).
Another intriguing question is how the workers use the specialized setae on tibia I. One hypothesis is that this brush is used to handle resin. The workers probably carry resin on the corbicula, the same way males do using the external surface of tibia III ( Fig. 42 View FIGURES 38 – 46 , detail), and the specialized setae could be a tool to aid removal from hard surfaces, but no evidence of resin was found on those setae. Males do not have the spatulate setae on tibia I, but carry resin on tibia III. Their use on manipulation of resin remains to be investigated. On the other hand, the hairs are always shinning, reflecting light as if they were lubricated with oil. But if these bees collect oils, in the same manner as Centridini (Apidae) , then how they would be using this oil? Another interesting fact is that the inner side of tibia I lack those specialized setae (generally slightly spatulate, broadened near the apex or chiseled) possibly related to some grooming behavior (see Jander 1976), present in most, if not in all, of the bees. Could the long and narrow basitarsus II be filling some role of the tibia I? With some more detailed observation of bee behavior it will be possible to figure out how these legs work.
The observations presented here were made during a very short period of time (5 days), and the goal was to keep the colony alive for future studies. Unfortunately, the colony died prematurely, and it was not possible to more fully elaborate on the life history of this intriguing bee. Until other nests of Trichotrigona camargoiana sp. nov. or T. extranea are discovered and detailed studies on their behavior are carried out, many questions remain about these enigmatic bees.




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Tribe |
Meliponini |
|
Genus |