
Melitaea shahvarica, Kolesnichenko & Kotlobay, 2022
|
publication ID |
https://doi.org/10.5852/ejt.2022.830.1865 |
|
publication LSID |
lsid:zoobank.org:pub:C1F47AD9-ECF6-4F0F-9928-55A45332FF4B |
|
DOI |
https://doi.org/10.5281/zenodo.6855862 |
|
persistent identifier |
https://treatment.plazi.org/id/295892C7-2C50-471F-A36B-CE1F69FA8E8C |
|
taxon LSID |
lsid:zoobank.org:act:295892C7-2C50-471F-A36B-CE1F69FA8E8C |
|
treatment provided by |
Felipe |
|
scientific name |
Melitaea shahvarica |
| status |
sp. nov. |
Melitaea shahvarica sp. nov.
urn:lsid:zoobank.org:act:
Figs 4A – D View Fig , 14 View Fig , 16A – C View Fig , 18A – C View Fig , 21–28 View Fig View Fig View Fig View Fig View Fig View Fig View Fig View Fig , 29A View Fig , 30 View Fig ; Tables 4–5 View Table 4 View Table 5
Differential diagnosis
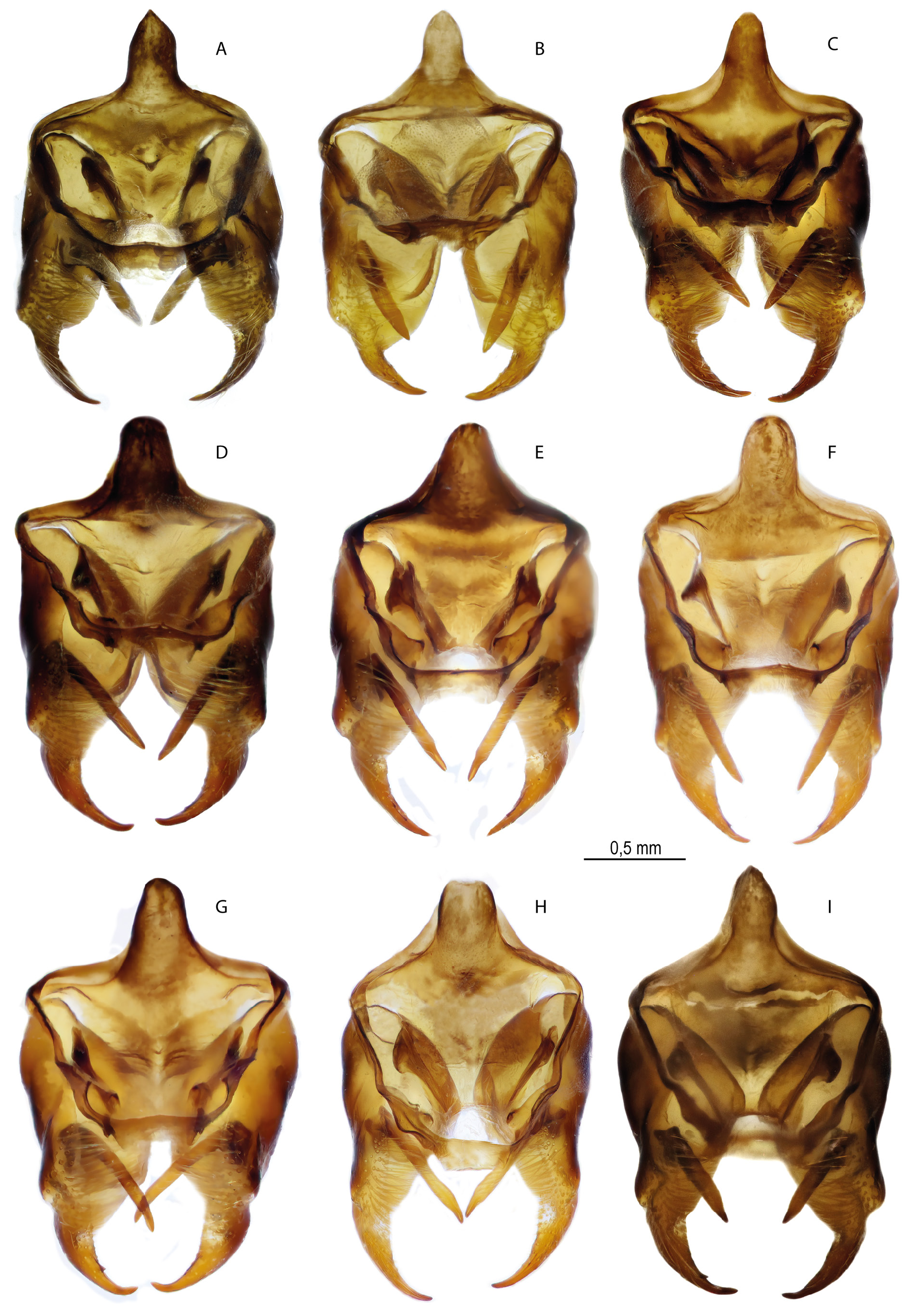
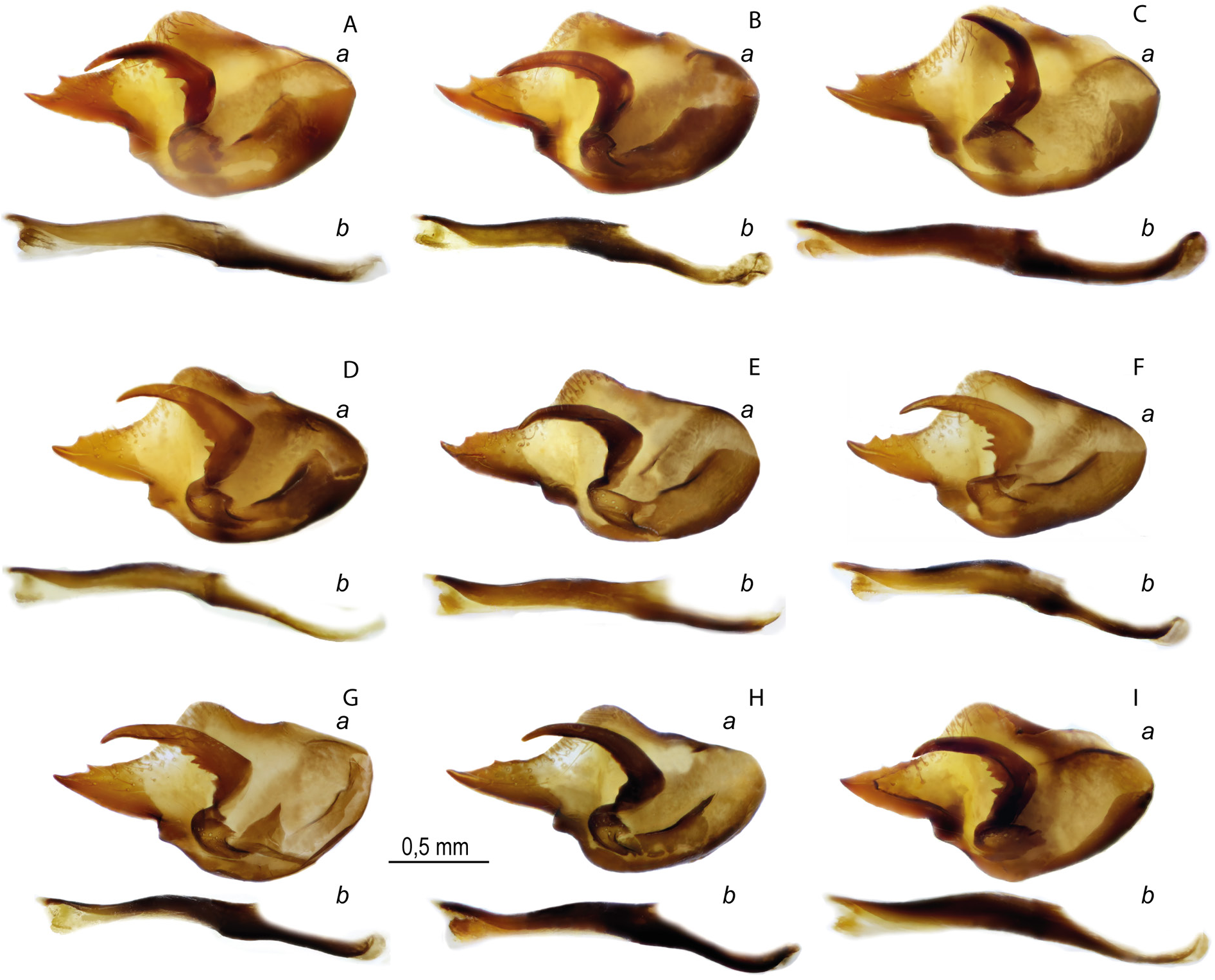
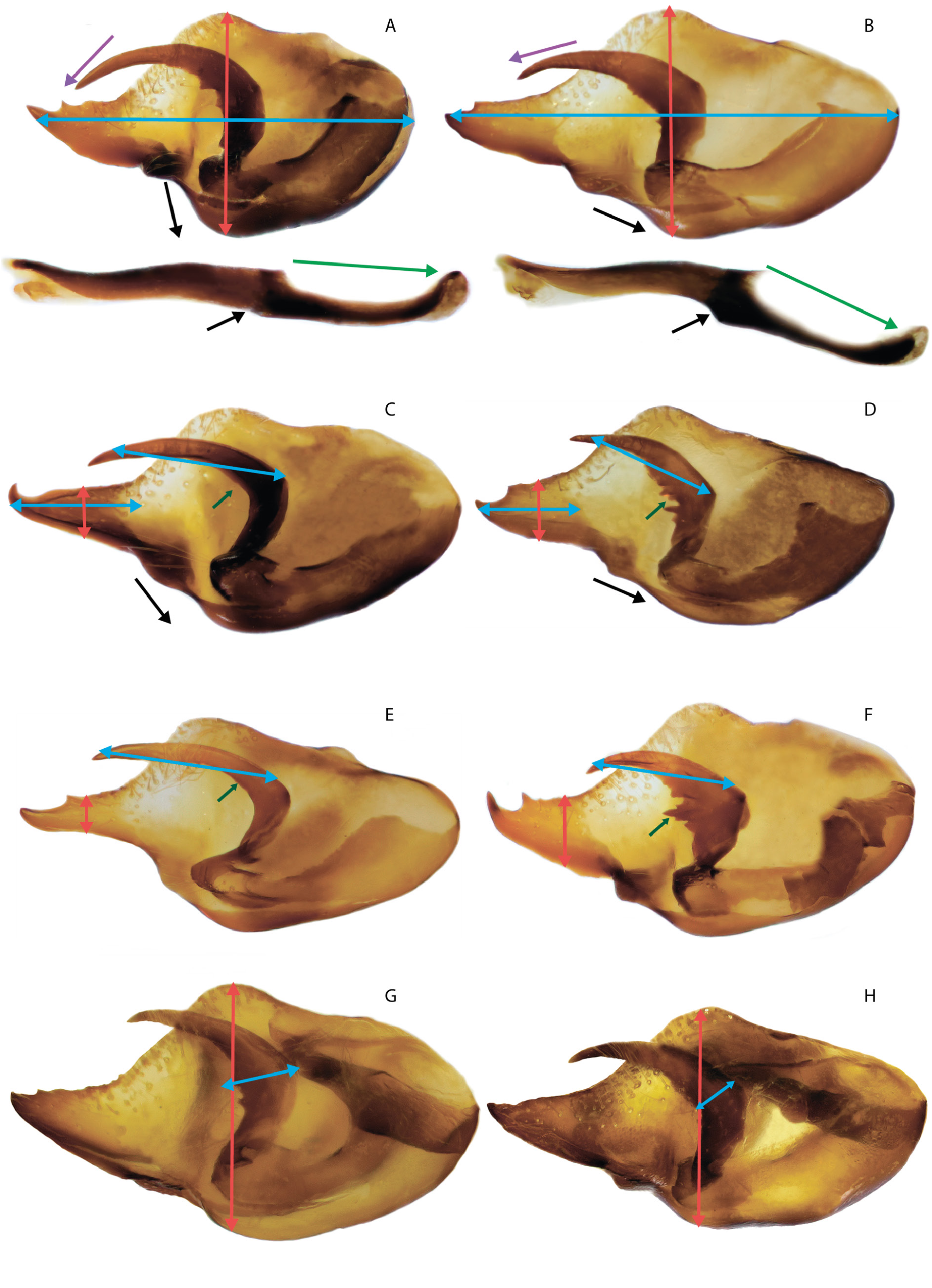
Melitaea shahvarica sp. nov. differs well from the close species M. timandra in its smaller size (small specimens of M. timandra binaludica subsp. nov. have a wingspan of more than 16.5 mm, while the wingspan of M. shahvarica is on average 16.5 mm), a noticeable reduction in the black pattern of the UPS (absence of submarginal lunules, weak development of black basal darkening and complete or partial absence UPH discal spots), absence of developed deposition of UNH dark scales, presence of UPF fused costal black dots of the submarginal and postdiscal rows centered by pale-orange spot. For the male genitalia, a characteristic feature is the presence of a noticeable protrusion of the ventral side, due to which the valva is visually noticeably expanded in the central part, so the length of the valva is only 1.5 times as great as the width. In M. timandra , the ventral protrusion of the valva is smoothed and the length of the valva is at least 2 times as great as the width ( Fig. 29 View Fig ). The aedeagus is characterized by the absence of a noticeable protrusion on the ventral side at the junction of the anterior and posterior parts so both parts of the aedeagus are approximately on the same straight line. The female genitalia are characterized by an oval postvaginal plate.
Etymology
The name is toponymic and denotes the name of the mountain on the slopes of which the type series was collected.
Type material
Holotype ( Fig. 21A, I View Fig ) IRAN • ♂; Semnan Prov., Shahrud area, S macroslope of Shahvar Mt. ; alt. 2200–2400 m; 5–6 May 2018; K. Kolesnichenko leg.; SDM .
Paratypes ( Fig. 21B–H, J–P View Fig )
IRAN – Semnan Prov. • 34 ♂♂ (10 dissected), 8 ♀♀ (4 dissected); same collection data as for holotype; EDMSU • 2 ♂♂ (all dissected); same collection data as for holotype but 18–19 May 2017; EDMSU • 16 ♂♂ (10 dissected), 5 ♀♀ (all dissected); Shahrud area, S macroslope of Shahvar Mts, Tohar v. vicinity; 7–8 May 2009; K. Kolesnichenko leg.; EDMSU • 32 ♂♂ (3 dissected), 9 ♀♀ (2 dissected), Gerdab. Elburs Ridge, Shahvar Mt. ; alt. 2200 m; 6–7 May 2018; A.A. Kotlobay leg.; coll. A.A. Kotlobay • 9 ♂♂; same collection data as for preceding but 9 May 2019; coll. A.A. Kotlobay.
Description
Male ( Fig. 21A–D, I–L View Fig )
WINGS. FW length is 16–17 mm, the holotype is 17 mm. UPS ground color is bright orange-red. UPS black pattern reduced; UPS thin black marginal border with well-defined small marginal spots along the outer edge of the wings; UPF submarginal row usually represented by black dots or strokes. UPH submarginal row is absent or reduced. A characteristic feature for most specimens of the new species is the UPF black macule located between veins Sc and R5 and formed by the fused submarginal and postdiscal costal dots centered with orange or pale spot; UPF discal row represented by small black spots not fused with each other, UPH discal row absent or formed by small black spots reduced closer to the costa; UPF postdiscal pale-yellowish area, located behind the black discal spots, is weakly expressed or expressed only at the costal edge: usually, only 2–3 yellowish macules situated near the costa; UPF paleyellowish macule in the distal part of the discoidal cell weakly expressed; UPH basal black suffusion covers less than ¼ of the wing surface; UNF pale area along the outer edge of the wing between the veins Sc and M2 is well developed; UPH discal macules are either absent or expressed only as a row of dots between the veins A2–M3, in rare cases UPH discal row developed and formed by small and delicate macules ( Fig. 21A View Fig ); UNF pale macules in the postdiscal area are not pronounced; white UNH ground color with a slight admixture of dark scales; UNH lunules forming the proximal edge of submarginal orange fascia outwardly concave and sharply pointed between veins M3 and Cu2.
MALE GENITALIA ( FigS 16A – C View Fig , 18A – C View Fig , 22 View Fig , 29A View Fig ). A broad valva with a wide and elongated distal caudal process with 2–3 small spines on the dorsal surface. The relatively short harpe is expanded in the central part with a small number of separately spaced or fused teeth on the inner surface. Curved aedeagus with a rounded convex dorsal edge. There is a smooth bend on the ventral surface of the aedeagus at the junction of the anterior and posterior parts, as a result of which the aedeagus looks S-shaped. Saccus is thin, its length is 2 times as great as its width. In some specimens the saccus is deeply divided into relatively wide rounded lobes.
Female ( Fig. 21E–H, M–P View Fig )
WINGS. FW length is 19–20 mm. UPS ground color is bright orange; UPF postdiscal pale area is more pronounced than in the male; there is a distinct pale macule in the distal part of discoidal cell; UPH submarginal and discal rows are usually reduced, but in some specimens well developed ( Fig. 21E View Fig ); UNH discal row usually represented by 2–4 black spots near the costa; UNS pattern is generally similar to that of the males, but the main color is bright white.
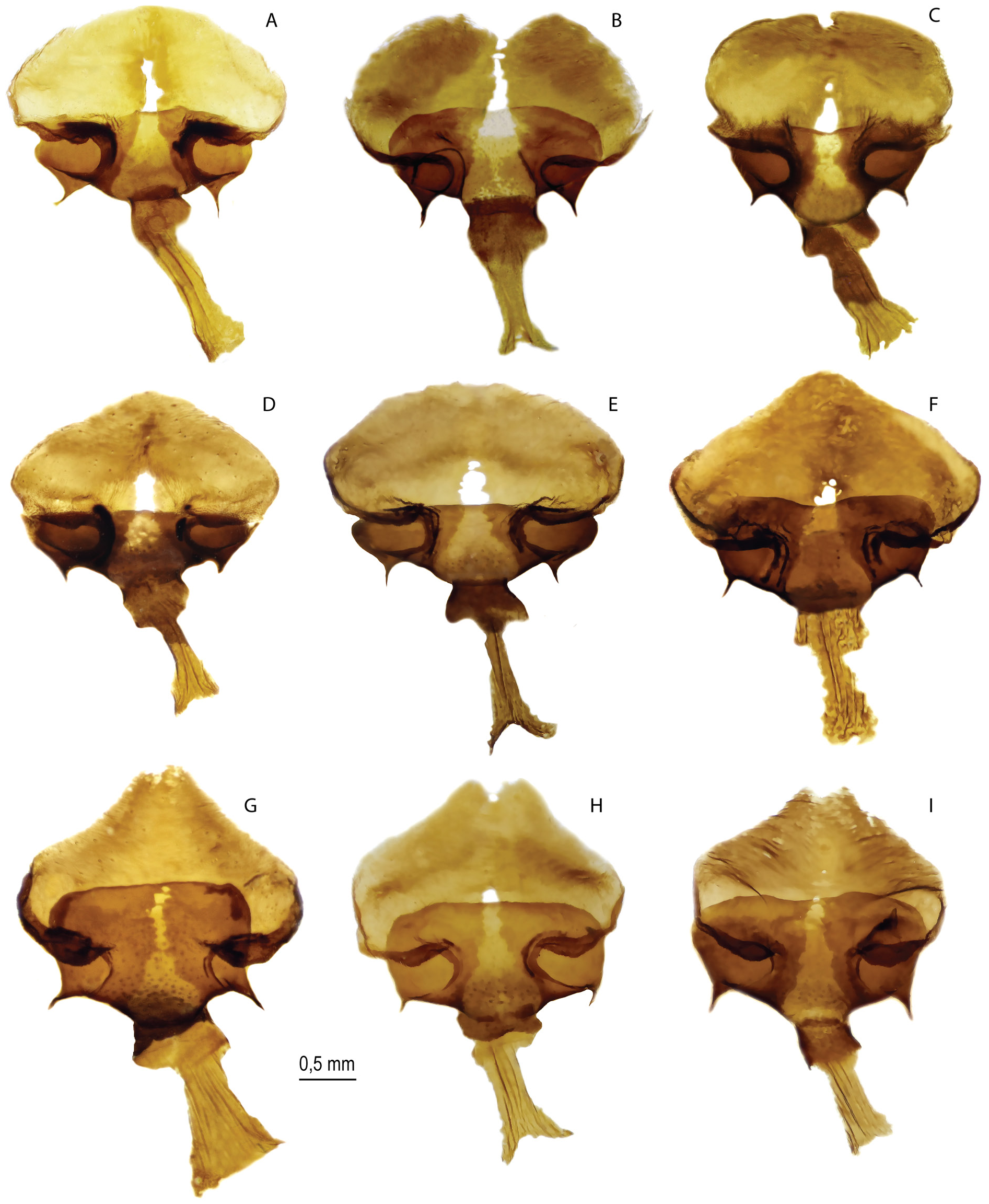
FEMALE GENITALIA ( Fig. 4A–D View Fig ). The postvaginal plate is oval or wide-oval, in rare cases-oval-triangular. The antevaginal plate is narrow in the dorsoventral direction, its outer edge does not extend beyond the boundaries of the bend of the postvaginal plate (auricules).
Preimaginal stages ( Figs 23–26 View Fig View Fig View Fig View Fig , Tables 4–5 View Table 4 View Table 5 )
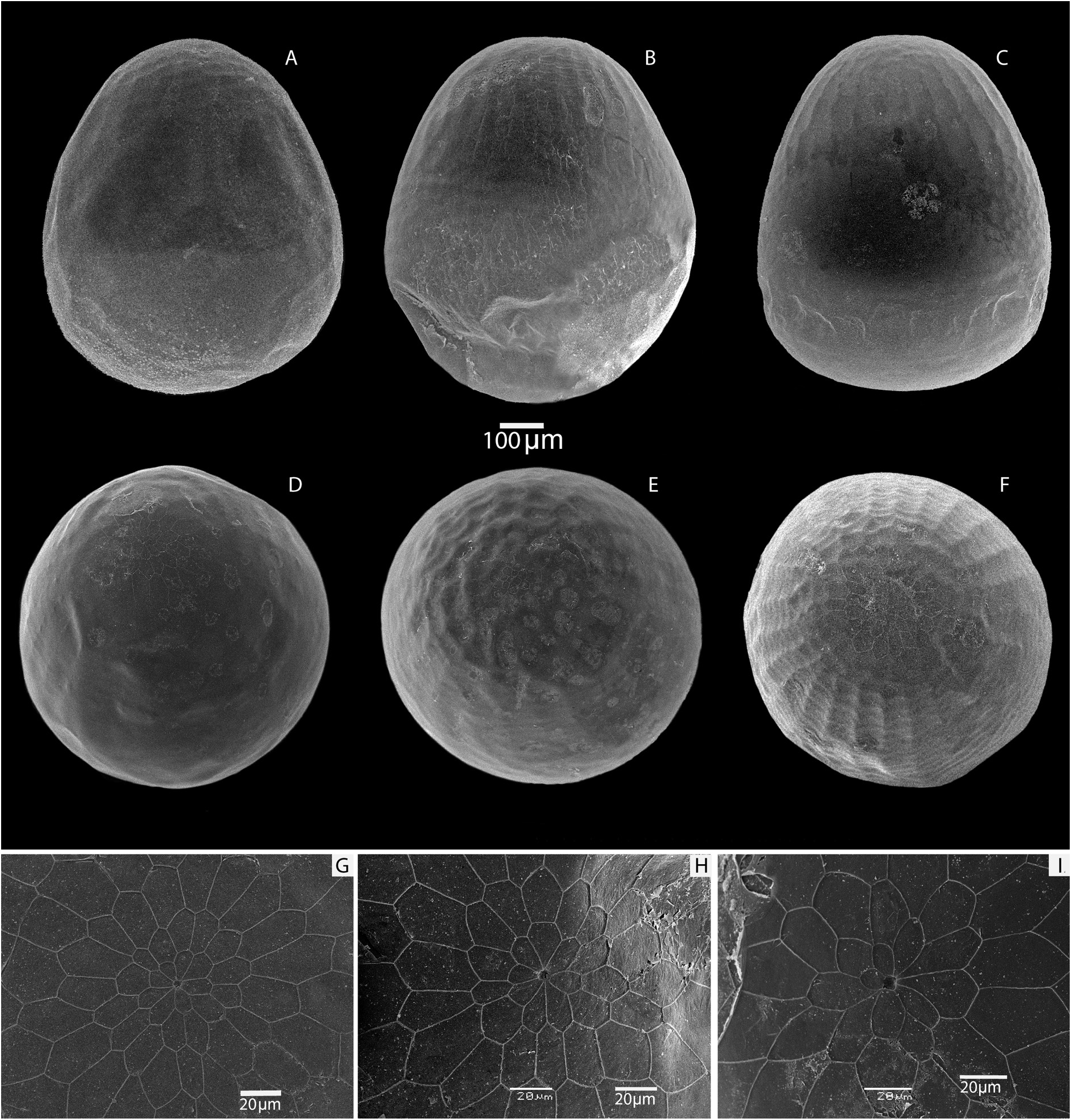
Egg ( Fig. 23 View Fig , Table 4 View Table 4 )
Material examined: 26 eggs from three clutches: Iran, Semnan Prov., Shahrud area, S macroslope of Shahvar Mts, alt. 2200–2400 m.
The egg has a well-defined pear shape. The height of the egg is from 699.4 µm to 700.6 µm, the width is from 606.5 µm to 610.0 µm ( Table 4 View Table 4 ). The sculpture of the micropile region is formed from four to five rows of penta-hexahedral cells of various lengths and widths. The diameter of the micropile rosette in the widest part varies from 41.0 µm to 54.4 µm. The micropile rosette is formed by 7–8 primary quadra-pentahedral cells with width from 7.0 µm to 17.6 µm and length from 10.0 µm to 25.0 µm. The micropile is rounded in shape, with an average diameter of 5.1 µm. There are 24–26 lateral longitudinal ribs in the micropilar region which fall to ⅓ of the egg surface. The transverse ribs are often not or weakly expressed. Below the lateral ribs, the chorion has a cellular structure. The freshly laid eggs are yellow.
In contrast to M. timandra binaludica subsp. nov., whose eggs are oval with a slight narrowing in the apical region ( Kolesnichenko & Kotlobay 2020), the eggs of M. shahvarica sp. nov. are characterized by a pear shape. In addition, the eggs of M. timandra binaludica are on average larger, their maximum height is about 850.0 µm, in M. shahvarica the egg height is on average about 700.0 µm. In M. timandra , the micropile rosette is formed by 9–10 primary cells, and in M. shahvarica , the micropile is surrounded by 7–8 primary cells.
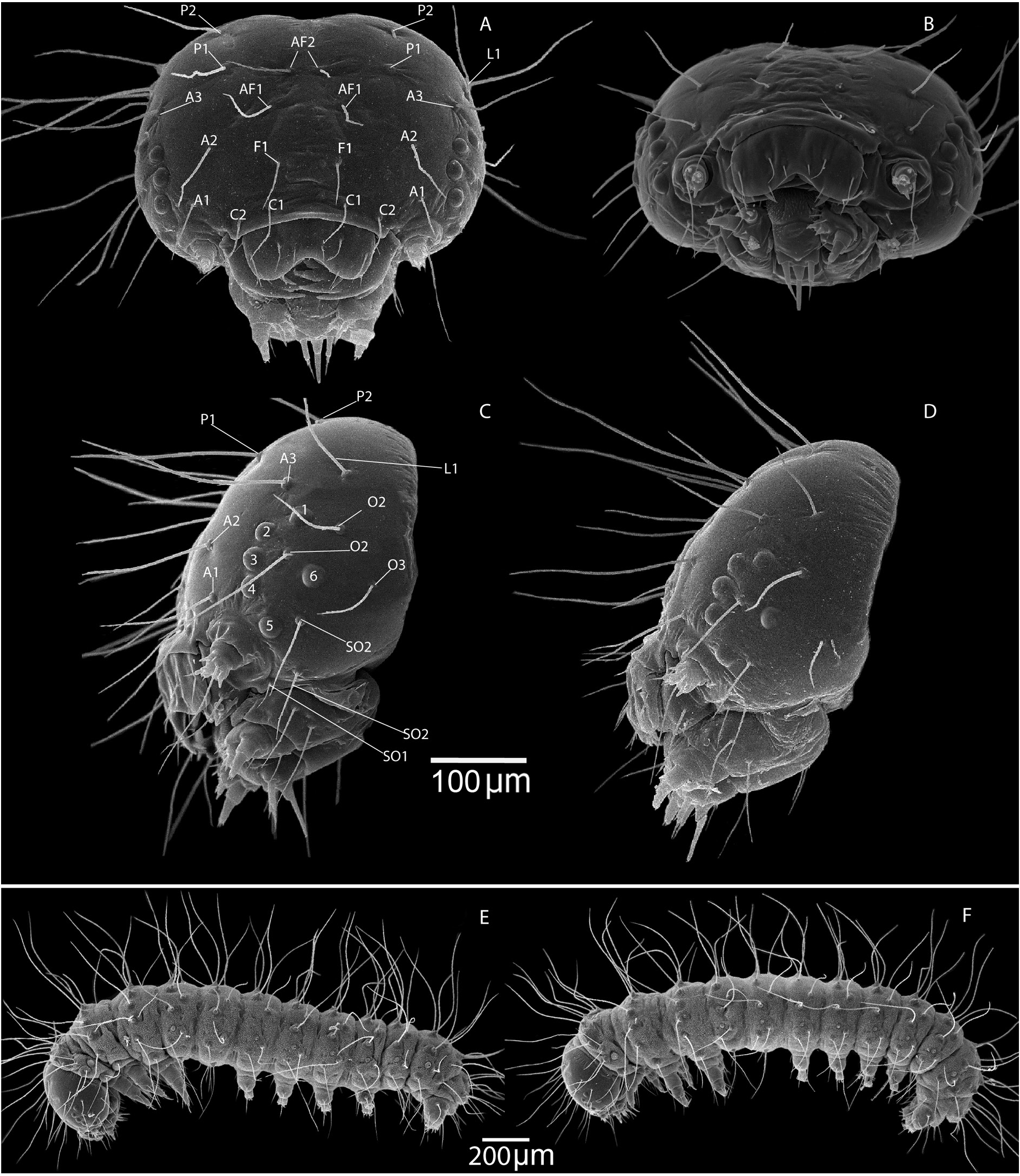
First instar caterpillar ( Figs 24 View Fig , 25A–B View Fig , Table 5 View Table 5 )
The body length of a caterpillar recently hatched from an egg is about 2 mm (1976.4 ± 2.2 µm) ( Figs 24E– F View Fig , 25A View Fig ). The body length of a caterpillar of the first instar before molting is about 3 mm ( Fig. 25B View Fig ). A caterpillar hatched from an egg is light green with a black head, covered with long hairs. The width of the head capsule is 412.5 ± 0.2 µm, the height is 322.5 ± 0.7 µm ( Fig. 24A View Fig ). From the side, the head capsule is triangular, expanding to the area of the oral organs ( Fig. 24C–D View Fig ). The distance in the widest part of this triangle between the labrum and the base of the head is 240.0± 1.4 µm. The eyes are approximately the same size, their location is similar to that of M. timandra binaludica subsp. nov. ( Kolesnichenko & Kotlobay 2020). The labrum with a width of 134.4± 0.4 µm is divided into 2 blades. The arrangement of the bristles on the head capsule is conservative. The distance between the bristles is shown in Table 5 View Table 5 .
We compared the distances between the bristles located on the lateral part of the head capsule of the first-instar caterpillars M. shahvarica sp. nov. ( Table 5 View Table 5 ) and M. timandra binaludica subsp. nov. ( Table 6). The location of the bristles and the distance between them on the head capsule are relatively stable for each species. Differences in the distance between the bristles in M. timandra binaludica and M. shahvarica are observed between L1–O1 (about 111.0 µm for M. shahvarica and about 113.0 µm for M. timandra binaludica ) and L1–O3 (about 122.0 µm for M. shahvarica and about 127.0 µm for M. timandra binaludica ).
Second instar caterpillar ( Fig. 25C View Fig )
The body length is about 3.5 mm. The main color is brown, the head and the scoli are black, the bases of the scoli are flesh-colored. The main lines of the body are expressed due to a darker color.
Third instar caterpillar ( Fig. 25D View Fig )
The body length is about 4.5–5 mm. The main color is brown with flesh-colored specks. The head, the scoli and the tops of the legs are black. The black outgrowths of the linea dorsalis are located on a brown background and are present only on the abdominal segments. The bases of the scoli of the subdorsal stripe are bright yellow. The bases of the scoli of the dorsal and other stripes are flesh-colored.
Forth instar caterpillar ( Fig. 25E View Fig )
The body length is about 9–12 mm. The head capsule is black. The body pattern is formed by a black main background with light orange or flesh-colored small specks. The scoli are light orange or flesh-colored. The segments of the thoracic part are characterized by the absence of yellow or flesh-colored scoli on the unpaired linea dorsalis. The paired linea subdorsalis is marked with flesh-colored scoli at their base. The linea epistigmalis paired stripe above the spiracles is black. Bright yellow scoli are located on each segment. The stigma pair (linea stigmata) is of the base color as well. The linea hypostigmalis is black with flesh-colored scoli located on each segment. Linea basalis and linea subbasalis are black with a flesh-colored scoli on each segment.
Fifth instar caterpillar ( Fig. 26A View Fig )
The body length is 15–17 mm. The head capsule is black. The body pattern is formed by bright yellow or flesh-colored scoli and a background color that varies from dark brown to black with pronounced fleshcolored specks. Unpaired linea dorsalis is marked with several bright yellow scoli with a dark (black or dark brown) base. However, there are no such outgrowths on the thoracic segments. The subdorsal (linea subdorsalis) and suprastigmal (linea epistigmalis) paired stripes located on the dorsal side and above the spiracles have a complete set of bright yellow scoli located on each segment of the body, with the exception of XII. Moreover, the scoli marking linea subdorsalis have a bright yellow base. The paired stigmata stripe (linea stigmata) has a background color. On each segment of the body (with the exception of the middle-thoracic, posterior-thoracic and last segments of the abdomen) in the middle of this stripe, under the base of the outgrowths, there are black spiracles bordered with yellow. The poststigmal line (linea hypostigmalis) is marked with a bright yellow scoli. Linea basalis and linea subbasalis (at the very bases of the legs) are located close to each other so that they practically form a single stripe with bright yellow scoli placed near each other. Poststigmal as well as basal and subbasal outgrowths are absent on the last segment of the body. The legs of the thoracic segments are black, the false legs are brown, with a flesh-colored border at the base.
It should be noted that the color described above is characteristic of some of the caterpillars that have passed into the fifth instar. For some of the caterpillars of the fifth instar, the black color of the main background and the presence of black scoli are characteristic, with the exception of the paired linea subdorsalis, whose scoli are painted in bright yellow.
Sixth instar caterpillar ( Fig. 26B–C View Fig )
The body length is 19–21 mm. The head capsule is black, 2.8 mm wide and 2–2.3 mm high ( Fig. 26C View Fig ), covered with black hairs. The area of simple eyes is black. The eyes are approximately the same size, four of them are located closer to the epicranial suture, and two are closer to the occipital region. The forehead, the platypus, the frontal sclerites of the frontal sutures and the area of the epicranial suture are painted black. The body pattern (the 2 nd and 3 rd thoracic and all abdominal segments) is formed due to bright yellow scoli and the background black color. The unpaired linea dorsalis bears black scoli, there are no such outgrowths on the thoracic segments. The paired linea subdorsalis is marked with bright yellow scoli with a bright yellow base. The paired linea epistigmalis located above the spiracles has a complete set of black scoli located on each segment of the body, with the exception of XII. On the last segment, only the black dorsal and bright yellow subdorsal outgrowths are well developed. The paired linea stigmata has a background color. On each segment of the body (except for the middlethoracic, posterior-thoracic and last segments of the abdomen), black spiracles are located in the middle of this stripe under the base of the outgrowths. The linea hypostigmalis is formed by black scoli. The linea basalis and subbasalis (at the very bases of the legs) are located close to each other so that they practically form a single stripe with black scoli placed near each other. Poststigmal, as well as basal and subbasal outgrowths are absent on the last segment of the body. The false legs and legs of the thoracic segments are black.
The described pattern refers to the caterpillars of the sixth instar, passing into the estivation mode.
Biology ( Figs 27–28 View Fig View Fig )
Observations in nature on the behavior and biology of M. shahvarica sp. nov. were carried out in May 2018 and 2019 and in the first half of July 2019 in the vicinity of the village of Gerdab on the southern macroslope of the Shahvar Mountain on the Elburs Ridge ( 36°31′ N, 54°43′ E). Observations on the development of eggs and caterpillars of M. shahvarica sp. nov. were performed from May to September 2018 and from May 2019 to April 2020 under laboratory conditions in Moscow.
Within the surveyed territory ( Fig. 27A View Fig ), M. shahvarica sp. nov. forms small but very dense local populations, strictly tied to the places of growth of the host plant. We found two populations at an altitude of 2150–2200 m and one at an altitude of 2500 m above sea level, separated by 2–3 km from each other. The flight of M. shahvarica imago at an altitude of 2150–2200 m begins, depending on weather conditions, at the beginning of the first or second decade of May and continues throughout the month. Melitaea shahvarica is the earliest species of fritillaries and one of the earliest, in terms of departure, species of butterflies on Shahvar Mt. At an altitude of 2500 m the flight starts 2 weeks later than at an altitude 2150–2200 m. The males fly out first, 5–7 days earlier than the females. Butterflies fly in the morning in sunny weather. Males patrol the bottoms of inter-ridge valleys in search of females. The butterfly rises to the top of the valley, passes over the ridge and descends across the neighboring valley, repeating the cycles of ascents and descents many times. At the most intensive flight period, up to 30 males flying in one direction are recorded in one place per hour. Fresh-born unfertilized females stay in the bottoms of valleys, sit on plants or make short flights. Fertilized females climb the slopes of the ridges to the places where the host plants grow.
The host plant of M. shahvarica sp. nov. is Phlomoides molucelloides (Bunge) Salmaki ( Fig. 27C View Fig ). Phlomoides molucelloides is occasionally found in dense beds consisting of several plants on crushed stone dumps along the banks of dry riverbeds at an altitude of 2000 m above sea level in a wide intermountain valley at the foot of the Shahvar mountain. But neither the imago nor the caterpillars of M. shahvarica was found at this station. At an altitude of 2150–2500 m above sea level, Ph. molucelloides occurs outlying spurs of the southern macroslope of Mount Shahvar on the upper third of the slopes of the western, northwestern and northern exposure, as well as on the tops of the ridges. It should be noted that the form of Ph. molucelloides growing in the described conditions is characterized by its miniature size. The height of the peduncle is usually only 5–10 cm, and even in the largest specimens does not exceed 20 cm. The growth station of Ph. molucelloides at an altitude of 2150–2500 m above sea level consists of steep rubble-clay scree and clay-rubble areas occupied by sparse friganoid communities of acantholimons ( Acantholimon spp. ), esparcetes ( Onobrychis spp. ), astragalus ( Astragalus spp.) and cushion-shaped sagebrush ( Artemisia spp. ) with the latter dominating ( Fig. 27B View Fig ). Phlomoides molucelloides occurs in single plants or in small groups of 2– 3 specimens almost across the specified territory, but sometimes forms very sparse (the distance between individual plants is from several to one and a half dozen meters) associations consisting of dozens to hundreds of plants and occupying a significant area. It is in such places that M. shahvarica lays eggs and caterpillars develop.
The fertilized female, sitting on the edge of the leaf of the host plant, feels the soil under the leaf rosette with the ovipositor for several minutes, choosing the optimal characteristics of the substrate for laying ( Fig. 27D View Fig ). Fourteen fresh clutches of M. shahvarica sp. nov. were found during the observation of egg-laying females and the examination of 38 plants of Ph. molucelloides . The size of clutches ranged from 24 to 146 eggs. The clutches were placed directly on the ground or rubble under the leaves of the basal rosette ( Fig. 28A–B View Fig ). All the clutches were found under host plants that had not yet begun to develop a peduncle. A total of 79 eggs were collected in nature in order to further monitor the development of the preimaginal stages of M. shahvarica under laboratory conditions. The female, placed in a small plastic cage with a host plant, laid 162 eggs in three batches, with an interval of 1–2 days. Freshly laid eggs are light yellow greenish in color, less than 1 mm in diameter. The hatching of the first caterpillars from the eggs begins after 3 days, and the mass release occurs on 7–9 days after egg laying. Of the 241 eggs (79 collected in nature and 162 obtained from the female in the laboratory) available to us, 233 caterpillars were born (96.7% yield). The day before the release, the part of the egg, where the head capsule of the caterpillar is located, darkens. Fresh-born caterpillars are about 1 mm long, light yellow-greenish in color, covered with long light hairs. The head capsule is dark brown. Immediately after emerging, the caterpillars partially eat the chorion and fall into a stupor for several hours. After coming out of the stupor, they gather in groups on the upper side of the leaf of the host plant ( Fig. 28E View Fig ). First, they scrape the surface, then gnaw through the leaf, leaving the veins intact. After 2–4 days after leaving the eggs, the caterpillars molt and pass into the second instar. Immediately after molting, the head capsule is pale green, after a few hours it turns black. The surface of the body of caterpillars of the second instar darkens, but instead of hairs, outgrowths of a light green color appear with a bunch of spines on top. The caterpillars actively feed and keep together in communities. In the second instar, the caterpillars spend 3–4 days and molt into the third instar. Caterpillars of the third instar are about 1 cm long, black, the head capsule is black. Dorsal and lateral outgrowths on the body are light yellow with a bunch of black spines on the top ( Fig. 28C View Fig ). The III instar lasts 5–7 days. The caterpillars of the IV instar do not differ in color from the caterpillars of the III instar. After passing into the IV instar, the caterpillars leave the groups and move on to an individual lifestyle. After another 5–7 days, the caterpillars pass into the V instar. In the vast majority of V instar caterpillars, the color of the lateral outgrowths changes from yellow to black, they stop feeding, seek shelter and fall into a stupor ( Fig. 28F View Fig ). A total of 17 V instar caterpillars did not change color and continued to feed. The cessation of feeding and the change in color occurred in these caterpillars a week after the transition to the VI instar. Changes in the behavior and appearance of the caterpillars clearly indicate a state of diapause. The transition to the diapause of caterpillars of III–V instars for many species of Melitaea is an obligate stage of the development cycle (Walberg et al. 2001). Of the 233 caterpillars, 210 (90.1%) survived to the diapause stage. The larvae died mainly from the last batch of eggs laid by the female in the cage. The caterpillars from this group were smaller in size and somewhat slower in development. The main cause of death was a violation of the molting process, caused, apparently, by birth defects of development. The caterpillars collected in nature at the beginning of July and fed further in the laboratory also passed to diapause at V instar. Of the 23 caterpillars of instars III–IV collected in nature, 21 (91.3%) survived to diapause. Thus, the development of M. shahvarica from the moment of egg laying to the departure of the caterpillars into diapause lasts, on average, about a month. In nature, in the conditions of the Shahvar Mountain, the vegetation of Ph. molucelloides at an altitude of 2150–2200 m ends in mid-June. The vegetative and generative parts of the plant completely dry out and the caterpillars of M. shahvarica are not observed on them. At an altitude of 2500 m, the vegetation of Ph. molucelloides continues until the first decade of July, and at this time, we still found caterpillars of III–IV instars of M. shahvarica on the host plants. According to observations in nature, the caterpillars of M. shahvarica actively feed in the morning and before sunset, the rest of the day they are sedentary, sitting on the underside of the leaf or on the soil under the leaves of the host plant. It should be noted that almost all Ph. molucelloides plants have completely sclerotized leaves by the end of the growing season. This indicates a mass feeding of caterpillars and a high density of local populations of M. shahvarica The caterpillars of M. shahvarica are strict oligophages. Under laboratory conditions, we used Phlomoides tuberosa Moench as a host plant, which is systematically close to Ph. molucelloides and grows wild in central Russia. The caterpillars of M. shahvarica rejected the proposed fresh leaves of other plants known as host for the genus Melitaea ( Kolesnichenko 2006) , such as plantain ( Plantago spp. ), veronica ( Veronica officinalis L.), wormwood ( Artemisia spp. ), and other species of Lamiaceae ( Lamium spp. , Mentha spp. ).
We assumed that the caterpillar of M. shahvarica sp. nov., like that of most other members of the genus Melitaea , goes through seven instars in its development and must continue feeding after diapause and go through 1–2 more instars before pupation. Given the weather conditions on the Shahvar mountain, the output of the caterpillars of M. shahvarica from diapause should occur no earlier than the end of April. Unfortunately, we were not able to determine the factor that interrupts the diapause of the caterpillars and complete the development cycle of M. shahvarica in the laboratory. The caterpillars taken out of the refrigerator in March, April, and May did not respond to changes in light conditions, temperature, and humidity, to mechanical stimulation, or to the fresh host plant leaves. They did not show motor activity and soon died. Certain questions are also raised by the contradiction between observations of the development cycle of M. shahvarica in nature and in the laboratory. In nature, the emergence of the imago occurs in early May almost simultaneously with the beginning of the vegetation of the host plant. Obviously, in this case, the butterflies are born from overwintered pupae. Otherwise, the need for further nutrition for the caterpillar that has emerged from the diapause, two more instars and pupation would have shifted the appearance of the imago 2–3 weeks later, to the second half of May. In addition, we did not find in the first half of May any overwintered caterpillars or signs of their vital activity (leaf gnawing, droppings) despite a thorough examination of many host plants. The final answer to the question about the duration and stage of diapause in the development cycle of M. shahvarica should be given by further observations of the development of preimaginal stages both in nature and in the laboratory.
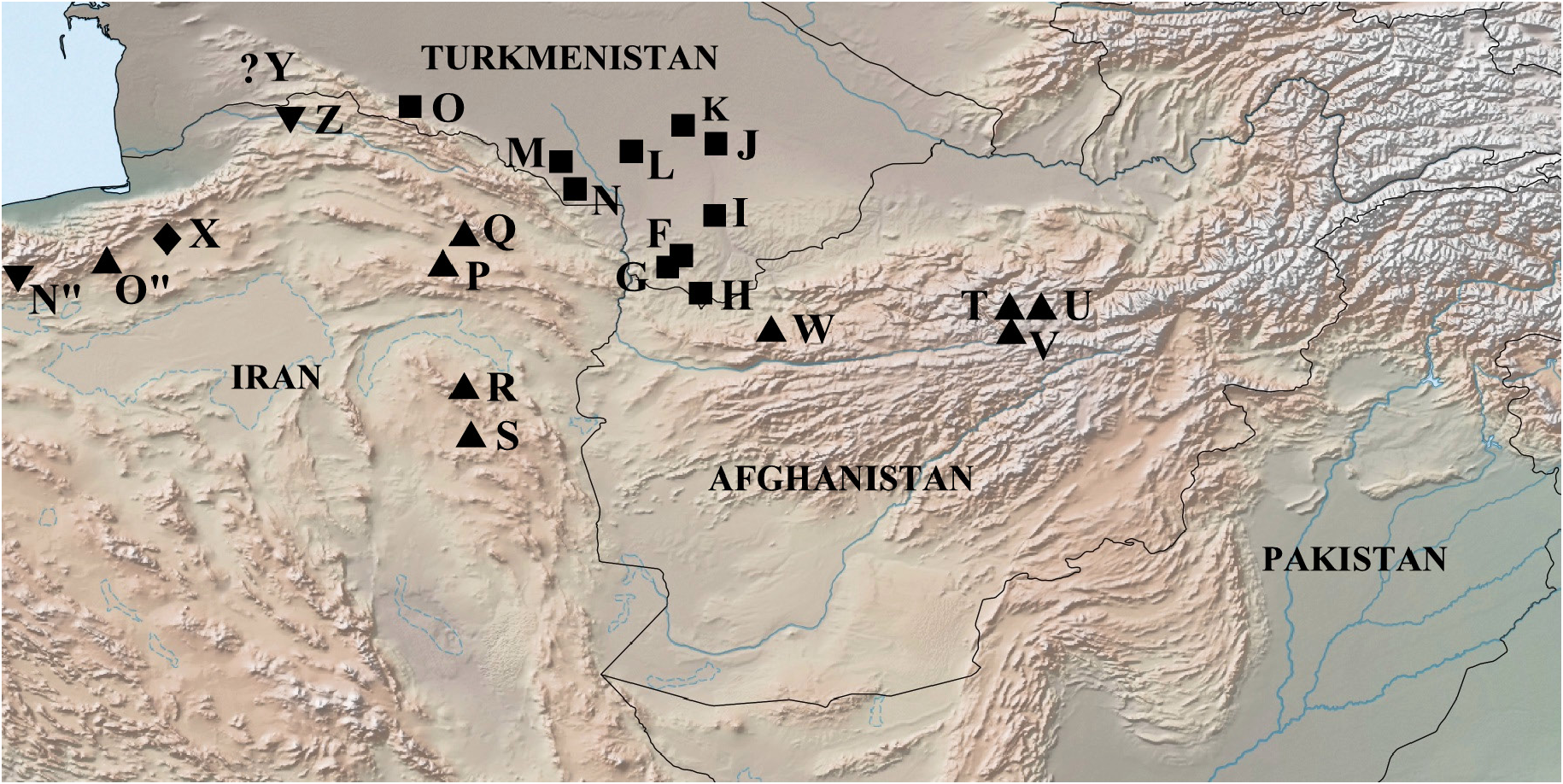
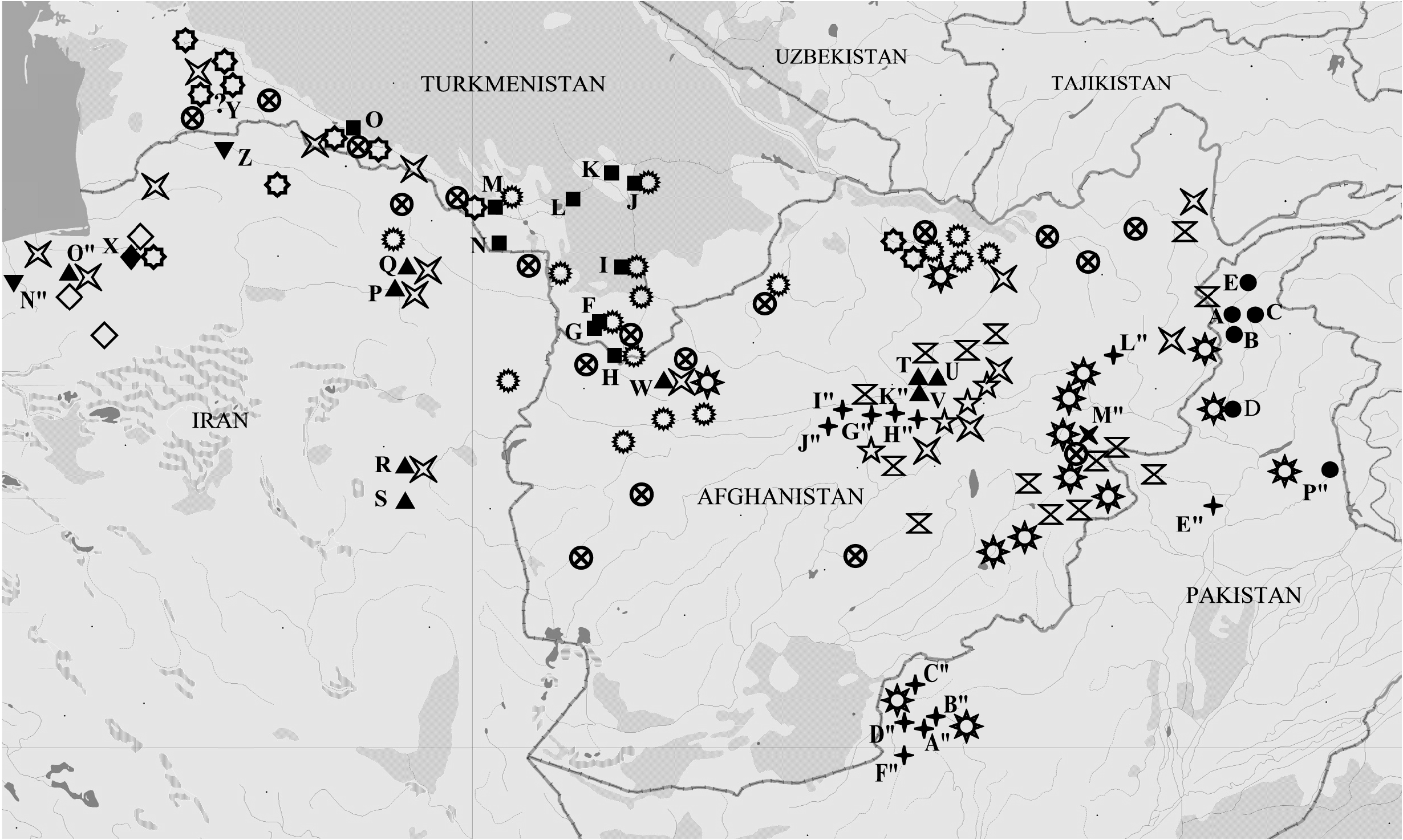
Distribution ( Fig. 14 View Fig )
Iran, Elburs Ridge, Shahvar Mt.
Of undoubted interest are the data ( Tshikolovets et al. 2014; van Oorschot & Coutsis 2014) on the findings of butterflies from the lutko group ( M. lutko mimetica in the authors’ understanding) to the west and north of the habitat of M. shahvarica sp. nov. – on the Elburs Ridge in the vicinity of Demavend Mt. (label: Tehran, Elburz, Demavend, Ask, Lucien leg.) and on the Palyzan Ridge in the northwestern tip of the Turkmen-Khorosan Mountains (label: Golestan, E Maraveh Tappeh, N Ghazan Ghayeh, Palizan Mts). In the first case, it is about a single male, caught on 28 Jun. 1967, which, according to van Oorschot & Coutsis (2014), is characterized by a “transitional” type of the genitalia. However, the same authors expressed doubts about the correct labeling of this specimen on the grounds that for almost 50 years of active collecting of butterflies in the vicinity of Demavend Mt. there are no finds confirming the presence of representatives of the M. lutko group in this area. The presence of M. timandra on the Palyzan Ridge, in our opinion, is more realistic, despite the cardinal differences in the natural and climatic conditions of this area. It is not possible to say anything with certainty about the systematic status of these specimens since the authors of both these publications do not provide images of imago or genitalia. However, based on the characteristics of the habitat and the set of species of potential host plants ( Ph. labiosiformis (Popov) Adylov, Kamelin & Makhm. , Ph. boissieriana (Regel) Adylov, Kamelin & Makhm. ), we believe that systematically the butterflies from Meraveh Tappeh should most likely be closer to M. timandra binaludica subsp. nov. despite the apparent proximity of the Palyzan Ridge to the type locality of M. shahvarica sp. nov. Our assumption is supported by the absence of significant barriers for the spread of M. timandra binaludica to the west, while M. shahvarica , which lives on the slopes of Shahvar Mt, is separated from the northwestern part of the Turkmen-Khorosan Mountains by a high ridge, the northern slopes of which are covered with very moist dense forests.






| SDM |
Stroud and District Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Nymphalinae |
|
Genus |