
Chaetonotus (Chaetonotus) eximius, Kolicka, Małgorzata, Kisielewski, Jacek, Nesteruk, Teresa & Zawierucha, Krzysztof, 2013
|
publication ID |
https://doi.org/10.11646/zootaxa.3717.2.7 |
|
publication LSID |
lsid:zoobank.org:pub:27BD65FD-18CF-4E9D-AE77-C7C0137CF1DC |
|
DOI |
https://doi.org/10.5281/zenodo.6164626 |
|
persistent identifier |
https://treatment.plazi.org/id/EF4A2F6C-6D52-FFD1-FF52-F984FD04FAA7 |
|
treatment provided by |
Plazi |
|
scientific name |
Chaetonotus (Chaetonotus) eximius |
| status |
sp. nov. |
Chaetonotus (Chaetonotus) eximius View in CoL sp. nov. ( Figs 13–17 View FIGURE 13 View FIGURE 14 View FIGURE 15 View FIGURE 16 View FIGURE 17 ; Table 6 View TABLE 6 )
Locality: Sites 1, 2, 3; session I, II, III.
Material: 3 samples, 22 specimens ( 20 adult and 2 juvenile), 17 of them have been photographed. The photographs of holotype and three paratype are available in Natural History Collection at Adam Mickiewicz University in Poznań under accesion number NHC-GCCE- 2-1-10 /h ( holotype) and NHC-GCCE- 2-11-41 /p ( paratypes). Additional all photographs are available at Adam Mickiewicz University in Poznań, in the collection of the first author.
Etymology: From Latin “ eximius ”—unique, remarkable, extraordinary.
Diagnosis: Body length from 86.4 to 95.8 Μm. Stocky body without neck constriction. Semicircular five–lobed head of seemingly three–lobed appearance, epipleuria small and shifted to the dorsal part of the head present. Hypopleuria approximately three times longer than epipleuria and visible in the head outline. Hypostomium elliptical in shape. Two pairs of tufts of cephalic cilia, 8.5–15.4 Μm in length. Mouth ring 4.6–5.4 Μm in diameter located ventrally. Pharynx 20.4–28.3 Μm in length with two dilatations at the ends and distinct cuticular reinforcements in the anterior part. Furca of the length 12.8–15.4 Μm, parted externally, spaced apart from the body width. The adhesive tubes 8.1–10.4 Μm long, strongly curved outwards. Sensory bristles anchored by double– keeled scales. The whole body, except for the ventral interciliary field, covered with one–lobed scales with pronounced keels and spines, arranged in 22–26 longitudinal alternating rows and 29–34 scales in each row. Scales of triangular shape with rounded edges and posterior notches, located close and next to one another, in the middle body section touching the edges but never overlapping. Spines without lateral denticles. Posteriormost lateral spines straight, not longer and not thicker than the trunk spines, even slightly shorter than the longest trunk spines. Ventral interciliary field covered with minor, oval scales without keels, but with short, thin spines. Ventral interciliary terminal scales elongated, oval–shaped, with keels and long straight spines without lateral denticles. Ocellar granules absent.
Description: Chaetonotus (Chaetonotus) eximius sp. nov. is a small species. The body is stocky and neck narrowing is absent. The five–lobed head has semicircular shape with a developed cephalion (U1–U4) and two pairs of pleurs (U3–U10). The cephalion and hypopleuria are well visible in the body outline, while the epipleuria are small and shifted to the dorsal part of the head, and thus not visible in the outline ( Fig. 14 View FIGURE 14 ). The hypopleuria are approximately three times longer than the epipleuria and are located at U4–U10. This arrangement of plates creates an apparent impression of a three–lobed head. Two pairs of cephalic ciliary tufts are present on the head. The first pair (three cillia per tuft) is located between the cephalion and the epipleuria (shorter cilia), the second pair of ciliary tufts (four cillia per tuft) arises between the epipleuria and the hypopleuria (longer cilia). Ocellar granules are absent. The mouth ring is situated ventrally at U2–U5. In the mouth ring there are reinforcements typical of the genus Chatonotus. The hypostomium is elliptical in shape. Near the anterior hypostomium edges there are weak transverse cuticular bars. The neck width does not differ from the head width, and gradually passes into the trunk (U34–U86). The trunk slightly and gradually expands to ca. U65 and subsequently slightly and gradually tapers from U77 to the furca base. The furca begins at U87. The furca branches arranged unparallel and tips of adhesive tubes are widely apart. Thick adhesive tubes, slightly tapered to the end (U92–U100) and strongly curved in two dimensions: dorsal–ventral and laterally are present ( Fig. 15 View FIGURE 15 ; 17B).
The head, neck and trunk on the dorsal, lateral and ventral sides are covered with one–lobed scales with pronounced keels ( Fig. 17 View FIGURE 17 C) and spines without lateral denticles. On the head, neck, and trunk the scales show uniform shape. They are triangular with rounded edges and posterior notches; distinct keels begin directly behind the anterior scale edges and pass into the spines. The scales are arranged in 22–26 longitudinal alternating rows, 29–34 scales in each row. The scales are arranged directly next to one another, in the middle body section (U47– U73) they touch the edges of neigbouring scales, but never overlap. The longitudinal rows of scales begin directly after the cephalion and pleurs and run parallel to the furca base. The length and width of the scales gradually increase from the head, through the neck to the widest trunk part (U65–U76), and subsequently gradually decrease towards the furca base (U87). The spines are curved, without lateral denticles and gradually tapering towards the end. Similarly to the scale size, their length increases to the widest trunk part and subsequently gradually decreases towards the furca base. There are two pairs of sensory bristles anchored at the double–keeled scales. The anterior pair of the double–keeled scales is located at U34–U36, the posterior pair of them is located at U80–U83.
On the ventral surface longitudinal ciliary bands start at U5 and end at U84. The interciliary field (U10–U84) is covered with one–lobed scales arranged close and next to one another. They are situated in alternating rows of 9–11 scales. The ventral field scales are oval without keels, with short, tiny spines without lateral denticles. A pair of ventral terminal scales, at U84–U89, is significantly longer than the other ventral scales and of an oval, elongated shape ( Fig. 17 View FIGURE 17 B). The ventral terminal scales possess weak keels and long, straight spines extending beyond the furca indentation.
The pharynx (U3–U32) has a well–developed anterior dilatation (U3–U9) and a posterior dilatation (U26– U32) wider than the anterior one. A very important feature is the presence of strong cuticular reinforcements inside the pharynx ( Fig. 17 View FIGURE 17 A), localised at U5–U8. These reinforcements are composed of two merged cuticular rods and cuticular granules. The pharynx is connected via the pharyngeal intestinal junction (U33) to the straight intestine without a separate enzymatic section (U32–U84).
Taxonomic remarks: The new species shows affiliations to species of the genus Heterolepidoderma and Halichaetonotus . From all of the described Chaetonotidae , only in six freshwater species have cuticular reinforcements in the anterior part of the pharynx: in C. (H.) furcatus Kisielewski, 1991 (as described below), C. (C.) intermedius Kisielewski, 1991, C. (C.) pawlowski Kisielewski, 1984, Heterolepidoderma jureiense Kisielewski, 1991 , H. dimentmani, Kisielewski 1999 , and H. joermungandri Kånneby, 2011 . Also, in marine species from the genus Heterolepidoderma , cuticular reinforcements in the anterior part of the pharynx are present. In the genus Halichaetonotus , cuticular reinforcements in the anterior part of the pharynx are a regular element of the construction, however, this genus has only been reported in marine habitats. C. (C.) eximius sp. nov. is the third identified species from the genus Chaetonotus having this type of reinforcement in the pharynx, and fourth from among congeneric species at all with cuticular reinforcements in the pharynx.
A—Dorsal view, B—View of internal morphology, C—Ventral view. All measurements in micrometers.
Representatives of three different genera, which according to morphological data (Hochberg & Litvaitis 2000), come from one branch of a phylogenetic clade and have a very similar cuticular structure at the same position of the pharynx may be explained as an adaptation to a specific habitat and kind of food i.e., convergence. However, this may also indicate a closer, i.e. than was previously thought, phylogenetic relationship between representatives of Chaetonotus , Halichaetonotus and Heterolepidoderma . Knowledge on the polyphyly of the genera Chaetonotus and Heterolepidoderma suggests that a closer relationship may be possible (Kånneby et al. 2013).
Differential diagnosis: Of all the hitherto known congeneric species, Chaetonotus (Chaetonotus) eximius sp. nov. seems to share few features with C. (C.) brevispinosus Greuter, 1917 , C. (C.) disiunctus Greuter, 1917, C. (C.) fluviatilis Balsamo & Kisielewski, 1986, C. (H.) furcatus Kisielewski, 1991, C. (C.) intermedius Kisielewski, 1991, and C. (C.) pawlowskii Kisielewski, 1984. In all of the compared species the adhesive tubes are not curved as in C. (C.) eximius sp. nov., and they are also thinner.
C. (C.) brevispinosus Greuter, 1917 (90–190 Μm in length) has a well developed neck constriction in contrast to the new species. Moreover, epipleuria are visible in the head outline and the pharynx is much wider (formula a: 37–39%; m: 29–36%; p: 39–47%). The shape of the scales (rounded anterior edges and absent keels) is different than in the newly described species. Scales distribution is also different (27–33 longitudinal alternating rows of scales, 21–30 scales in each row). Furthermore, in C. (C.) brevispinosus the last pair of the dorsal spines is longer than the other body spines (5–12 Μm), and the furca with the adhesive tubes is significantly narrower, which does not occur in the new species.
C. (C.) disiunctus Greuter, 1917 (120– 256 Μm in length) is a larger species than C. (C.) eximius sp. nov. This species has a different body shape and proportions, and the pleurs are well developed and distinctly marked in the head outline. The scales in this species are one–lobed without keels and with a shallow posterior indentation. Furthermore, the scales are longer (neck: 4–5 Μm; trunk: 5–8 Μm) and differently distributed (20–27 scales in single longitudinal row and 17–19 longitudinal alternating rows of scales). Additionally, the last lateral spines of C. (C.) disiunctus are markedly longer than the other spines (11–30 Μm).
C. (C.) fluviatilis Balsamo & Kisielewski, 1986 (107–140 Μm in length) is a larger species. Its body is slender. The head is rounded like in the new species, but it is wider than the neck and has larger epipleuria. The scales in C. (C.) fluviatilis are three–lobed and of varied shapes (neck: 4–4.5 Μm; trunk: 5–8.5 Μm), located at large distances from one another. Additionaly, scale distribution on this species is clearly different than in the newly described taxa (13–14 longitudinal alternating rows of scales, 13–14 scales in each row). Furthermore, in this species the furca is definitely longer (21–23 Μm) and only one pair of sensory bristles (the posterior one), instead of two, is present.
C. (C.) furcatus Kisielewski, 1991 (134–148 Μm in length) is a larger species. This species has three different dorsal types of three–lobed scales with various spines, including one type with a distal furcation. In the anterior part of the pharynx the species has one pair of thin, not merging cuticular rods that are comparable to typical reinforcements. Moreover, scale distribution is also different (17–18 longitudinal alternating rows of scales, 15–17 scales in each row). The head is clearly five–lobed with the posterior portion of the cephalion free, large hypopleuria, and head is considerably wider than the neck. Moreover, the ventral interciliary field in this species are covered with two different kinds of scales (scales with rudimentary spines and keels and spined scales) unlike C. (C.) eximius sp. nov. The next clear difference between the two compared species is the adhesive tubes (8.5–9.5 Μm), which are straight and thin and are slightly thinning distally in C. (H.) furcatus.
A—Ventral body side, B—View of internal morphology. All measurements in micrometers.
C. (C.) intermedius Kisielewski, 1991 (131–153 Μm in length) is a larger species. This is the only species hitherto described from the genus Chaetonotus that has cuticular reinforcements of medium size and thickness in the pharynx. The reinforcements in this species consist of one pair of cuticular pharyngeal rods occurring anteriorly, like in C. (C.) eximius sp. nov., but C. (C.) intermedius has no cuticular granule under the rods, and the rods are thinner. The head in C. (C.) intermedius is much narrower than the trunk and has a clearly five–lobed outline and protruding posterior pleurae. The scales in C. (C.) intermedius are of a long oval shape, with the posterior edges either rounded–off or slightly cut, and they are arranged similarly to the scales in the genus Heterolepidoderma (24–27 longitudinal alternating rows of scales, 25–28 scales in each row in C. (C.) intermedius), unlike the new species. In C. (C.) intermedius the spines are thin and short (1.5 Μm), and only in the ventrolareral and posterior part of the body are the spines longer (7–9.5 Μm). Furthermore, C. (C.) intermedius has a ventral, interciliary field covered only with one pair of terminal scales, with short spines, unlike C. (C.) eximius sp. nov.
C. (C.) pawlowskii Kisielewski, 1984 (167–172 Μm in length) is a larger species with a complicated system of cuticular reinforcements in the pharynx which do not correspond to those of the new species. The reinforcement system in this Chaetonotus consists of four pairs of cuticular rods, whose most anterior pair begins at the level of the mouth. Additionally, in the posterior pharynx dilatation selvages (system of cuticular rims) occur. Moreover, the pharynx of C. (C.) pawlowskii consists of two conspicuous dilatations (“bulbs”) and a short median section that is distinctly thinner, unlike C. (C.) eximius . Other features that differentiate it from the new species are an extremely well–developed hypostomium which adjoins the anterior pharynx dilatation, the head shape and a broad furca base with adhesive tubes parallel to one another. Furthermore, this species has a different scale shape, with the neck and trunk spine lengths overlapping with the length of these structures in the newly described taxa (3–5.5 Μm and 3–4.5 Μm, respectively). Moreover, their distribution is different (22–25 longitudinal alternating rows of scales, 23–30 scales in each row). Another difference is considerably shorter pair of rearmost lateral spines (1–1.5 Μm).
A—Pharynx with cuticular reinforements, B—Posterior body region with visible scales of interciliary field, C—Dorsal view of scales in trunk region. All measurements in micrometers.
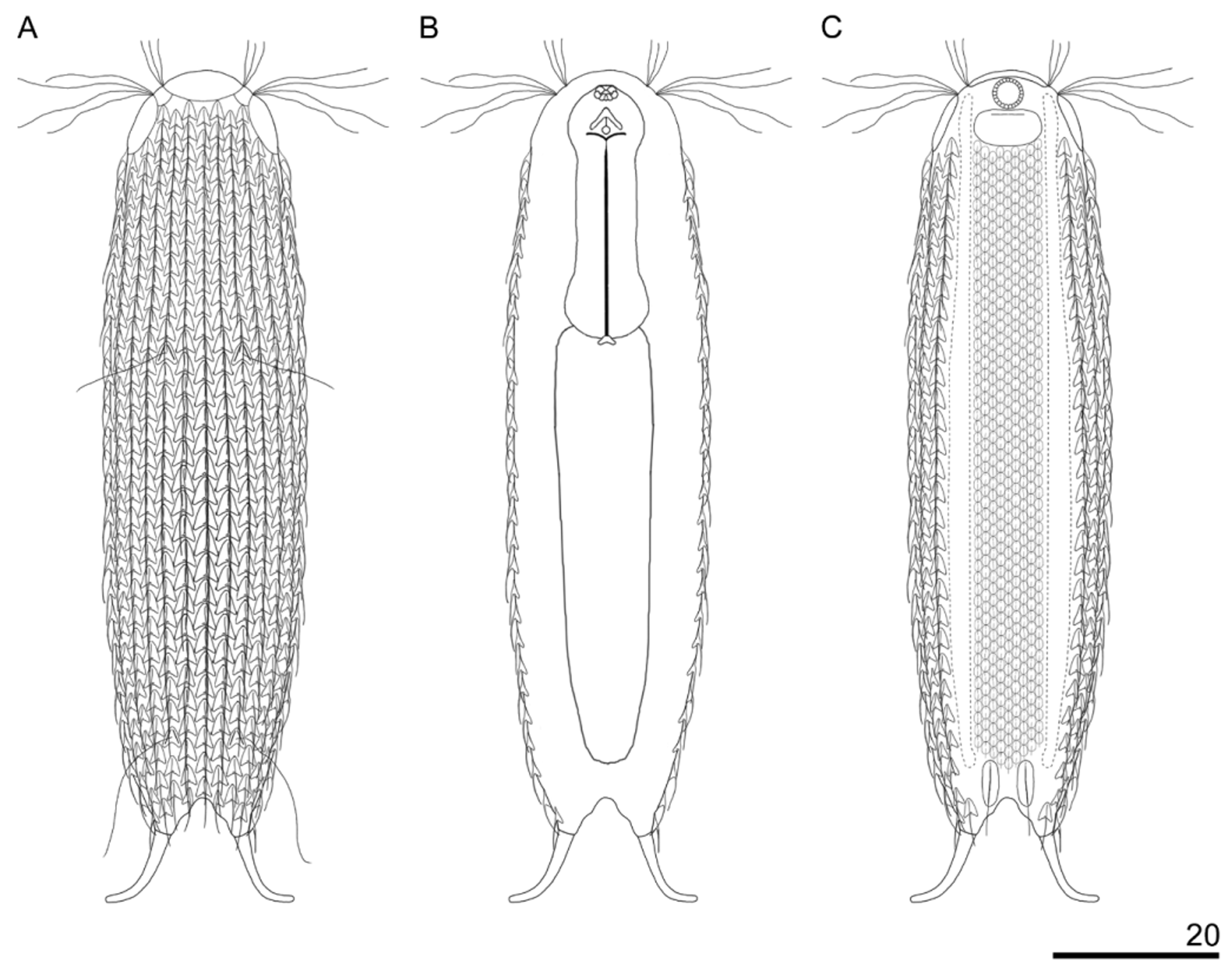

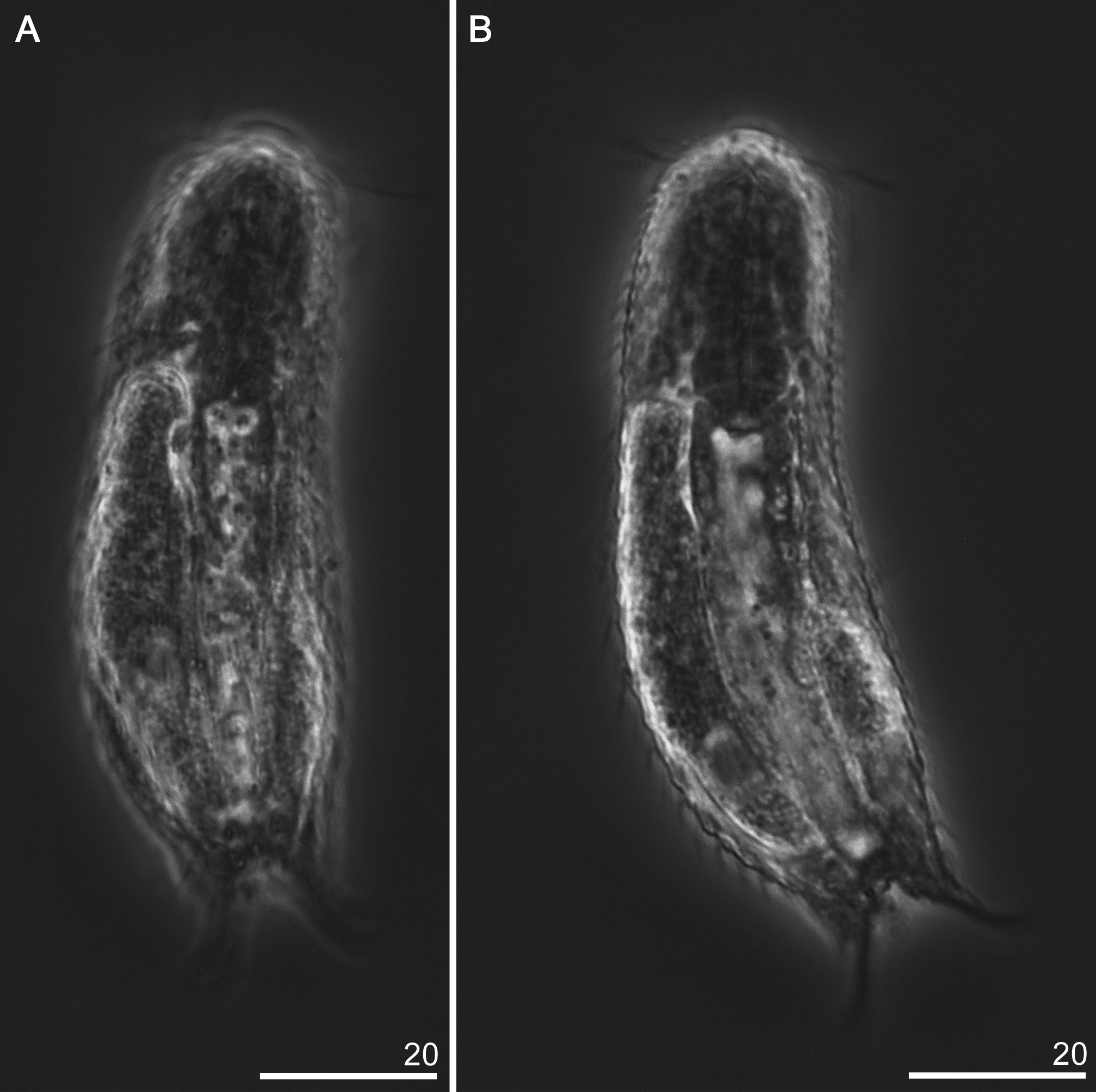


TABLE 6. Morphometric parameters for Chaetonotus (Chaetonotus) eximius sp. nov.; N— number of specimens or structures analysed, Range—the smallest and the largest structure found among all specimens measured, SD—standard deviation,? — structure oriented in a way unsuitable for measurement.
| Character | N Holotype Range | SD | Juvenile specimen |
|---|---|---|---|
| Body length | 14 90.2 86.4–95.8 | 2.71 | 86.4 |
| Pharynx length | 15 25.6 20.4–28.3 | 2.24 | 23.9 |
| Width of anterior pharynx thickening (a) | 6 8.7 6.6–8.9 | 0.95 | ? |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Order |
|
|
SubOrder |
Paucitubulatina |
|
Family |
|
|
SubFamily |
Chaetonotinae |
|
Genus |
|
|
SubGenus |
Chaetonotus |