
Paraphenice fluctus Bahder, Stroiński & Bartlett, 2024
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5406.3.5 |
|
publication LSID |
lsid:zoobank.org:pub:F49F1B8A-ECD4-47E8-972E-8FC37D1ED472 |
|
DOI |
https://doi.org/10.5281/zenodo.10630649 |
|
persistent identifier |
https://treatment.plazi.org/id/A10187AC-FF90-C76F-42AB-DAEBFE4DFEA3 |
|
treatment provided by |
Plazi |
|
scientific name |
Paraphenice fluctus Bahder, Stroiński & Bartlett |
| status |
sp. nov. |
Paraphenice fluctus Bahder, Stroiński & Bartlett , sp. n.
( Figures 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 )
Type locality. Madagascar, Hotel Feon’ ny Ala, Alaotra-Mangoro Region .
Diagnosis. Moderate-sized planthopper (~ 6.7 mm), general body color yellow to testaceous. Antennal pedicel large and bulbous. Wings transparent with median fuscous markings. Pygofer with large triangular projection of posterior margin in lateral view; medioventral process in ventral view strongly asymmetrical, left side round, invaginating at apex and right side projecting in irregularly sinuate process, slight more sclerotized on right side. Gonostyli with sclerotized process at midpoint angled ventrad on outer lateral sides. Aedeagus with multiple sclerotized processes with serrations at apex and highly complex endosoma, twisting and reaching base of aedeagal shaft.
Description. Color. General body color yellowish, darker dorsally, paler ventrally ( Figs 2 View FIGURE 2 & 3 View FIGURE 3 ), abdominal terga reddish. Second segment of antennae white basally, yellow distally. Forewings transparent (veins mostly white), weakly embrowned medially bearing dark spot at subapically fork of MP 1 ( Figs 3 View FIGURE 3 , 5 View FIGURE 5 ).
Structure. Body length (including wings) males: 6.6–6.7 mm (n = 11), females: 6.7–6.9 mm (n = 9). Head. In dorsal view, vertex elongated, approximately 3x long as wide, extending beyond eyes, lateral margins foliate, disc depressed, median carina absent; anterior margin strongly concave, posterior margin truncate, transverse carina present at fastigium, two complete rows of sensory pits along lateral margins, third incomplete row mesad ( Fig. 4A View FIGURE 4 ). In lateral view, head rounded (vertex weakly inclined and convex, frons arched), extending moderately beyond eye margin, slightly angled at fastigium (at transverse carina); In frontal view, lateral margins of frons sub-parallel, single row of sensory pits along lateral margins extending from dorsal margin to clypeus ( Fig. 4C View FIGURE 4 ). Frontoclypeal suture truncate, clypeus very elongately triangular. Eyes reniform in front of antennae. Lateral ocelli present, white, situated just below eye at posterior margin ( Fig. 4B View FIGURE 4 ). Antennae with scape ring-like, pedicle segment large and bulbous bearing two differently sized sensory plaques, smaller plaques over entire surface in uniform distribution, larger clustered over the apical 1/3 of segment ( Fig. 4 View FIGURE 4 ). Subantennal genal crest present (arched along ventroposterior antennal margin), slightly surpassing basal margin of second antennal segment in frontal view ( Fig. 4C View FIGURE 4 ).
Thorax. Pronotum tricarinate, in dorsal view with median carina and carinae at lateral margin (from dorsal view) between tegulae and head, anterior margin in dorsal view strongly convex, posterior margin broadly triangularly excised ( Fig. 4A View FIGURE 4 ). Mesonotum in dorsal view tricarinate, median carina becoming obsolete posteriorly near scutellum, lateral carinae arising laterad, extending transversely, then abruptly angled (nearly 90°), then longitudinally (slightly sinuate), becoming obsolete before caudal margin ( Fig. 4A View FIGURE 4 ); in lateral view, mesonotum humped (concave before upturned scutellum), bearing pair of crests near anterior margin of scutellum. Spinulation of hind tibia 5-6-5.
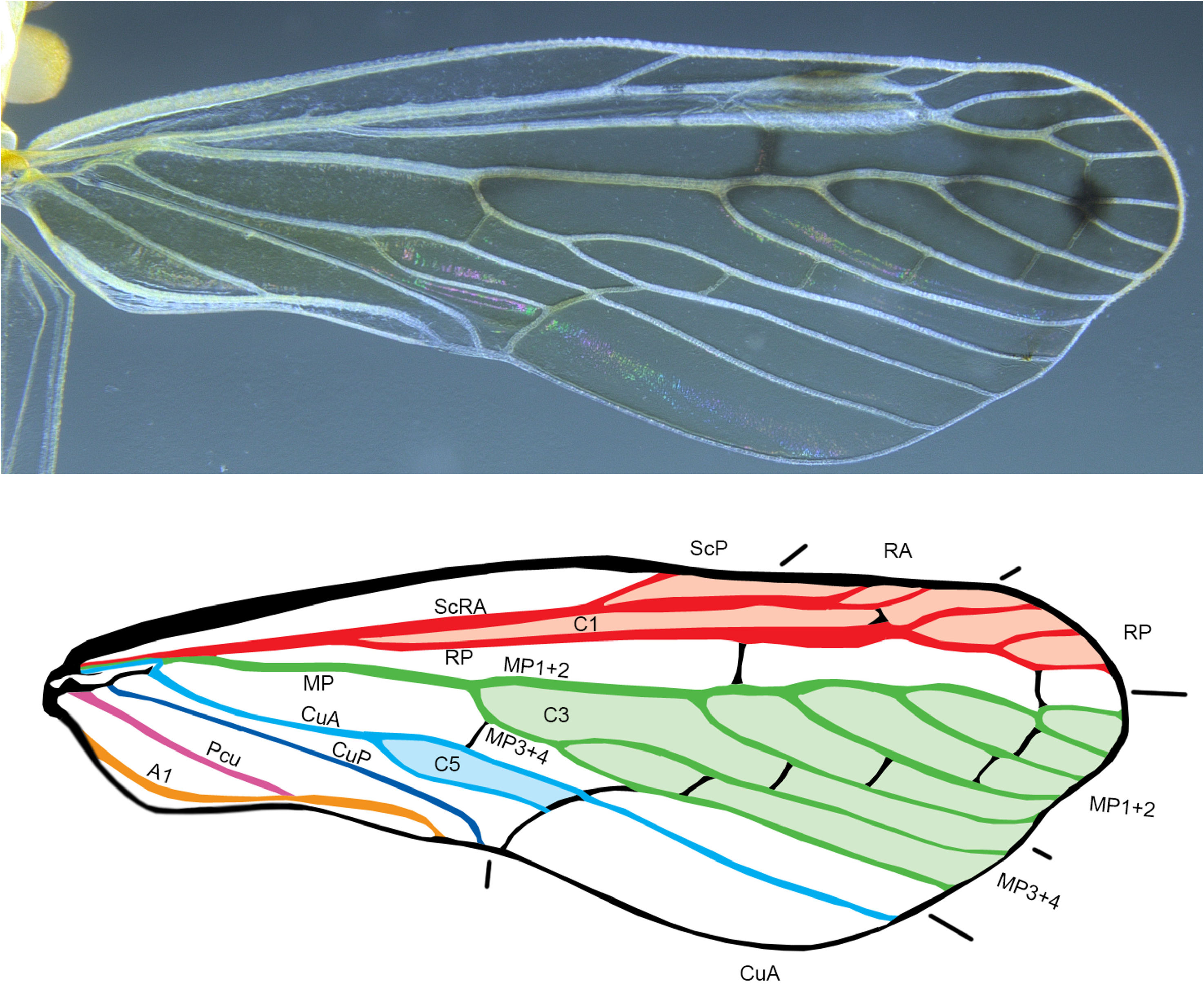
Forewing ( Fig. 5 View FIGURE 5 ) elongate, spatulate, approximately 2.5 times longer than broad (at greatest width), costal margin convex (bearing small pits near midlength), trailing margin expanded between claval apex and wing tip. Clavus closed (Pcu+A1 reaching wing margin well before CuP). Basal cell narrow and elongated; ScP+R+MP arising from leaving basal cell as common stem; MP forked after short common stem; RP forked from Sc+RA in basal third of wing just before fork of CuA, fork of MP 1+2 from MP 3+4 before claval apex. CuA anastomosing to form closed C5 (procubital) cell. Veins Pcu and A 1 fused just after claval midlength. Branching pattern RA 2-branched, RP 3-branched, MP 7-branched.
Hindwing ( Fig. 6 View FIGURE 6 ) almost as long as forewing with large basal cell. ScP+RA ending with single terminal about middle of costal margin; RP unbranched, MP single, CuA with 3 terminals, CuP single, Pcu single, A 1 with 2 terminals, A 2 single; stridulatory plate with outer margin weakly concave; crossveins r-m and m-cu at apical third of hindwing.
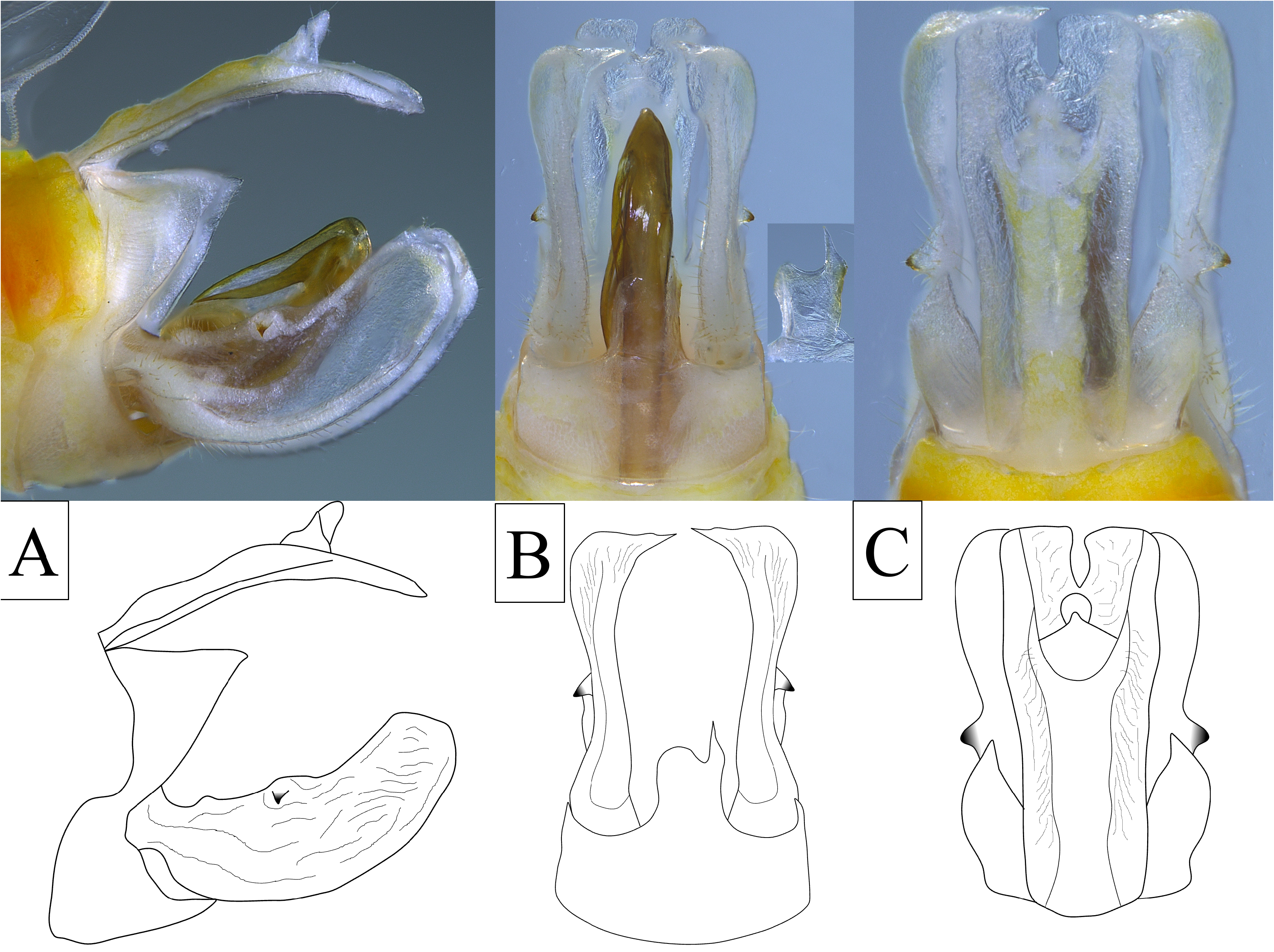
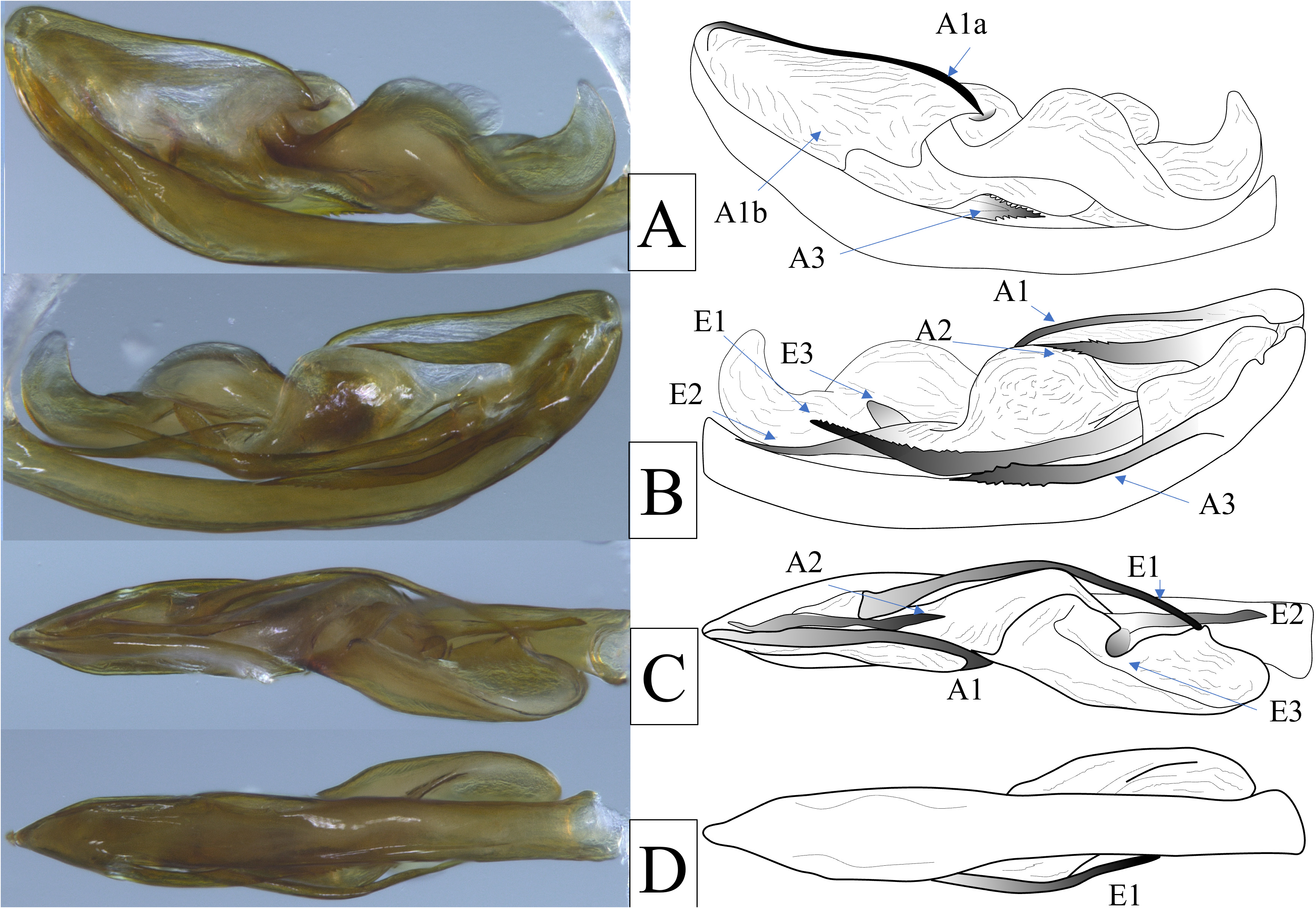
Genitalia. Pygofer in lateral view irregular, dorsal margin appearing to not reach dorsal margin, irregularly sinuate on anterior margin, lateral margins of pygofer opening bearing large, triangular lobes, constricted ventrally near midlength, expanded again to ventral margin ( Fig. 7A View FIGURE 7 ); in ventral view, medioventral process strongly asymmetrical, longer than wide, broadest at base, left lateral margin apically rounded, right lateral apex bearing acuminate and irregularly sinuate process ( Fig. 7B View FIGURE 7 ). Gonostyli in lateral view broadly and elongately spatulate, narrowest at base, irregularly sinuate on broadly concave dorsal margin bearing laterally projected sclerotized uncus at midpoint ( Fig. 7C View FIGURE 7 ), ventral margin broadly convex, apex bluntly rounded ( Fig. 7A View FIGURE 7 ); in ventral view, inner and outer margins sinuate, broad at base, constricting near midpoint, expanding subapically, apices truncated with elongate median points ( Fig. 7B View FIGURE 7 ). Aedeagus complex, bilaterally asymmetrical, shaft simple bearing three large retrorse sclerotized processes arising near apex; the first process (A1) arises at apex on right lateral side slender dorsal spine (A1a; sinuate, curving slightly ventrad) and weakly sclerotized, transparent broad flange entrally (A1b); second process (A2) arising subapically on left lateral side near dorsal margin, robust, sinuate, apex finely serrate, nearly reaching apex of A1; third process (A3) arising subapically on left lateral side longer and more slender than A2, apex irregularly apically serrated (both dorsally and ventrally) curved slightly dorsad, ( Fig. 8 View FIGURE 8 ); endosoma large and complex with twisted and wavy lingulate appearance, reaching base of aedeagal shaft bearing three elongated sclerotized processes; one large process (E1) arising near base, curved dorsad, sfinely serrated dorsally in distal half, second process (E2) arising on left ventral margin near midpoint, sinute, not serrated; third process (E3) arising near E2, angled mesad, spatulate, angled dorsad ( Fig. 8 View FIGURE 8 ). Anal t in later view narrow, irregularly sinuate on dorsal margin, curved along ventral margin to elongated ventrocaudally directed apex, apex nearly reaching apex of gonostyli ( Fig. 7A View FIGURE 7 ); in dorsal view, broad, quadrate, apex truncate, medially deeply notched ( Fig. 7C View FIGURE 7 ).
Plant associations. Livistona chinensis (Jacq.) R. Br. ex Mart. and Raphia P. Beauv. ( Arecales : Arecaceae ).
Distribution. Central-eastern Madagascar (Alaotra-Mangoro Region, Anosibe An’ala District, Andasibe commune; Atsinanana Region, Toamasina II District, Mahavelona (Foulpointe) commune, Analalava Forest Reserve).
Etymology. The specific name given, “ fluctus ” is Latin for ‘wave’ in reference to the large, wave-like appearance of the endosoma, and is intended as indeclinable.
Material examined. Holotype, male:“ Madagascar, Alaotra-Mangoro , Andasibe / Hotel Feon’ ny Ala / 26.I.2023, sweeping Livistona palm / Coll.: B.W.Bahder // Holotype / Paraphenice fluctus ♂ /” ( FLREC) . Paratypes: 8 males, 9 females same locality as holotype, FLREC ; 2 males: “ Madagascar, Atsinana , Analalava / Reserve lodge at entrance / 27.I.2023, sweeping Raphia palm / Coll.: B.W.Bahder ” ( FLREC) .
Sequence Data. For the novel taxon, a 713 bp was generated for the barcoding region of COI, a 1,498 bp product for the 18S rRNA gene, and a 754 bp product for the D9–D10 region of 28S. All accession numbers are presented in Table 2 View TABLE 2 .
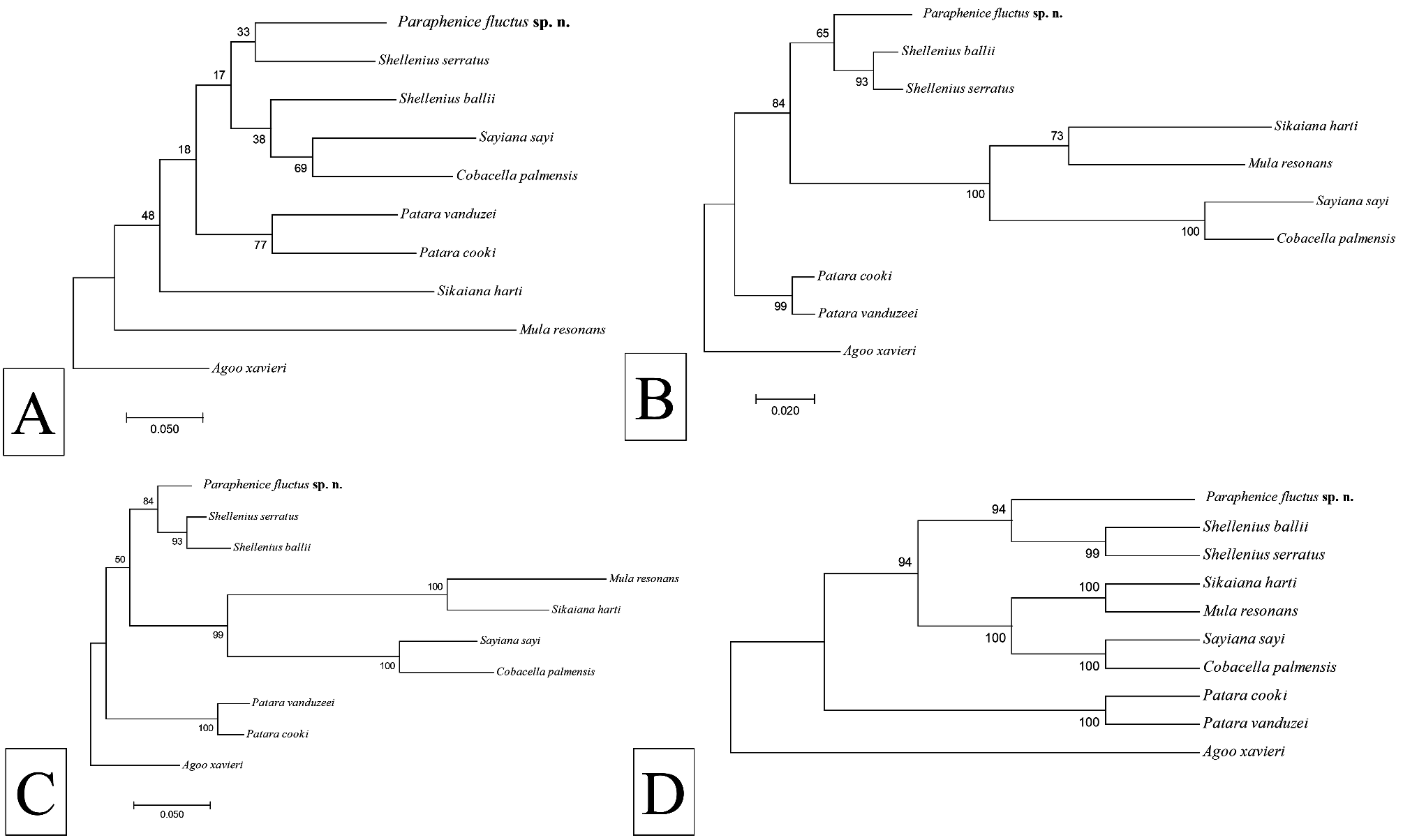
In the absence of molecular data from Phenicini and other Old World Otiocerinae , Paraphenice fluctus sp. n. resolved adjacent to Shellenius Ball with varying degrees of support for all loci as well as for the consensus tree based on all loci ( Fig. 9 View FIGURE 9 ). Bootstrap support in the COI tree was weak (<50), although support was stronger in the 18S tree (65) and 28S trees (84). The strongest bootstrap support (94) for placement of P. fluctus sp. n. next to Shellenius was observed from concatenated data for all three loci ( Fig. 9D View FIGURE 9 ).
Remarks. While Old World taxa were not included and with a relatively small number of taxa analyzed, the closer apparent relationship of Paraphenice fluctus sp. n. to Shellenius (Otiocerini) than the relationship of Shellenius to C. palmensis and S. sayi (also Otiocerini), suggests that the monophyly of tribes in the Otiocerinae needs to be more clearly established. Of the species currently described in Paraphenice , there appears to be substantial variability in form of the anal segment, gonostyli, and medioventral process of the pygofer. In general, there appears to be a common pattern of the aedeagus among the described taxa; possessing a large, complex endosoma that is twisted with multiple lobes/projections.
Among Phenicini , Paraphenice appears to be the only genus with subantennal ridges. Among members of Paraphenice , the closest to P. flucutus sp. n. appears to be P. arebiensis Synave, 1973 (figs 389–392), but this species has dark forewings, a strongly downturned apex of the anal tube, more broadly spatulate gonostyli, and different sizes of processes on the right side of the aedeagus ( Synave 1973 does not illustrate the pygofer or left side of aedeagus). The general forewing pattern appears similar among species of Paraphenice and while there likely exists variation in features of branching, crossvein positions, and cell morphology, the color pattern appears to vary from species to species (where it has been illustrated), and as such, P. fulcutus sp. n. appears to have a unique color pattern, in this case almost lacking a pattern some slightly fuscous areas but with a prominent, subapical black spot. Finally, while there are some species of Paraphenice with distinct and sometimes asymmetrical medioventral processes of the pygofer, the form observed in P. flucutus sp. n. diverges significantly from these other forms. Geographically, P. mawae Wilson, 1987 is the closest species to P. flucutus sp. n., having been documented in Tanzania, however differs significantly in many features ( Wilson 1987, figs 57–65). All other Paraphenice are known from western Africa.




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Otiocerinae |
|
Tribe |
Phenicini |
|
Genus |