
Albanerpeton galaktion Fox & Naylor, 1982
|
publication ID |
https://doi.org/ 10.5281/zenodo.5378709 |
|
persistent identifier |
https://treatment.plazi.org/id/038C6153-283F-FFE1-FF79-FB6BFE98FBF5 |
|
treatment provided by |
Marcus |
|
scientific name |
Albanerpeton galaktion Fox & Naylor, 1982 |
| status |
|
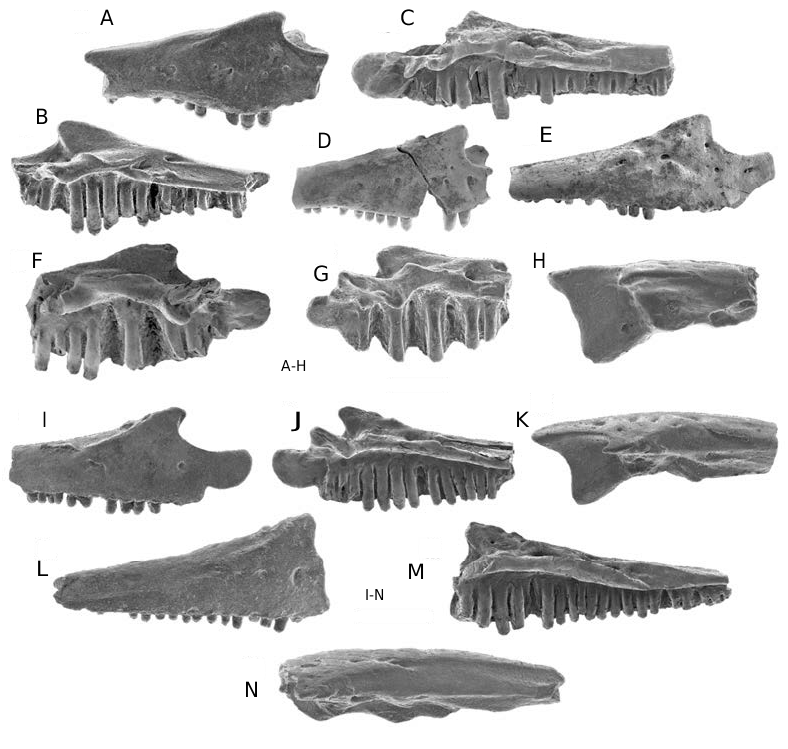
Albanerpeton galaktion Fox & Naylor, 1982 ( Figs 2 View FIG J-M; 3I-N; 4; 6F-K; Tables 2; 3)
Albanerpeton nexuosus Estes, 1981: 24 (in part: referred jaws from the Milk River Formation)
Albanerpeton galaktion Fox & Naylor, 1982: 121-127 , figs 1a, b, d-h; 2-4 (original description)
HOLOTYPE. — UALVP 16203 , nearly complete left premaxilla missing maxillary process and dorsomedial end of pars dorsalis, and having five complete and three broken teeth (Fox & Naylor 1982: figs 1a, b; 2ac) ( Fig. 4A, B View FIG ).
HOLOTYPE HORIZON AND LOCALITY. — Upper Cretaceous (lower Campanian; i.e. Aquilan in age) Deadhorse Coulee Member , Milk River Formation ; UALVP MR-6, Verdigris Coulee , southeastern Alberta.
REFERRED SPECIMENS. — Deadhorse Coulee Member, Milk River Formation. Eight localities, Alberta: UALVP MR-2: UALVP 16218, dentary; UALVP MR-4: UALVP 16255, maxilla; UALVP 39940, 39941, dentaries; UALVP 39950, frontals; UALVP MR-6: UALVP 16210-16212, 39930, 39934, premaxillae; UALVP 16240, 39935, 39936, maxillae; UALVP 16219, 16236, 39939, 39942, dentaries; UALVP 16216, 39946, 39948, 39949, 39951, 39952, frontals; UALVP MR-8: UALVP 39931, premaxilla; UALVP MR-9A: UALVP 39932, premaxilla; UALVP MR-9B: UALVP 16217, dentary; UALVP MR-12: UALVP 16221, dentary; UALVP 39945, frontals; UALVP MR-20: UALVP 16204, 16205, 16213, 39933, premaxillae; UALVP 16241, 39937, 39938, maxillae; UALVP 16222, 39943, 39944, dentaries; UALVP 39947, frontals.
Oldman Formation. Five localities, Alberta: RTMP L0406: RTMP 95.177.81, premaxilla; RTMP L1127: RTMP 96.78.186, premaxilla; RTMP L1128: RTMP 96.78.118, 96.78.184, premaxillae; RTMP L1131: RTMP 96.78.122, premaxilla; RTMP L1137: RTMP 96.78.124, premaxilla.
Kaiparowits Formation. OMNH V6, Utah: OMNH 60326, premaxilla.
Lance Formation. UW V-79032, Wyoming: UW 14593, premaxilla.
DISTRIBUTION ( Table 1). — Upper Cretaceous (Campanian and Maastrichtian), North American Western Interior: lower Campanian (Aquilan in age): Deadhorse Coulee Member, Milk River Formation, Alberta ; middle Campanian (Judithian in age): Oldman Formation , Alberta ; and Kaiparowits Formation, Utah ; upper Maastrichtian (Lancian in age): Lance Formation , Wyoming .
REVISED DIAGNOSIS. — Large-bodied species of Albanerpeton differing from all other albanerpetontid species in two premaxillary autapomorphies: lingual opening of suprapalatal pit occupying about one-fifth to one-quarter lingual surface area of pars dorsalis and palatal foramen diameter greater than one and onethird diameter of medial teeth. Differs further from most congeners, but resembles A. cifellii and Albanerpeton gracilis n. sp. in one premaxillary synapomorphy: suprapalatal pit triangular- to slit-shaped in lingual outline. Primitively differs from A. cifellii in having premaxilla with lingual face of pars dorsalis lacking facet and dorsally expanded lateral internal strut for contact with nasal and from A. gracilis n. sp. in having maxilla with anterior end of tooth row lying well anterior to leading edge of nasal process.
DESCRIPTION
Albanerpeton galaktion is best known from the Milk River Formation and my descriptions below rely largely on specimens from this unit in the collection of the UALVP. Of the 39 catalogued specimens listed by Fox & Naylor (1982: 121) for A. galaktion , I retain the holotype premaxilla and 15 referred specimens (UALVP 16204, 16205, 16210-16213, premaxillae; 16240, 16241, maxillae; UALVP 16217-16219, 16221, 16222, 16236, dentaries; UALVP 16216, frontals) in the species. Other catalogued jaws listed by Fox & Naylor (1982) belong to A. nexuosus (see previous account), whereas parietals and atlantes from the formation cannot reliably be assigned to either species and, accordingly, are considered under the account for “Genus and Species indeterminate.” Other jaws and frontals from the Milk River Formation and eight premaxillae from Judithian and Lancian units are also referrable to A. galaktion . Descriptions below are composites, unless stated otherwise.
Premaxilla ( Fig. 4 View FIG ; Tables 2; 3)
Twenty premaxillae are available. The two most nearly complete specimens are the holotype UALVP 16203 ( Fig. 4A, B View FIG ) and UALVP 16204 ( Fig. 4C, D View FIG ). The latter is also the largest specimen, with a height of nearly 3.4 mm. Although the largest available premaxillae of A. galaktion and A. nexuosus are comparable in size, in the former species the bone is more gracile in build, the pars dorsalis is relatively taller and narrower ( Table 3), and the dorsal end of the pars dorsalis is smoother, indicating that the process was less strongly sutured in life with the nasal. All premaxillae at hand for A. galaktion are isolated and none shows evidence of having been fused medially in life with its opposite. The medial flange on the holotype is medially broad and dorsoventrally short, being restricted along the medial edge of the bone to the upper one-half of the pars dentalis and basalmost part of the pars dorsalis; however, on many referred premaxillae the flange is narrower and extends further dorsally up the medial edge of the pars dorsalis. The lacrimal notch typically is deep, but varies considerably in width ( Tables 2; 3). Labially, the upper one-quarter to one-third of the pars dorsalis bears an indistinct boss, best developed on UALVP 16204 ( Fig. 4C View FIG ) and RTMP 95.177.81 ( Fig. 4E View FIG ), that is weakly ornamented with low, anastomosing ridges and shallow, irregular pits. The remainder of the labial surface is smooth, except for scattered, small external nutritive foramina. Although not visible in Figure 4E View FIG , specimens such as RTMP 95.177.81 show that some of the foramina perforate the pars dorsalis to open medially inside the suprapalatal pit.
The lingual face of the pars dorsalis is dominated by a cavernous suprapalatal pit that approaches the shape of a right-angled triangle in lingual outline ( Fig. 4B, D, G View FIG ). The suprapalatal pit is large in both absolute ( Table 2) and relative terms, occupying slightly more than one-fifth (0.21- 0.23; n = 4) of the lingual surface area of the pars dorsalis. This enlarged pit extends across the medial two-thirds to three-quarters of the pars dorsalis and is located low on the process, with the ventral floor of the pit formed by the dorsal surface of the pars palatinum. Specimens broken across the pars dorsalis reveal that the tooth pulp cavities open dorsally into the floor of the pit. The medial and lateral walls of the suprapalatal pit are each formed by a mediolaterally narrow, but lingually deep and prominent internal strut. Each strut arises adjacent to the dorsal margin of the suprapalatal pit and expands lingually as it extends down the inner face of the pars dorsalis. The base of each strut extends about one-half of the distance lingually across the dorsal face of the pars palatinum. The lateral face of the more lateral strut is perforated by up to ten tiny foramina ( Fig. 4F View FIG ) that open inside the suprapalatal pit.
Preserved intact on UALVP 16212 ( Fig. 4G, H View FIG ) and RTMP 96.78.124 ( Fig. 4I View FIG ), the pars palatinum is lingually broad and bears prominent palatine and maxillary processes, both of which are indented lingually by shallow facets for contact with one or more palatal bones. The unnamed dorsal and ventral processes adjacent to the lingual edge of the maxillary process are each weakly developed – the former is a low ridge, isolated from the lateral internal strut, and the latter is a low, drumlin-shaped knob. The palatal foramen is remarkably large in absolute and relative terms, with a diameter about one and one-third to two times greater than the diameter of the bases of the medial teeth. The palatal foramen opens dorsally into the floor of the suprapalatal pit and ventrally about one-half the distance lingually across the pars palatinum, in line with the second to fourth loci. The canal connecting the dorsal and ventral openings of the foramen extends dorsoventrally through the pars palatinum. A tiny, unnamed foramen typically penetrates the bone ventrally at the junction between the pars palatinum and pars dentalis to open dorsally in the floor of the suprapalatal pit. In most premaxillae, this unnamed foramen is lateral to the palatal foramen, but in two specimens (UALVP 16210 and 39932, both unfigured) the unnamed foramen is medial to the palatal foramen.
Maxilla ( Fig. 3 View FIG I-N)
The two most nearly complete of the seven specimens at hand from the Milk River Formation are UALVP 16240 ( Fig. 3 View FIG I-K) and 16241 ( Fig. 3 View FIG L-N). These overlap in the region of about the second to fifteenth tooth positions, document essentially all of the structure of the element, and are from comparable-sized individuals. The former specimen is about 4 mm long as preserved and the bone was probably about 5 mm long when complete. A less nearly complete specimen, UALVP 39935 (not figured), was from an individual about one fifth again as large as those represented by UALVP 16240 and 16241. The labial surface is unsculpted, except for several small external nutritive foramina scattered across the anterior one-third of the bone. In labial view, the nasal process is triangular in outline and the pars facialis becomes shallower posteriorly. UALVP 16241 preserves most of the posterior end of the bone, which labially bears a shallow, anteroposteriorly elongate facet for articulation with the jugal. The ventral edge of the pars dentalis is straight to shallowly convex ventrally. The anterior end of the tooth row extends several loci anterior to the point of maximum emargination along the leading edge of the nasal process.
The premaxillary lateral process is longer than its height at the base and somewhat spatulate in labial or lingual outline, with the ventral edge strongly convex ventrally. The premaxillary dorsal process is a broad flange, with a low ventral ridge extending transversely across the posterior limit of the process. The pars palatinum is broad lingually, narrows towards the posterior end of the bone, and bears a raised patch dorsally for contact with the base of the lacrimal and a shallow trough dorsolingually for articulation with one or more unknown palatal bones. The internal narial margin spans seven or eight tooth positions.
Dentary ( Fig. 2 View FIG J-M)
Twelve incomplete dentaries from the Milk River Formation can be referred to the species. The two most nearly complete specimens, UALVP 16221 ( Fig. 2J View FIG ) and 16217 ( Fig. 2 View FIG K- M), are each broken posteriorly between the anterior edge of the opening for the Meckelian canal and the posterior end of the tooth row. UALVP 16221 is from a small individual and exhibits features typical of dentaries from smaller individuals, such as a more gracile build, a poorly developed ventral scar for attachment of the intermandibularis muscles, and a relatively low dental parapet that extends only up the ventral one-half of the tooth pedicels. UALVP 16217 is from a larger individual and the bone would have been nearly 10 mm long when complete. Although comparable in size to the largest available dentaries of Albanerpeton nexuosus, UALVP 16217 is less robust in build. Each of the referred dentaries of A. galaktion is unornamented and bears a short row of external nutritive foramina along about the anterior one-third of the bone. The dorsal edge of the dental parapet is straight in labial outline. The presence or absence of a dorsal process behind the tooth row cannot be determined, because no specimen at hand preserves this part of the bone. The symphyseal eminence is prominently developed, particularly on large dentaries. Nine specimens preserve intact symphyseal prongs and show that one or two prongs occur with about equal frequencies on either the left or right dentary. The remain- der of the lingual structure of the dentary is unremarkable.
Dentition ( Figs 2 View FIG J-M; 3I, J, L, M; 4)
Marginal teeth exhibit the characteristic albanerpetontid pattern of attachment and construction: highly pleurodont, non-pedicellate, with labiolingually compressed, chisel-like, and faintly tricuspid crowns. Of the eight premaxillae with a complete tooth row, six have eight tooth positions and one each has seven and ten positions. The maxilla with the most nearly complete tooth row, UALVP 16241 ( Fig. 3M View FIG ), preserves the posterior 21 loci and likely had no more than 25 tooth positions when complete. The two figured dentaries each have a nearly complete tooth row: UALVP 16217 preserves the first 26 tooth positions and probably had a further four to six teeth when complete, whereas UALVP 16221 retains 17 intact teeth and spaces for about 12-14 more teeth. Maxillary and dentary teeth are weakly heterodont in size anteriorly, with the longest teeth only slightly longer than nearby teeth. The longest teeth occupy about the fifth position on the maxilla and the sixth to tenth positions on the dentary. The relative sizes of teeth at the eighth and ninth positions on UALVP 16217 ( Fig. 2L View FIG ) are exaggerated by the presence of shorter replacement teeth at adjacent tooth positions. Most jaws have at least one empty tooth slot and one premaxilla (RTMP 96.78.124; Fig. 4I View FIG ) has a nearly functional replacement tooth at the fourth locus from the medial edge.
Frontals ( Fig. 6 View FIG F-K)
Nine azygous pairs of frontals are available from the Milk River Formation. UALVP 16216 ( Fig. 6F, G View FIG ), the most nearly complete specimen, is missing most of the anterior end (contra McGowan 1998), including the internasal and anterolateral processes, and the posterior end of both ventrolateral crests. This specimen is about 3.8 mm long as preserved and the bone was probably about 4.4 mm long in midline length when complete. UALVP 39946 ( Fig. 6H View FIG ) consists of the anterior part of a pair of fused frontals, broken obliquely between the slots for receipt of the prefrontals, and was from an individual about 1.5 times larger than that represent- ed by UALVP 16216. Frontals are solidly fused along the midline, regardless of size. UALVP 16216 shows that the fused frontals are triangular in outline and moderately elongate, with ratio of midline length to width across posterior edge between lateral edges of ventrolateral crests about 1.12, as preserved, and probably no more than about 1.2 when the bone was complete. UALVP 39946 demonstrates that the internasal and anterolateral processes and the more anterior slot for receipt of the nasal are well developed. The internasal process is acute and relatively broad (i.e. length subequal to width across the base) in dorsal outline and laterally has the groove for contact with the nasal. UALVP 16216, 39945 ( Fig. 6I, J View FIG ), and 39951 ( Fig. 6K View FIG ) collectively show that the lateral edge of the bone behind the anterolateral process diverges at about 20-25° from the midline, the dorsal and ventral margins of the more posterior slot for receipt of the prefrontal are emarginated medially, the orbital margin is shallowly concave, and the posterior edge of the frontal roof is shallowly concave to either side of the midline.
Frontals dorsally bear the usual albanerpetontid ornament of broad, polygonal pits enclosed by low ridges. As in Albanerpeton nexuosus , this ornament varies independent of overall frontal size and is more prominent on some specimens than others (cf. Fig. 6F, H, I View FIG ).
The ventrolateral crest is moderately wide and triangular in transverse view. The crest widens with growth: width of crest behind slot for receipt of prefrontal increases from 0.65-0.71 mm and ratio of crest width to width across posterior edge of frontals between medial face of crests increases from 0.25-0.40 (n = 3). The transverse profile of the crest also changes with growth as the bevelled ventral face broadens, assumes a more ventrolateral orientation, and becomes flatter or, at most, shallowly concave on the largest specimens (cf. Fig. 6G, J, K View FIG , respectively). The remainder of the ventral structure of the frontals is unremarkable, with one notable exception. UALVP 16216 is unique among albanerpetontid frontals in having one large foramen opening at the midline between the anterior ends of the ventrolateral crests. This anomalous condition differs from the typical albanerpetontid pattern (see Fig. 8C View FIG ), in which a smaller foramen opens to either side of the midline in the ventral face of the anterior part of the ventrolateral crest (Gardner 1999d).
REMARKS
In addition to the diagnostic frontal characters, assignment of Albanerpeton galaktion to Albanerpeton is further supported by the triangular lingual outline of the suprapalatal pit. A triangular to slit-shaped suprapalatal pit is unique among albanerpetontids to A. galaktion , A. cifellii , and A. gracilis n. sp. and allies these species in the less inclusive gracile-snouted clade (Gardner 1999b, c).
Fox & Naylor (1982: 121) diagnosed A. galaktion using five features of the jaws. Their first feature – “deep, large pit behind pars facialis” – refers to the enlarged suprapalatal pit; this feature occurs in all premaxillae that I attribute to A. galaktion and is reliably diagnostic for the species (see below). None of the other four features are as diagnostically useful. Lack of labial ornament on the more ventral part of the premaxillae is a symplesiomorphy of albanerpetontids that differentiates A. galaktion only from the two Tertiary congeners, both of which have ornament covering the labial face of the pars dorsalis in large individuals. Fox & Naylor’s (1982: 121) statement that A. galaktion is diagnosed by “enlarged teeth in anterior part of dentary and maxillary” is based on specimens from the Milk River Formation that I attribute to A. nexuosus . Maxillae and dentaries that I refer to A. galaktion have teeth that are weakly heterodont in size anteriorly. The latter pattern is not particularly diagnostic, because it is primitive for the genus and differentiates A. galaktion only from A. nexuosus . Neither a short medial flange nor a prominent lacrimal notch (= “medial ridge” and “indentation for prefrontal”, respectively, of Fox & Naylor 1982) are diagnostically useful for albanerpetontids, because the proportions of both structures vary more within and overlap more among species than Fox & Naylor (1982) suspected (see also Gardner 1999a, 2000a).
McGowan (1998) recently presented a revised diagnosis for A. galaktion based on four frontal characters. In formulating his diagnosis, McGowan (1998) relied entirely on photographs (Fox & Naylor 1982: fig. 1d, e) of the referred frontals UALVP 16216 ( Fig. 6F, G View FIG ). McGowan (1998) believed UALVP 16216 was relatively complete, at least anteriorly. In fact, the specimen is broken between the slots for receipt of the prefrontals (= “lacrimal facets” of McGowan 1998) and lacks about the anterior one-quarter of the bone. UALVP 16216 thus provides no information about the form of the internasal process (= “anterior process” of McGowan 1998). The surface that McGowan (1998) regarded as the outline of the internasal process is actually the broken leading edge of the specimen (Gardner 2000a). UALVP 39946 ( Fig. 6H View FIG ) shows that the internasal process is acute in outline, with the length and width subequal – not short, broad, and wedge-shaped as depicted in McGowan’s (1998: fig. 1B) reconstruction. In lacking the anterior part of the bone, UALVP 16216 also underestimates the relative length of the orbital margin. When the bone was complete, the anterior end of the orbital margin would have been about in line with the anteroposterior midpoint of the bone, not anterior to this point as McGowan (1998) claimed; the former condition occurs in all species of Albanerpeton and is diagnostic for the genus (Gardner 2000a). Contrary to McGowan (1998), emargination of the slot for receipt of the prefrontal is not useful for differentiating species of Albanerpeton because there is no substantial interspecific variation in the degree of emargination among large individuals (Gardner 2000a). Emargination of the slot increases with growth (see Gardner 1999d: pl. 2, G-L) and this alone accounts for the shallowly excavated slots in UALVP 16216. McGowan (1998) correctly observed that the orbital margins in UALVP 16216 are less divergent than in frontals of A. inexpectatum . The former condition is a function of the azygous frontals being relatively more elongate and resembling an isosceles triangle in outline. This condition is primitive for the genus and differentiates A. galaktion only from A. inexpectatum , a species in which the frontals are relatively broader and more nearly resemble an equilateral triangle.
Albanerpeton galaktion is convincingly diagnosed by two premaxillary apomorphies that are unique among albanerpetontids: 1) suprapalatal pit occupies about one-fifth to one-quarter of lingual surface area of pars dorsalis (suprapalatal pit smaller in other albanerpetontids: accounts for about 1% of surface area of pars dorsalis in non- Albanerpeton albanerpetontids and A. arthridion , and 4 to 15% of pars dorsalis in other species of Albanerpeton ); and 2) palatal foramen large, with diameter about one and one-third to two times greater than bases of medial teeth on premaxilla (diameter of foramen primitively subequal to and generally smaller than that of medial teeth). Given that the suprapalatal pit and palatal foramen are intimately associated in albanerpetontids, enlargement of these bony openings in A. galaktion is probably linked. Soft structures associated with these openings probably were involved with feeding or olfaction (Gardner 1999b, 2000a) and can be expected to have been enlarged in size and, presumably, enhanced in function in A. galaktion relative to other albanerpetontids.




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Albanerpeton galaktion Fox & Naylor, 1982
| Gardner, JD 2000 |
Albanerpeton galaktion
| Fox & Naylor 1982: 121 - 127 |
Albanerpeton nexuosus
| Estes 1981: 24 |