
Aulosaccus schulzei Ijima, 1896
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3628.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:37D2D7F2-FA0C-40E9-B6D0-9C74EBB6C7F0 |
|
DOI |
https://doi.org/10.5281/zenodo.5261626 |
|
persistent identifier |
https://treatment.plazi.org/id/03D287B2-FFA4-3609-9AD7-FCCD2F61FEE7 |
|
treatment provided by |
Felipe |
|
scientific name |
Aulosaccus schulzei Ijima, 1896 |
| status |
|
Aulosaccus schulzei Ijima, 1896 View in CoL
( Figs. 14 View FIGURE 14 & 15 View FIGURE 15 , Table 7)
Synonymy. Aulosaccus schulzei Ijima, 1896: 252 ; 1898: 51; 1904: 110; Koltun 1967: 68; Tabachnick 2002: 1447; Lee et al. 2007: K257; Stone et al. 2011: 33.
Aulosaccus albatrossi Okada, 1932:78 View in CoL ; Koltun 1967: 68.
Material examined. USNM# 1196554 View Materials , ROV ' Jason II' from RV ' Roger Revelle', dive J2105, 06 August 2004, 4.5 km W of Cape Sajaka, Tanaga Island , Aleutian Islands , Alaska, 51º52.021'N, 178º17.720'W, 1320 m, 1 partial specimen attached to cobble, dry & ethanol GoogleMaps .
Comparative material examined. A. schulzei holotype, MCZ 10004, Sagami Bay, Japan, prior to 1896, dry. A. albatrossi holotype, USNM 22111, USFS 'Albatross' stn 4797, 20 June 1906, Staritschkof Island off South Kamchatka Peninsula, 52˚37.8'N, 158˚49.8'E, 1248 m, ethanol
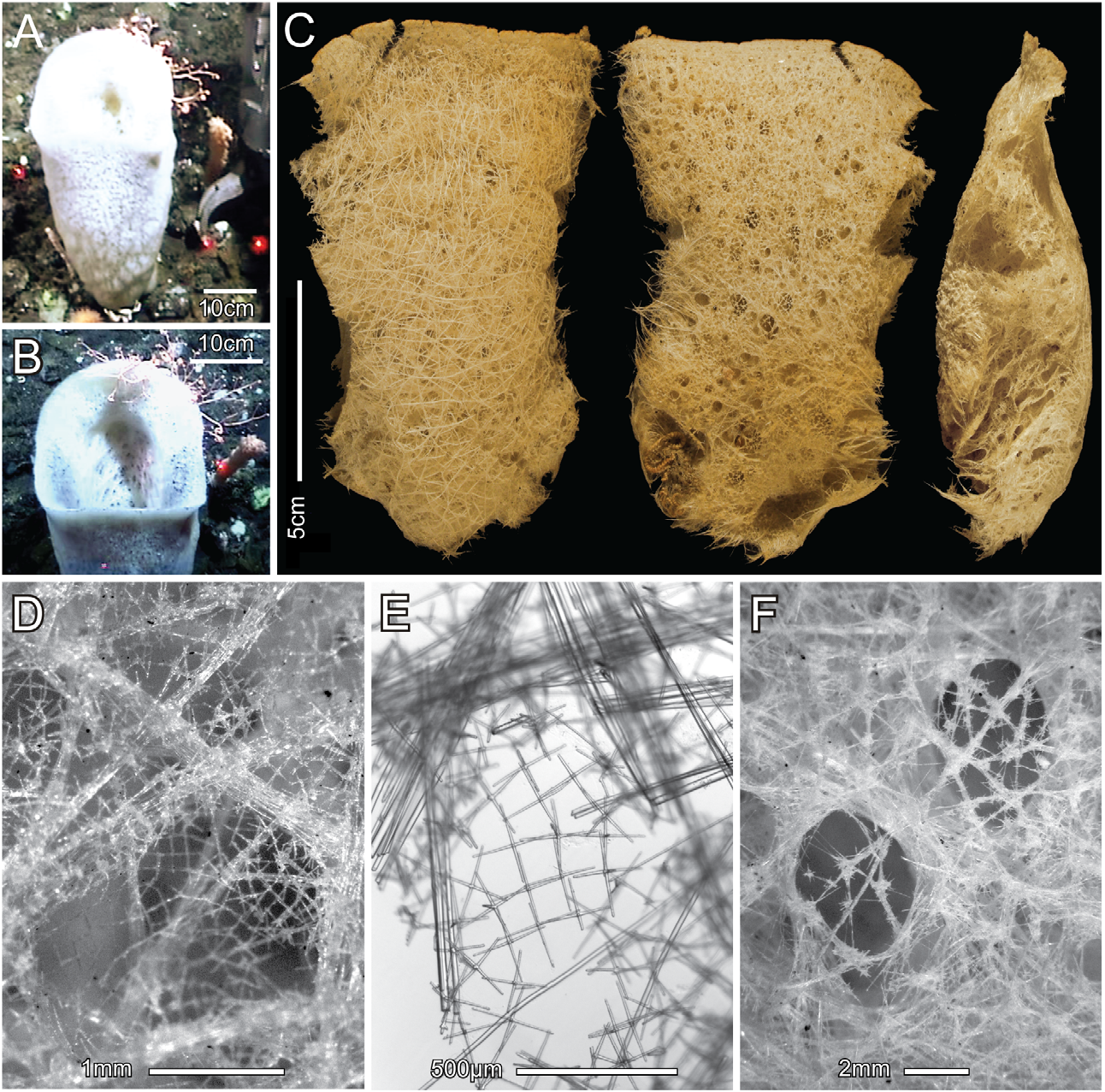
Description. The collected fragment is the top quarter of a slightly flattened sac, 53 cm long by 25 cm diameter, with large flared oscular margin ( Figs. 14A, B View FIGURE 14 ); dimensions of the fragment ( Fig. 14C View FIGURE 14 ) are 135 mm tall, 110 mm wide, and 34 mm in wall thickness in the middle. All surfaces are smooth, without prostalia, veil, or conules. The oscular margin is thin and without marginalia. The dermal surface has visible strands of hypodermal pentactins and diactins, oriented in all directions, forming a subsurface network supporting the dermal spicule lattice and obscuring the apertures of the subdermal inhalant canals ( Figs. 14C, D View FIGURE 14 ). The dermal lattice ( Figs. 14D, E View FIGURE 14 ) is a rectangular meshwork with mean side length of 122 µm, composed of about equal numbers of hexactins, pentactins, and stauractins. The atrial surface has openings of exhalant canals generally visible through the less regularly organized atrial lattice ( Fig. 14F View FIGURE 14 ). Meshes of the atrial lattice, composed mainly of hexactins with a few stauractins and pentactins, are not rectangular and microscleres, not present in the dermal lattice, make up a significant component of this structure. The main supporting skeleton is a network of loose interwoven unfused diactins occurring in bundles or singly and oriented without order. Spicule fusion probably occurs only in the basidictyonal skeleton, not collected. Within the moderately firm surface layers, the main parenchyma is cavernous, resulting in a very soft overall body texture. Color of the fresh specimen was white; when preserved in ethanol or dried it is light brown.
Megascleres: (spicule dimensions are given in Table 7). Hypodermal pentactins ( Fig. 15A View FIGURE 15 ), and a very few triactins and irregular tetractins, are mostly crucial, rarely semi-paratropal, and smooth, with long tapered rays ending in rough sharp tips. Their tangential rays are bundled with thick and thin diactins to form primary support for hexactine dermalia. Dermalia ( Fig. 15B View FIGURE 15 ) are entirely rough spicules with cylindric rays ending in rounded and sometimes inflated tips; 37% are hexactins, 31% pentactins, and 32% stauractins. Hexactins are distributed mainly over the hypodermal strands, pentactins at edges of strands, and stauractins over the unsupported middle of the larger lattice fields. Atrialia are composed of the same three spicule types ( Fig. 15C View FIGURE 15 ), but they are larger than dermalia, tend to have elongate proximal rays in pentactins and one axis elongate in hexactins. Parenchymal megascleres are all diactins, including thick principal diactins ( Fig. 15D View FIGURE 15 ) and thin intermediate and comital diactins ( Fig. 15E View FIGURE 15 ). Thick principal diactins are slightly curved, generally smooth, with no medial swelling and rough rounded or parabolic tips. Thin diactins are straight to sinuous, generally smooth with a significant medial swelling and rough parabolic tips. Short straight diactins ( Fig. 15F View FIGURE 15 ) are associated with only the atrial surfaces.
µm unless otherwise indicated).
Microscleres consist of immense discasters, also known as solasters, hemioxyhexasters, oxyhexactins and variants, and microdiscohexasters. The subdermal solasters ( Fig. 15G View FIGURE 15 ) are derived discohexasters with centrum expanded to a solid sphere, entirely enclosing the primary rays. Terminal rays emanating directly from the centrum are unequal in length, ornamented with short reclined barbs, and end in marginally toothed discs. Oxy-tipped microscleres ( Fig. 15H View FIGURE 15 ) are very thin and mainly associated with the atrial skeleton; they include regular oxyhexactins, hemioxyhexasters and an array of forms with reduced ray numbers—2, 3, 4, 5 developed rays—but never complete oxyhexasters. They are all rough, all terminal rays ornamented with reclined spines. Many have sharp bends at the end of the primary rays, with one or two developed rays and a short spur emanating from that point; compact spiral variants are rare. Microdiscohexasters ( Fig. 15I View FIGURE 15 ) are very rare and associated with both surface skeletons. They have short, thick, smooth primary rays, each of which supports 5–10 spined, irregularly sinuous terminal rays ending in very small discs with recurved marginal teeth.
Remarks. This species is widely distributed in the N Pacific, from Sagami Bay, Japan through the Kurile Islands and the Okhotsk Sea (Tabachnick, 2002) and has recently been reported from Central California ( Lee et al. 2007). Our discovery of A. schulzei in Alaska is not surprising, but it represents a considerable range extension of ca 2,000 km from its nearest previously reported occurrence in the Okhotsk Sea. The new Aleutian Islands specimen agrees in virtually all details with the holotype specimen from Sagami Bay, Japan. In reviewing other possible assignments of this specimen, we discovered that there are no significant differences between A. albatrossi Okada, 1932 from off S Kamchatka Peninsula, and A. schulzei . In his description of A. albatrossi, Okada described its macrodiscohexasters (not figured) as 250–650 µm in diameter (we found 409–592–788 µm, n = 8, in the holotype), terminal rays as smooth with conically convex discs (we found terminal rays rough with hemispheric discs), and two size classes of microdiscohexasters of 32–40 and 50–56 µm diameters (we found one class, 23–34–42 µm, n = 127). Okada's description of A. albatrossi is, like many of his other descriptions, clearly unreliable and almost all reputed differences between the species evaporate when the holotypes are directly compared. Maximum size of macrodiscohexasters (solasters) is quite variable between specimens of the same species and is not alone considered a sufficient basis for distinguishing between species. We thus propose A. albatrossi Okada to be recognized as a junior synonym of A. schulzei Ijima. The description here of the Aleutian Islands specimen is the first documentation of A. schulzei spicule forms with SEM.
Review of all video footage collected with the ROV 'Jason II' indicates that it is a rare species occurring singly on bedrock, boulders, and cobbles at depths between 1270 and 1350 m. Associated fauna include juvenile lithodid crabs ( Paralomis verrilli and Lithodes couesi ), pandalid shrimps, and the large ophiuroid Gorgonocephalus eucnemis .

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Aulosaccus schulzei Ijima, 1896
| Reiswig, Henry M. & Stone, Robert P. 2013 |
Aulosaccus albatrossi Okada, 1932:78
| Koltun, V. M. 1967: 68 |
| Okada, Y. 1932: 78 |