
Chironomus blaylocki, Wuelker, Wolfgang, Martin, Jon, Kiknadze, Iya I., Sublette, James E. & Michiels, Susanne, 2009
|
publication ID |
https://doi.org/ 10.5281/zenodo.186122 |
|
DOI |
https://doi.org/10.5281/zenodo.6215044 |
|
persistent identifier |
https://treatment.plazi.org/id/BD339627-FFDD-FF89-FF3E-632FFAE0BC81 |
|
treatment provided by |
Plazi |
|
scientific name |
Chironomus blaylocki |
| status |
sp. nov. |
Chironomus blaylocki View in CoL sp. n.
Chironomus tentans Fabricius, 1805 View in CoL ; Blaylock et al. 1964; Blaylock 1965, 1966a, 1966b: karyotype and chromosomal polymorphism (misidentification); belongs to thummi-cytocomplex ( Wuelker et al. 1968).
Chironomus View in CoL “ decorus View in CoL . B” Martin 1979: chromosome arm F
Chironomus “ tentans Blaylock ” Wuelker et al. 1989: karyotype
Chironomus “ tentans Blaylock ” Wuelker et al. 1991: karyotype
Chironomus species l Martin 2008: karyotype, and associated larva.
Chironomus blaylocki Kiknadze et al. 2004 View in CoL : banding pattern of 5 chromosome arms (nomen nudum)
Type material. Holotype, U.S.A., White Oak Creek, nr. Oak Ridge, Anderson Co., Tennessee, Sgc, 31.I.1964, " C. tentans ", slide 3, leg. BGB, in ZSM.
Other material examined. We are in possession of only 10 salivary gland chromosome squashes of this species (coll. BGB), all from Tennessee, of which only one has the larval body on the same slide: White Oak Creek, 6 chromosome squashes as Holotype but various dates (VI. 1961 20.IX.1962 and I.1964), 4 with larval body (22.VI.1967) (location of 3 of these, partially measured, is now uncertain but likely in ZSM or UMN); Ten Mile Creek, nr. Knoxville, Knox Co. 2 SGCs (8.XI and 18.XII.1962); Mile 37.5, Mc Coy Branch, Clinch River, Knox Co. 2 SGCs (20.XI. and 17.XII.1962). There are a few male adults and larvae from White Oak Creek at the same date as the larval samples, but not clearly associated. The remark of Blaylock et al. (1964) that “7 different Chironomus species occur in Ten Mile Creek” is a warning against unproved association, particularly since other species, one of which has a larva similar in gross morphology to that of C. blaylocki ( Spies et al. 2002) , have been obtained from White Oak Creek. However, there are also some measurements of three associated larvae, and a male collected at the same place and time as these larvae. This male, while problematically associated, shows overall similarity to the adults associated by rearing of C. bifurcatus , whose larval material constitutes a closely related cytomorphological species pair, makes the description appropriate.
Other species recorded from the locality would not be expected to show such similarity. However, associated reared material will be necessary to confirm the adult morphology.
Etymology. Elected by J.E. Sublette as a manuscript name, after B.G. Blaylock.
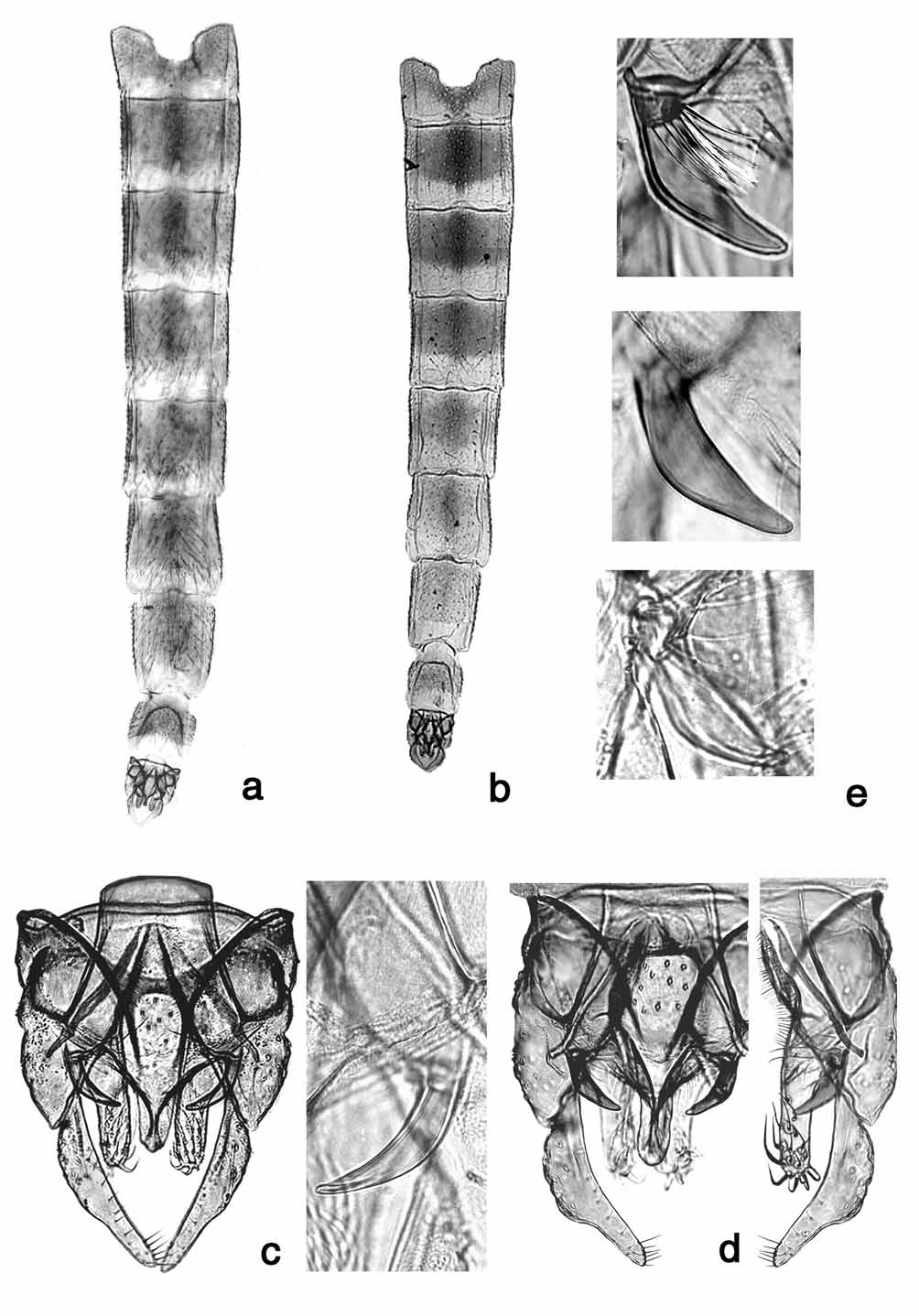
Diagnostic characters. This species is presently most accurately identified by the unique banding pattern of the polytene chromosomes. There are unique patterns in three of the seven chromosome arms. Most of the shared patterns (arms B, C2, E and F) are with species b ( Martin 2008), which may be a subspecies of C. blaylocki (see Discussion). There is a shared pattern of arm C with C. bifurcatus and in arms E and F1 with C. decorus (Martin 1979) and C. decorus R & F ( Rothfels & Fairlie 1957). Due to the presence of polymorphic sequences, only the pattern blaA1 (1a–e, 8–9, 2d–3e, 15–14, 2c–1f, 16a–d, 7–4, 13a–f, 10– 12, 3i –f, 17–19) and the characteristic arm G can be expected in all specimens.
The presumptive adult male (see below) has genitalia ( Fig. 3 View FIGURE 3 c) very similar to those of C. bifurcatus but the superior volsella on C. blaylocki , while dark as C. maturus Johannsen, 1908 and C. bifurcatus , is more smoothly curved but slightly angled as in both of those species and is almost evenly tapered from the base to the tip. The phallopodeme is distinctly different from those species, resembling that of Chironomus atrella (Townes, 1945) .
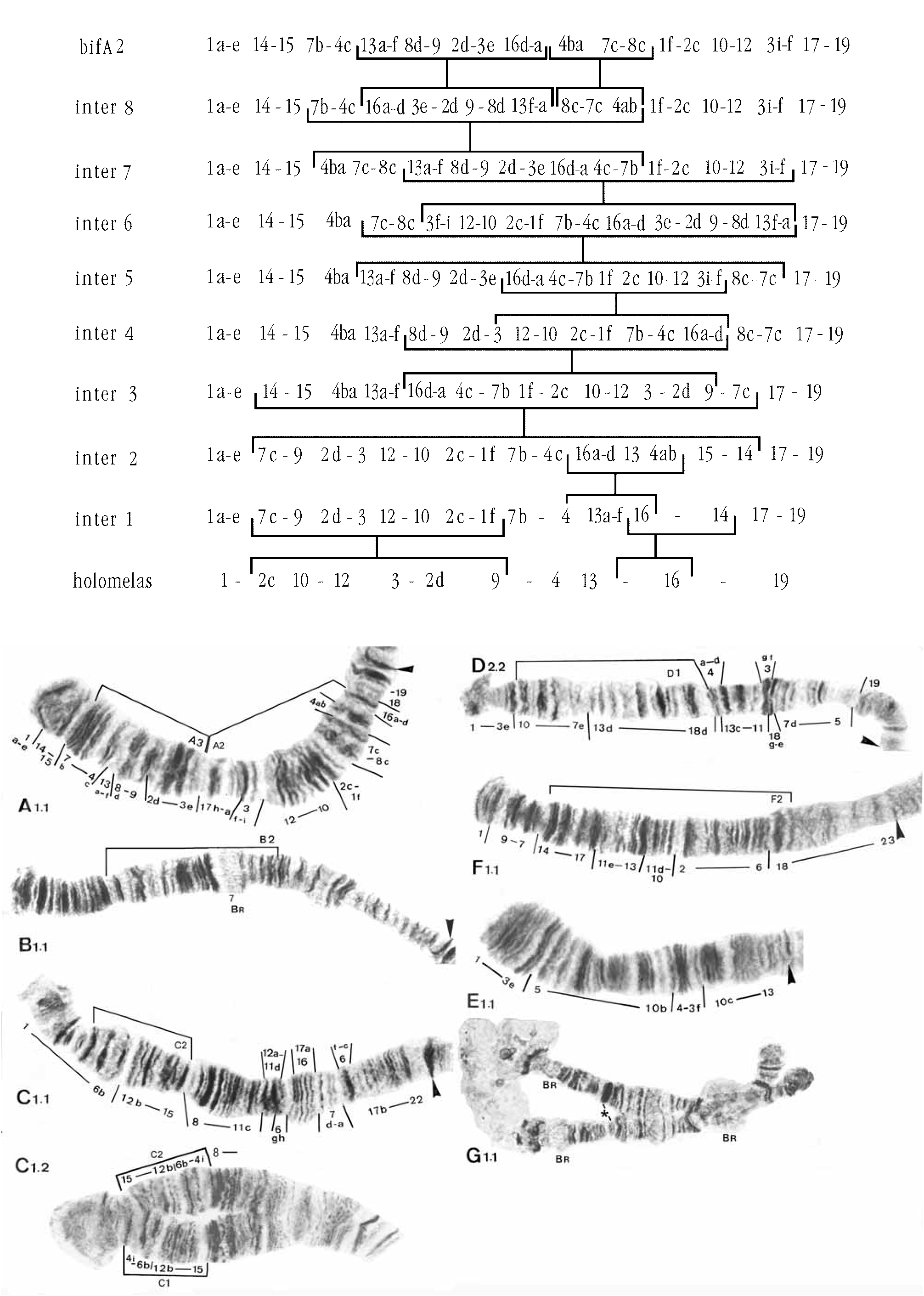
Karyotype ( Fig. 1 View FIGURE 1 )
Chromosome arm combination AB, CD, EF, G (thummi-cytocomplex). Nucleolus subterminal in arm G, no nucleolus in the long chromosome arms. Three BRs are normally visible, two in arm G (one just distal of the nucleolus, the other near the other end of the chromosome) and one in arm B (normally near middle of the arm, but may be distal due to inversion polymorphism).
Inversion polymorphism is known in chromosome arms B, C, D and F. Some polymorphism was described by Blaylock et al. (1964) using different names. The relationship to the identification used here is given in Table 1.
This paper Blaylock et al. 1964 blaA1 2R
blaB1 2L
blaB2 2Lab blaC1 1R
blaC2 1Ra
blaD1 1L
blaD2 1Lc
blaE1 3L
blaF1 3R
blaF2 3Ra
Arm A
There is only one banding pattern known in arm A, blaA1, which can be derived from arm A of Chironomus harpi Sublette, 1991 (Wuelker et al. 1991) another member of the decorus-group, by two simple inversion steps. In3f–16, leads to a hypothetical intermediate, then In13–2c leads to blaA1. Arm A of C. utahensis , also in the decorus-group, is a further simple inversion step from C. harpi (see below).
Arm B
B1 with a BR in the middle of the arm, followed distally by a group of dark bands. These appear to correspond to band groups 7 and 8 respectively, in the standard system of Dévai et al. (1989). Blaylock et al. (1964) report a complex inversion in this arm, which they call 2Lab. They considered 2Lab to be composed of two independent components that were always found together. However, our interpretation of this heterozygote, here called B1.2 ( Table 1), is that it is due to overlapping inversions, similar to F1.2 of C. utahensis (Wuelker et al. 1991) . The limits of these inversions are shown in Fig. 1 View FIGURE 1 . The longer of these inversions takes the BR to near the distal end of the arm with the dark bands proximal to it ( Fig. 4 View FIGURE 4 a).
Arm C
Two banding patterns have been recorded. Although less frequent, blaC1 appears phylogenetically more basal, as it is only a simple inversion step from the common pattern recorded in a number of species of Chironomus (Wuelker 1991) . C2 differs from C1 by a small simple inversion, 2f–6.
There is an additional polymorphism of this chromosome, in which the centromere band is puffed. In the heterozygote the puffing may disrupt pairing in the vicinity, leading to Blaylock et al. (1964) describing it as an inversion (1Rb). The arm C in Fig. 1 View FIGURE 1 is homozygous for the puffed band.
Arm D
Of the two patterns reported for this arm by Blaylock et al. (1964), we have only seen one. There are two equally parsimonious explanations for the derivation of the more frequent pattern D1. It can be derived from the common pattern of Wuelker (e.g. 1991) by 4 intermediate patterns involving 6 simple inversions. Note that the step from inter 3 to inter 4 in this scheme would restore group 18 to its original state, which in inter 2 is 18g –a.
Equally, it may be derived from the pattern of Chironomus piger Strenzke, 1951 by a similar number of steps:
The second sequence, D2, on the basis of the description and photograph in Blaylock et al. (1964), is derived from D1 by a simple inversion of about 11–19g.
Arm E
Only one pattern, blaE1, has been found in this arm. It differs from the basic pattern for this arm as found in Chironomus aberratus Keyl, 1961 ( Wuelker 1980) by the simple inversion 5–8.
The E1 pattern is also found in other decorus-group species, such as C. decorus (R&F) and C. species 3a (Martin et al. 1979), as well as in C. maturus (Wuelker & Martin 1974) and C. stigmaterus Say, 1823 (Martin & Wuelker 1974) .
Arm F
Two patterns have been reported in arm F. The only homozygote represented in our sample is blaF2.2. This pattern differs from blaF1 by the simple inversion 5c–16, while F1 differs from the standard (basic) pattern of C. piger by the characteristic inversion of the decorus-group, 2–9.
The pattern blaF1 is also found in C. decorus (Martin et al. 1979) .
Arm G
Long, stretched, and paired, according to Blaylock et al. (1964), but with pairing gaps in our material. One BR near one end, a subterminal nucleolus at the other end and another BR distal of the nucleolus.

| ZSM |
Bavarian State Collection of Zoology |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Chironomus blaylocki
| Wuelker, Wolfgang, Martin, Jon, Kiknadze, Iya I., Sublette, James E. & Michiels, Susanne 2009 |
Chironomus blaylocki
| Kiknadze et al. 2004 |
Chironomus tentans
| Fabricius 1805 |