
Allodrilus Evangelista
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3847.4.2 |
|
publication LSID |
lsid:zoobank.org:pub:46E41A86-A877-4690-80D9-1C91149A4F8E |
|
DOI |
https://doi.org/10.5281/zenodo.6124148 |
|
persistent identifier |
https://treatment.plazi.org/id/C0248799-FFFE-687B-FF38-CF936166FBE4 |
|
treatment provided by |
Plazi |
|
scientific name |
Allodrilus Evangelista |
| status |
gen. nov. |
Allodrilus Evangelista gen. nov.
Type-species: Cymbomorpha nitidipennis Funkhouser, 1922a: 14 .
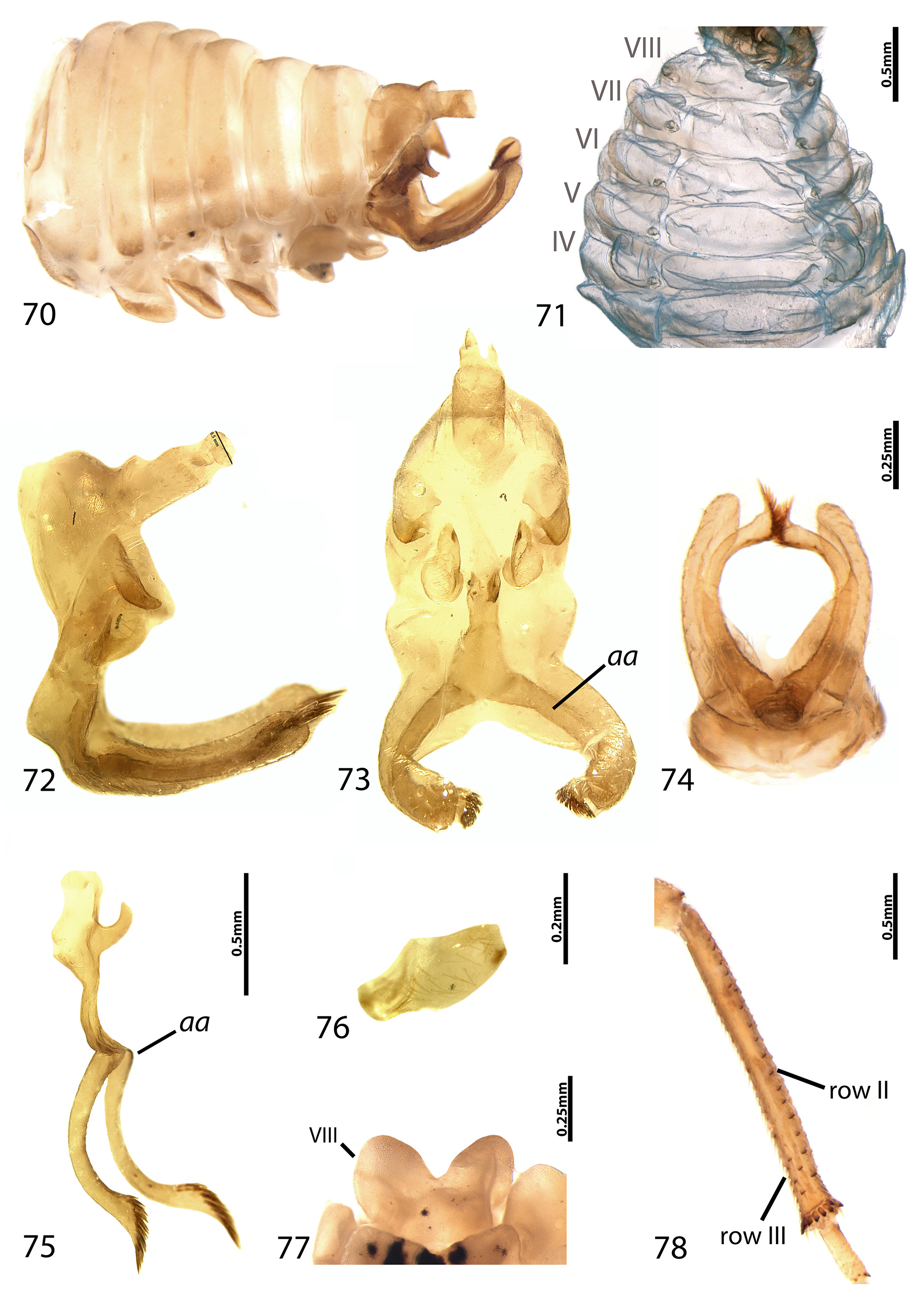
Diagnosis. Vertex moderate to distinctly concave below ocelli; pronotum low to slightly elevated, convex dorsally in lateral view, lacking suprahumeral horns, tubercles and spines; in dorsal view, anteriorly round at metopidium, widest between humeral angles, posterior process triangular, tapering towards acute apex in dorsal view; forewings with initial division of vein R as follows: R1+R s A[R2+3] and R s P[R4+5] (i.e., R1 diverging after first bifurcation of R), crossveins s, r-m and m-cu 2 present (one each); row III of hind tibia with cucullate setae smaller in comparison to row I and row II, present only along apical half or third; males with elongate, cylindrical, and bifurcate aedeagus apophysis that extends along subgenital plate, curved and sharp distally, apex covered in comb of hairs.
Description. Color: Head, pronotum and legs brown, ferruginous, and yellow, or variegated with a combination of these colors; longitudinal bands occasionally present. Fore- and hind wings mostly hyaline. Forewings with occasional dark macula; coriaceous areas opaque, usually with same colors as pronotum.
Head: triangular, dense and roughly punctate; eyes and ocelli well produced, eyes globose; vertex pentagonal, superior margin arched, area below ocelli moderate to distinctly concave, pressed down in comparison to rest of vertex; coronal suture distinct, not conspicuously grooved; ocelli located approximately on the imaginary line that crosses center of eyes, distance between ocelli and eyes variable; superantenal ledges broad, sharply carinate, foliaceous; lower margins of vertex encompassing supra-antennal ledges and frontoclypeus curved forwards; frontoclypeus concave, diamond-shaped, about as wide as long, exceeding lower margins of vertex.
Pronotum: low, convex dorsally in lateral view, lacking suprahumeral horns, tubercles and spines; in dorsal view, anteriorly round at metopidium, widest between humeral angles, posterior process triangular, tapering towards acute apex in dorsal view; surface roughly punctate, pits smaller and more numerous near base of metopidium, slightly enlarged towards apex of posterior process; metopidium curved above suprahumeral angles in lateral view, dorsum straight to broadly arched; posterior process triangular, gradually tapering to acute apex.
Wings: forewings with coriaceous, densely punctate areas near base; veins prominent, with discreet adjacent punctation; R, M and Cu fused at base: R separating shortly after, M and Cu confluent for a short distance, diverging at basal third; initial division of R: R1+R s A[R2+3] and R s P[R4+5] (i.e., R1 branching out after first bifurcation of R); crossveins s, r-m and m-cu 2 present (one each); one discoidal cell; M1+2 and M3+4 conspicuously oblique; A1 well marked and elongate, A2 indistinct. Hind wings with one r-m and one m-cu, A1 and A2 confluent basally, then diverging at apical third; jugal lobe well developed, broadly round.
Legs: pro-, meso- and metathoracic legs without cucullate setae (except for hind tibia); tibia prismatic, not foliaceous; hind tibia with three rows of cucullate setae, row III with smaller setae in comparison to row I and row II, extending along apical half or third of tibia.
Abdomen: abdominal segments lacking conspicuous punctation, dorsal tuberosities or fenestrae. Males with lamellar laterotergites in segments IV–VII (not as distinctly lamellar in A. alboferrugineus sp. nov.), VIII sternite rectangular or heart-shaped; subgenital and lateral plate leaf-like, usually swollen, with or without spines and tubercles; aedeagus small in comparison to lateral plates and aedeagal apophysis, shaft cylindrical, U-shaped, with remarkably elongate, cylindrical apophysis bifurcate at or near base, extended along subgenital plates, apex acute, covered by comb of hairs; styles usually smaller than aedeagal shaft, hidden behind lateral plates, comma-shaped or rectangular, apex truncate with minute ventral lobe. Female genitalia according to subfamily description ( Deitz, 1975a: 148).
Distribution. GUYANA (Cuyuni-Mazaruni: Pakaraima Mountains [unknown municipality]). BRAZIL (Amapá: Serra do Navio; Amazonas: Itacoatiara, São Gabriel da Cachoeira, Santa Isabel do Rio Negro, Benjamin Constant; Pará: Santa Isabel do Pará, Óbidos, Belém, Santarém; Acre: Mâncio Lima; Rondônia: Ouro Preto do Oeste, Chupinguaia, Vilhena; Mato Grosso: Sinop). BOLIVIA (Santa Cruz: San Rafael). PERU (Loreto: Iquitos; Madre de Dios: Reserva Nacional Tambopata). COLOMBIA (Vaupés: Apaporis River [unknown municipality], Leticia). ECUADOR (Sucumbios: Shushufindi, Garza-cocha; Napo: Tena, Coca, Reserva Etica Waorani; Orellana: Parque Nacional Yasuni, Puerto Francisco de Orellana; Pastaza: Cerros de Abitagua).
Remarks. Species classified in Allodrilus gen. nov. closely resemble those in Smiliorachis , especially in their coloration, pronotal shape, and forewing venation. Despite their similarity, the male genitalia have considerable importance to genus and species recognition. In this newly proposed genus, the aedeagus shaft and styles are small, the latter often concealed by lateral plates. The aedeagus bears a remarkably elongate and bifid apophysis that extends along the subgenital plate, each branch curved upwards, and deflected distally into an acute hairy apex ( Fig. 75 View FIGURES 70 – 78 ). In most cases, the branches of aedeagal apophysis are partially enclosed by the subgenital plate, whose lobes are dorsally folded to accomodate them. The function of such an unusual and elaborate structure is unknown, but given reduced paramere size, the apophysis could serve to hold or assistist in holding the female during copula. In Allodrilus gen. nov., the vertex is medially concave to various degrees between species, showing slight intraspecific variation in A. nitidipennis comb. nov., and A. alboferrugineus sp. nov. This concavity encompasses the area just below the ocelli, suprantennal ledges and frontoclypeus, depressed anteriorly, and curved forwards posteriorly. The frontoclypeus is conspicuously scoop-shaped in most species (sensu Dietrich et al., 2001). The laterotergites of abdominal segments IV–VII are lamellar in males, and the VIII sternite is often emarginated, giving the impression that the VIII laterotergite is also lamellar ( Fig. 70–71 View FIGURES 70 – 78 , abdominal segments indicated). The chaetotaxy of the hind tibia in D. intectus —with setal row III reduced or absent—was previously discussed as an unusual feature that could suggest a phylogenetic relationship with membracines ( Dietrich et al., 2001a; Lin et al., 2004a). The misidentified specimen of D. intectus (not examined in this study) may belong to Smiliorachis or Allodrilus gen. nov. In the latter genus, the hind tibial setal row III is reduced in comparison to rows I and II (both in size and number of setae), with setae usually extending along the apical third of the tibiae. However, no absence of row III was observed in the examined specimens. A similar reduction of setal row III is seen in a few Smiliorachis species, for instance, in S. bracaatingae Sakakibara and Laroca. Nevertheless , this is can be a useful character to differentiate representatives of Allodrilus and Smiliorachis when examination of the male genitalia is not possible. Other aspects, such as the medially concave vertex, and lamellar laterotergites in males are also observed in Rhexia and some Smiliorachis , but are nonetheless important for recognition of this new genus. The biology of Allodrilus gen. nov. is mostly unknown, but examined specimens indicate they are tended by honeydew harvesting ants. Three species of ants in the genera Camponotus , Crematogaster , and Ectatomma were recorded in association with A. nitidipennis , and a number of specimens consistently collected in the same event suggest that this species exhibits some degree of aggregation of adults, nymphs or both.
Most representatives of Allodrilus gen. nov. can only be accurately determined after examination of their male genitalia, hence identification of females may depend on association with males from the same collecting event. Due to the small number of type specimens of certain newly proposed taxa, intra-specific variation in body coloration and pronotal shape could not be critically evaluated in all cases. Notwithstanding, nearly identical specimens can show deep divergences in mitochondrial loci (as exemplified by A. nitidipennis comb. nov., A. similis sp. nov., and A. intermedius sp. nov.; Evangelista et al., in preparation). An iterative approach revealed differences in head shape, pronotum, and male genitalia (especially in the shape of lateral plate), and these features were used to characterize new species based on morphology. Whenever possible, DNA vouchers were designated as primary types; sequences obtained from these specimens will be subsequently published. An integrative approach with a broader sampling of individuals would be necessary to illuminate species limits in such similar groups. This will help quantify the underestimated heteronotine diversity, as well as to test the classification proposed herein.
Etymology. The generic name ‘ Allodrilus ’ is derived from the Greek ‘ alloios ’ = different, strange, and ‘ drilos ’ = penis, and refers to the unusual morphology of the aedeagus.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.