
Dichrorampha rejectana (DE LA HARPE, 1858) Bryner & Huemer & Wittland, 2015
|
publication ID |
https://doi.org/ 10.5169/seals-787076 |
|
DOI |
https://doi.org/10.5281/zenodo.5851539 |
|
persistent identifier |
https://treatment.plazi.org/id/03D087DE-FF92-FFC6-3281-102025FB82A7 |
|
treatment provided by |
Carolina |
|
scientific name |
Dichrorampha rejectana |
| status |
|
Unterscheidung zwischen D. rejectana View in CoL und der D. plumbana -Artengruppe
Phänotypische Merkmale ( Abb. 8 View Abb ): D. rejectana und D. plumbana sind in beiden Geschlechtern nach äusseren morphologischen Merkmalen gut zu unterscheiden. Die GrÖssenunterschiede fallen sofort ins Auge und betreffen gleichermassen auch alle anderen Arten aus der D. plumbana -Gruppe. Die Männchen von D. rejectana haben eine Spannweite von 18–20 mm, die Weibchen von 16–17,5 mm. D. plumbana ist in beiden Geschlechtern deutlich kleiner (11–15,5 mm). Die GrÖssenverhältnisse zwischen den Geschlechtern sind bei den Arten gegenläufig: bei D. rejectana sind die Männchen erheblich grÖsser als die Weibchen, bei D. plumbana sind die Weibchen tendenziell grÖsser als die Männchen.
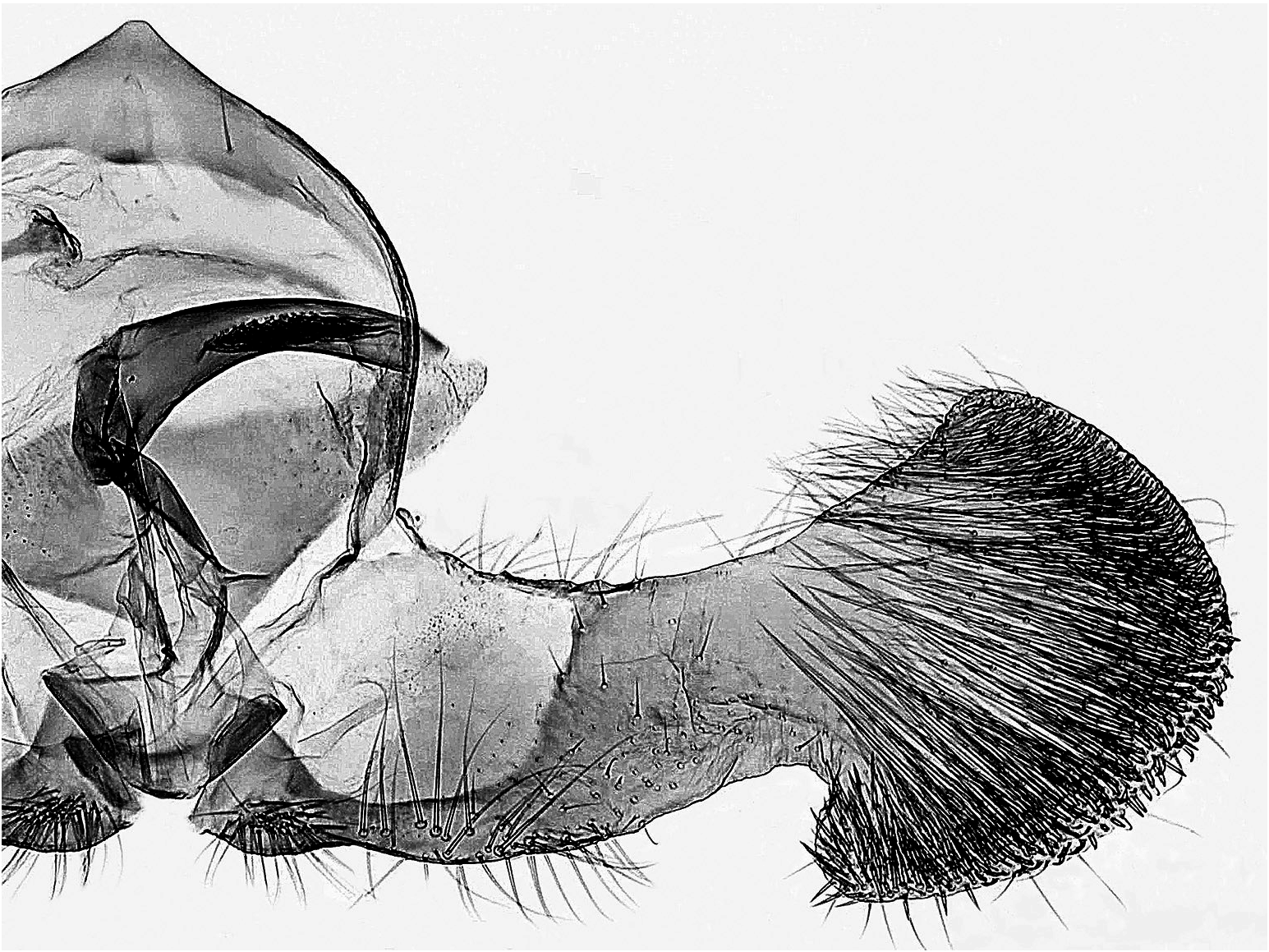
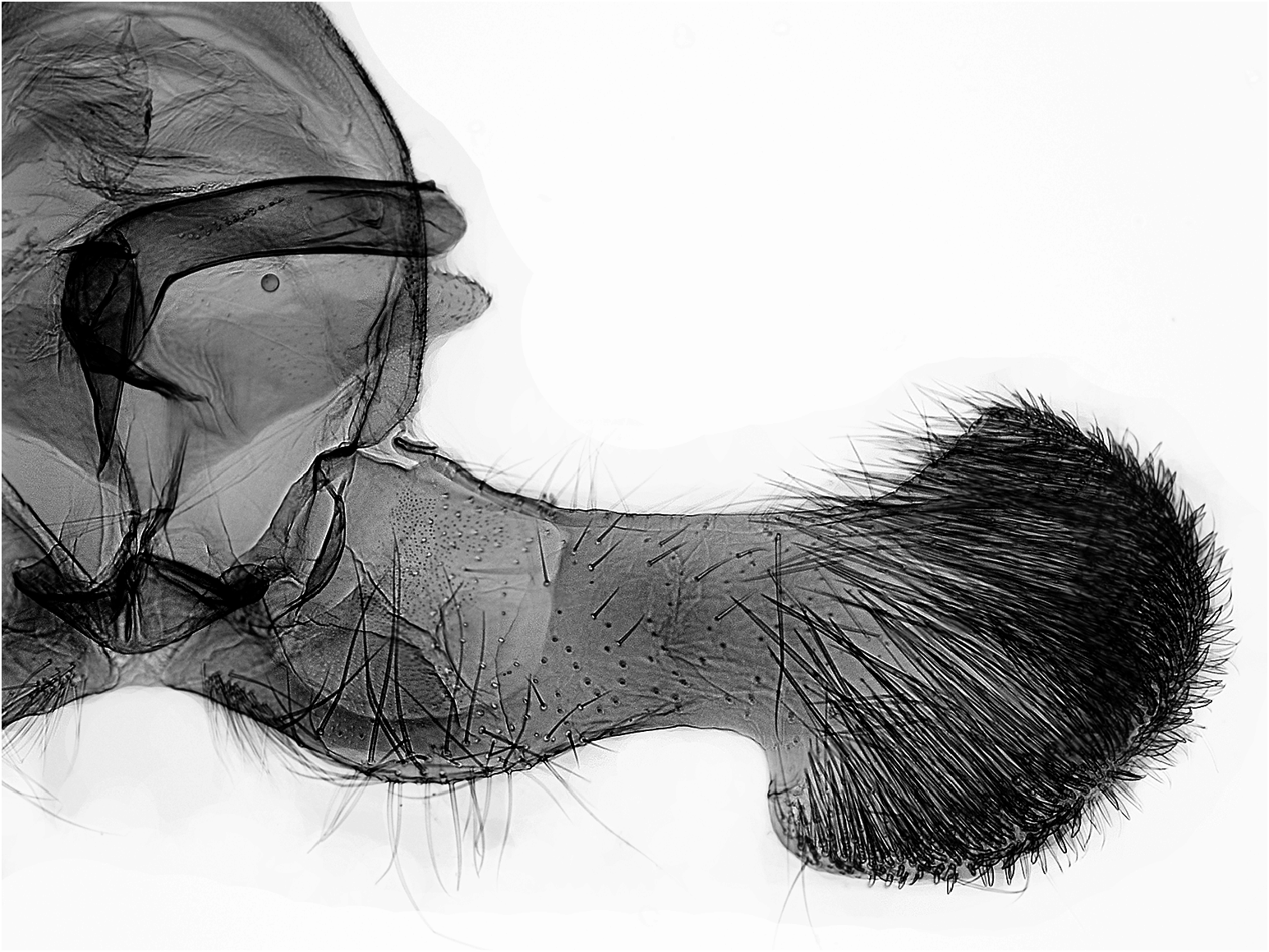
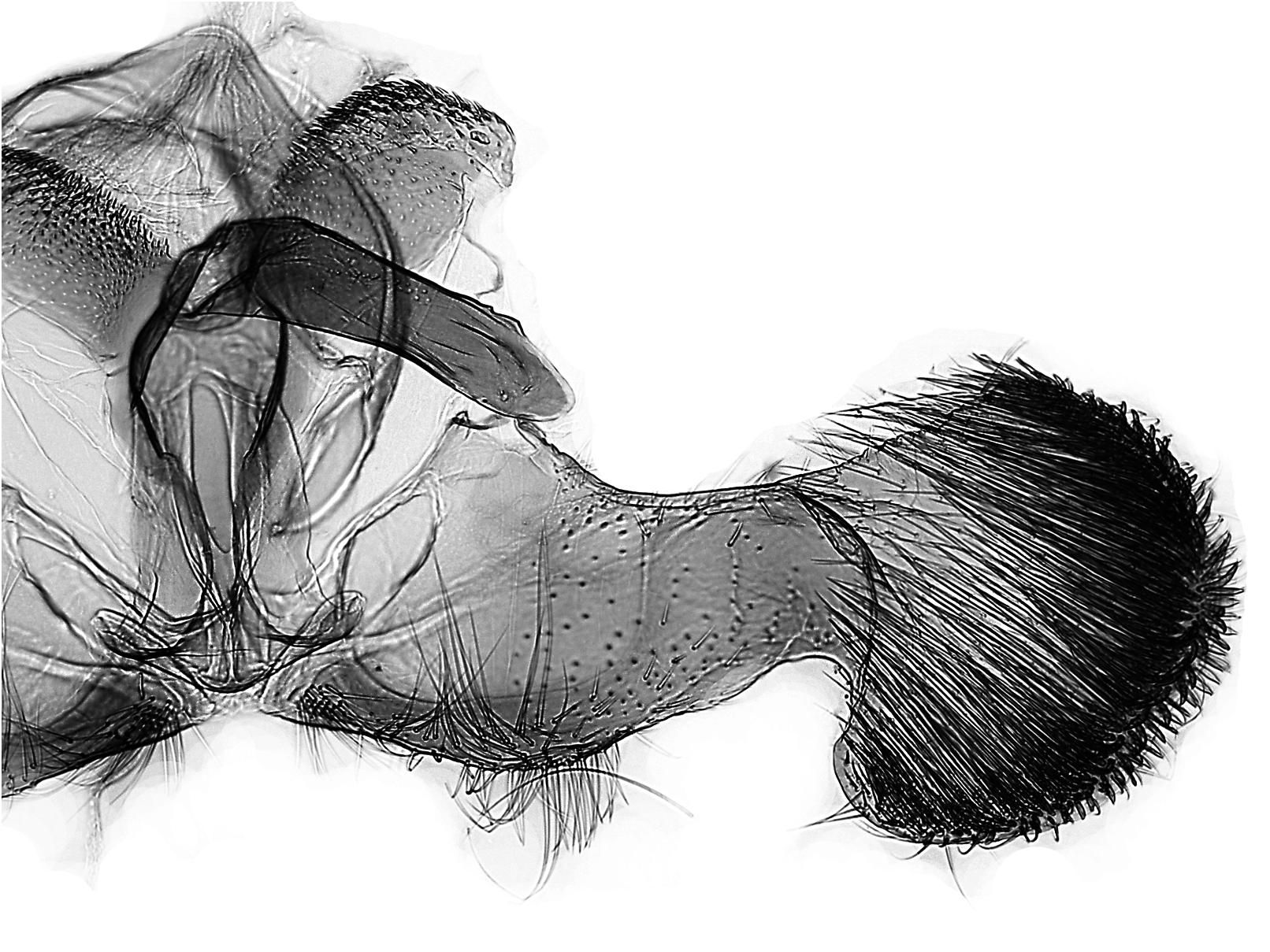
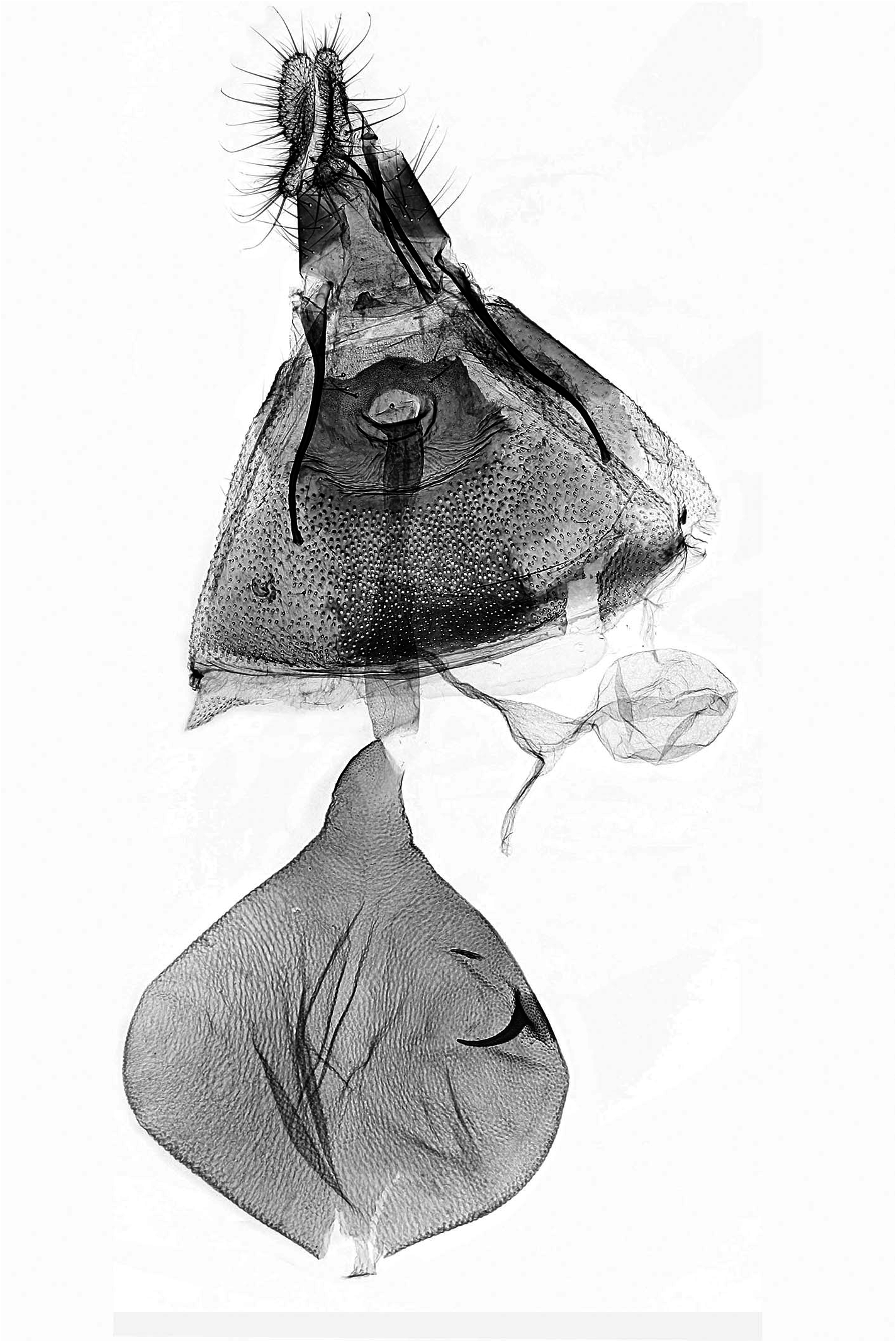
Unterschiede in der Genitalmorphologie
Männchen ( Abb. 9–12 View Abb View Abb View Abb View Abb ): D. rejectana hat im Gegensatz zu D. plumbana und den nahe verwandten Arten deutlich gestrecktere Valven, und in Relation zur Länge des Valvenarmes ist die Valvenbasis bei D. rejectana schmaler. Die Anellus-Lappen sind bei D. rejectana schlank und schwach bedornt, bei D. plumbana und D. aeratana breiter und kräftig bedornt. Der Aedoeagus von D. rejectana hat keinen Zahn vor der Spitze (vgl. D. sedatana und D. aeratana ).
Weibchen ( Abb. 13–16 View Abb View Abb View Abb View Abb ): Bei D. rejectana ist das Sterigma von feinen, gebogenen Leisten durchzogen, von denen einige annähernd parallel zum proximalen Rand des Sterigmas verlaufen. Bei der D. plumbana -Gruppe ist das Sterigma gleichmässiger sklerotisiert. Sofern Leisten erkennbar sind, verlaufen sie längs zur KÖrperachse. Das Colliculum ist bei D. rejectana gering sklerotisiert, relativ kurz und geht ohne Verjüngung in den Ductus bursae über. In der D. plumbana -Gruppe ist das Colliculum länger, stärker sklerotisiert und geht verjüngt in den Ductus bursae über.
Molekulare Daten
Insgesamt konnten vollständige Barcode-Sequenzen jeweils eines Männchens und Weibchens von D. rejectana ermittelt werden. Sie weisen keine intraspezifische Divergenz auf, unterscheiden sich jedoch signifikant von allen anderen bisher sequenzierten Arten, darunter insbesondere die bislang von einigen Autoren als konspezifisch bewertete D. plumbana und mit dieser nächstverwandte Taxa. D. rejectana weist im Barcode eine Distanz von 3,53% zum nächsten Nachbarn D. inconspiqua (DANILEVSKY, 1948) auf, eine Art, von der in BOLD 2 Sequenzen aus Bulgarien vorliegen. Auch der einzige ebenfalls aus Bulgarien stammende Barcode von D. infuscata (DANILEVSKY, 1960) ist mit einer Divergenz von 4,13% deutlich unterschieden. In Mitteleuropa bestehen die grÖssten genetischen Ähnlichkeiten zu D. alpigenana (HEINEMANN, 1863) , die sich allerdings mit minimaler Divergenz von 3,96% ebenfalls deutlich von D. rejectana absetzt. Schliesslich weisen auch die im Barcode untereinander nicht sicher unterscheidbaren Arten D. plumbana ( SCOPOLI, 1763) mit 5%, D. aeratana (PIERCE & METCALFE, 1915) mit 5,46%, D. sedatana BUSCK, 1906 mit 5,13% und D. tarmanni HUEMER, 2009 mit 5,29% signifikante Distanzen zu D. rejectana auf ( Abb. 17 View Abb ).
Biologie und Verbreitung
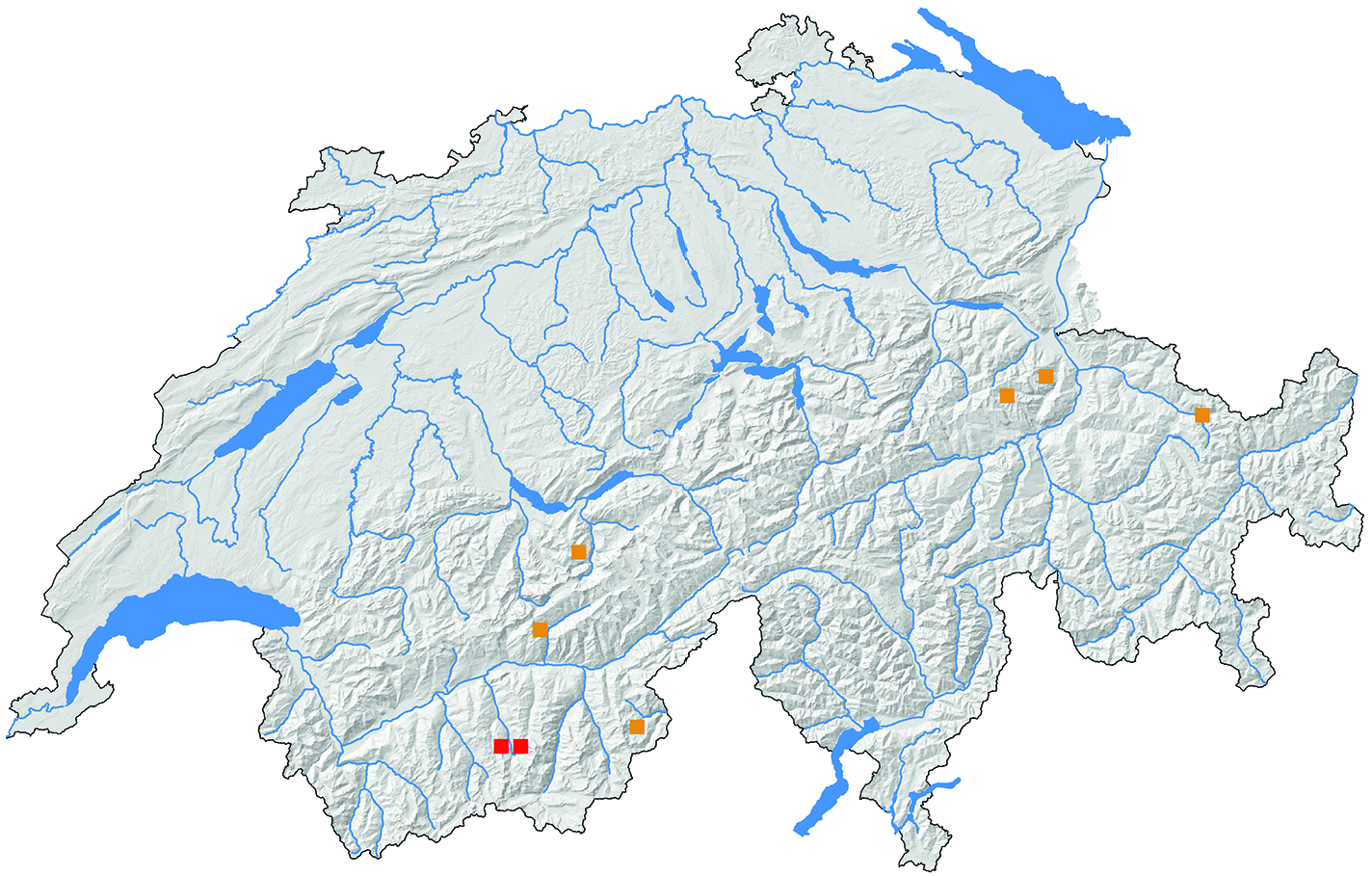
Die uns vorliegenden Funddaten von D. rejectana lassen auf eine Flugzeit der Falter von Ende Juni bis Anfang August schliessen (Grenzdaten: 28. Juni – 9. August). Die tägliche Hauptaktivitätszeit scheint am späten Nachmittag und Abend zu sein. Ob die Falter nachts auch ans Licht kommen, ist bislang unbekannt. Die Fundorte liegen auf HÖhen von 1500 bis 2100 Metern über Meer, also in der subalpinen Stufe der Alpen. Das Habitat im Zinaltal ist ein steiler, nach Norden und Osten gerichteter Berghang auf 1840–1860 m ü. M. Grünerlengebüsch ( Alnus alnobetula [= viridis]) und eine üppige Hochstaudenflur zeichnen den eher kühlfeuchten Lebensraum aus ( Abb. 18 View Abb ). In der Schweiz verteilen sich die Funde auf die Regionen Nordalpen, Wallis und Graubünden. Mit dem Nachweis aus dem Laggintal VS wird auch die Südschweiz gerade noch erreicht ( Abb. 19 View Abb ). Uns liegen keine Angaben über Funde ausserhalb der Schweiz vor.
Die ersten Stände von D. rejectana sind unbekannt. Aufgrund der Beobachtungen über die Entwicklungsstadien der nahe verwandten Arten kann man vermuten, dass sich die Raupen in den Wurzeln von Asteraceen entwickeln. Auffallend ist beispielsweise die Übereinstimmung der bisherigen Fundorte von D. rejectana mit der Verbreitung und den bevorzugten Wuchsorten von Achillea macrophylla und Achillea atrata ( Lauber & Wagner 2007) . Erstere der beiden ist in der Hochstaudenflur am Fundort im Zinaltal prominent vertreten.







| GP |
Instituto de Geociencias, Universidade de Sao Paulo |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.