
Echinoderes anniae, Sørensen & Rohal & Thistle, 2018
|
publication ID |
https://doi.org/ 10.5852/ejt.2018.456 |
|
publication LSID |
lsid:zoobank.org:pub:DE1B1DEE-9871-4803-9F67-025F2B439560 |
|
DOI |
https://doi.org/10.5281/zenodo.3818848 |
|
persistent identifier |
https://treatment.plazi.org/id/45B173E7-B6E9-443D-8C8F-3C424183DAFD |
|
taxon LSID |
lsid:zoobank.org:act:45B173E7-B6E9-443D-8C8F-3C424183DAFD |
|
treatment provided by |
Valdenar |
|
scientific name |
Echinoderes anniae |
| status |
sp. nov. |
Echinoderes anniae sp. nov.
urn:lsid:zoobank.org:act:45B173E7-B6E9-443D-8C8F-3C424183DAFD
Figs 2–3 View Fig View Fig , Tables 2–3 View Table 2 View Table 3
Diagnosis
Echinoderes with middorsal spines on segments 4, 6 and 8, and spines in lateroventral positions on segments 6 to 9. Tubes present in laterodorsal positions on segment 10, and glandular cell outlets type 2 in midlateral positions on segment 1, in subdorsal, laterodorsal, sublateral and ventrolateral positions on segment 2, in lateral accessory positions on segment 5, and in sublateral positions on segment 8. Tergal extensions of segment 11 extend into thin, flexible tips; sternal extensions short, rounded, with ventrolateral seta-like tuft of extended fringe tips. Males with three pairs of penile spines; females unknown.
Etymology
The first author (MVS) dedicates this species to his mother, Anni Larsen.
Material examined
Holotype
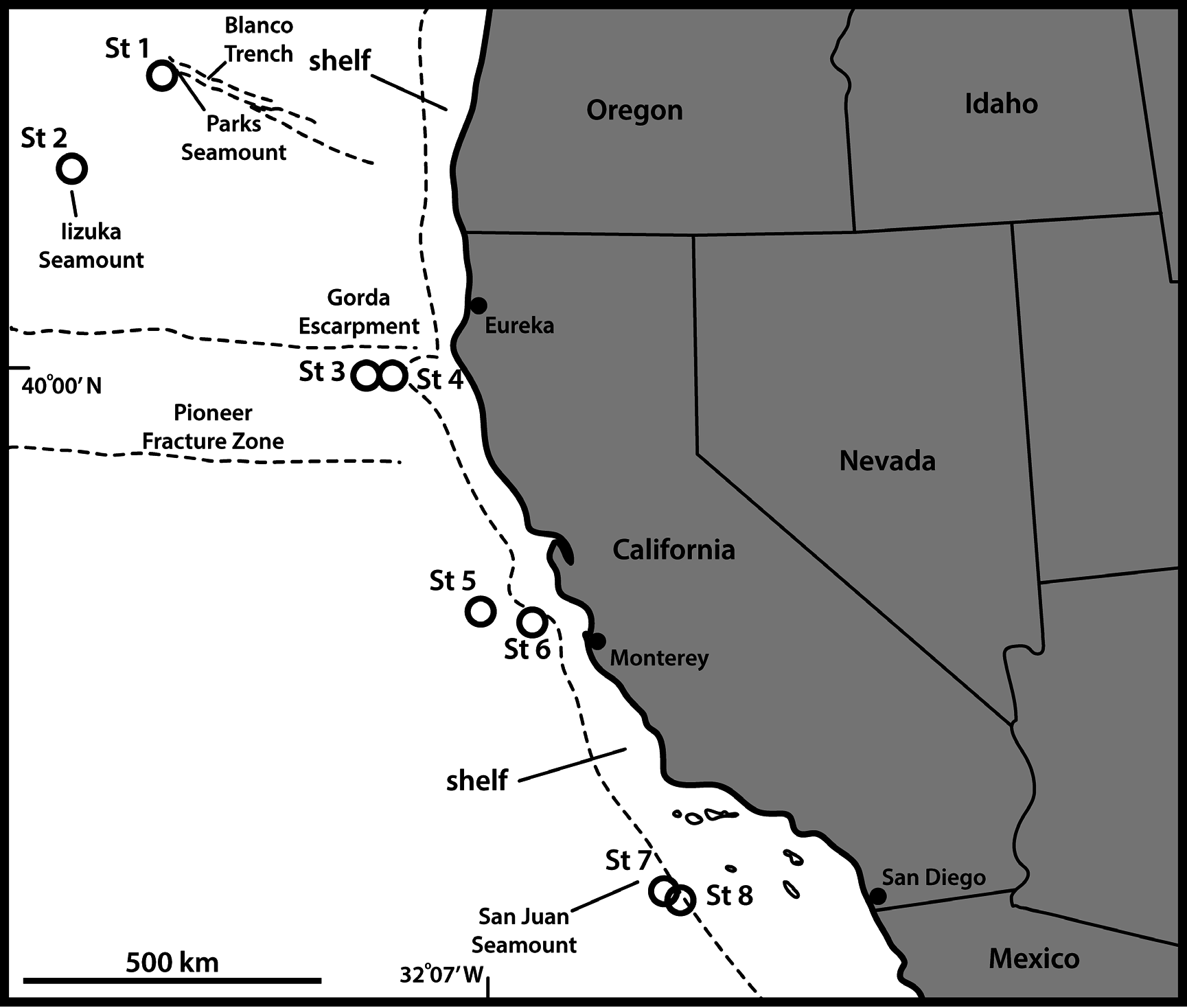
UNITED STATES OF AMERICA: adult ♂, off Oregon, US West Coast , 43°59′50″ N, 130°23′36″ W, St. 1, 3250 m deep,collected from mud, 16 Sep. 2008, mounted on an H-S slide in Fluoromount G (NHMD- 226483). No other specimens were found. See Fig. 1 View Fig for locality and Table 1 View Table 1 for detailed station data.
GoogleMapsDescription
Adults with head, neck and eleven trunk segments ( Figs 2 View Fig A–B, 3A). The trunk appears stout, with very well-developed pachycycli, almost parallel-sided, and round in cross-section. Only male morphology is known. For complete overview of measurements and dimensions, see Table 2 View Table 2 . Distribution of cuticular structures, i.e., sensory spots, glandular cell outlets, spines and tubes, is summarized in Table 3 View Table 3 .
The head was fully retracted in the holotypic specimen; hence, no information on head morphology is available. The neck has 16 placids, measuring 12 µm in length. The midventral placid is broadest, measuring 13 µm in width at its base, whereas all others are narrower, measuring 9 µm in width at their bases. Four dorsal and two ventral trichoscalid plates are present, with the ventral ones being broader than the four dorsal ones.
Segment 1 consists of a complete cuticular ring. Sensory spots are located on anterior segment half, but not immediately at the anterior margin, in subdorsal and laterodorsal positions ( Figs 2A View Fig , 3B View Fig ), and more posteriorly, in ventrolateral positions ( Figs 2B View Fig , 3C View Fig ). Glandular cell outlets type 2 are present in midlateral positions ( Figs 2 View Fig A–B, 3B–C), and glandular cell outlets type 1 in middorsal ( Fig. 3B View Fig ) and lateroventral positions. The segment appears to be devoid of cuticular hairs. The posterior segment margin is nearly straight, terminating in a well-developed pectinate fringe.
Segment 2 consists of a complete cuticular ring, with glandular cell outlets type 2 located in subdorsal, laterodorsal, sublateral and ventrolateral positions ( Figs 2 View Fig A–B, 3B–C). Pachycyclus of the anterior segment margin is well-developed, with weak indications of interruptions in middorsal and lateroventral positions. Sensory spots are located in laterodorsal and ventromedial positions. A single glandular cell outlet type 1 is located in middorsal position. Secondary pectinate fringe not detected on this or any of the following segments. They are evenly distributed around the segment. The posterior segment margin is nearly straight, and consists of a pectinate fringe with well-developed fringe tips.
Segment 3, and remaining segments, consist of one tergal and two sternal plates ( Figs 2B View Fig , 3A, C, E View Fig , G–H). Pachycyclus of the anterior segment margin is well-developed, and interrupted at the tergosternal junctions, but also middorsally. Sensory spots are located in subdorsal and midlateral positions ( Fig. 2 View Fig A– B). Glandular cell outlets type 1 are located in ventromedial positions ( Fig. 3B View Fig ). Cuticular hairs evenly distributed over tergal plate, between secondary fringe and intersegmental joint line, whereas the sternal plates only have hairs on their most lateral halves. Posterior segment margin straight, with pectinate fringe formed by well-developed fringe tips.
Segment 4 with acicular spine in middorsal position ( Figs 2A View Fig , 3B View Fig ). Sensory spots not observed. Anterior part of segment with glandular cell outlets type 1 in paradorsal and ventromedial positions. Pachycycli, pectinate fringe of posterior margin and cuticular hairs as on preceding segment.
Segment 5 without spines or tubes. Glandular cell outlets type 2 present in lateral accessory positions ( Figs 2B View Fig , 3E View Fig ). Sensory spots present in subdorsal, midlateral and ventromedial positions, and glandular cell outlets type 1 in middorsal and ventromedial positions. Pachycycli, pectinate fringe of posterior margin and cuticular hairs as on preceding segment.
Segment 6 with acicular spines in middorsal and lateroventral positions ( Figs 2 View Fig A–B, 3E). Sensory spots present in midlateral and ventromedial positions, and glandular cell outlets type 1 in paradorsal and ventromedial positions. Pachycycli, pectinate fringe of posterior margin and cuticular hairs as on preceding segment.
Segment 7 with acicular spines in lateroventral positions ( Figs 2B View Fig , 3E View Fig ). Sensory spots present in subdorsal, midlateral and ventromedial positions, and glandular cell outlets type 1 in middorsal and ventromedial positions. Pachycycli, pectinate fringe of posterior margin and cuticular hairs as on preceding segment.
Segment 8 with acicular spines in middorsal and lateroventral positions ( Fig 2 View Fig A–B, 3D, H). Middorsal spine is long, reaching the posterior margin of segment 10, and slightly thicker than the middorsal spine of segment 4. Glandular cell outlets type 2 present in sublateral positions ( Fig 2B View Fig , 3D, H View Fig ). Sensory spots and glandular cell outlets type 1 present in paradorsal and ventromedial positions. Pachycycli, pectinate fringe of posterior margin and cuticular hairs as on preceding segment.
Segment 9 with acicular spines in lateroventral positions ( Figs 2B View Fig , 3H View Fig ). Sensory spots present in paradorsal, laterodorsal ( Fig. 3D, F View Fig ) and ventrolateral positions, and glandular cell outlets type 1 in subdorsal and ventromedial positions. Small rounded sieve plates are present in sublateral positions ( Fig. 3F, H View Fig ). Pachycycli, pectinate fringe of posterior margin and cuticular hairs as on preceding segment. Segment 10 with laterodorsal tubes at posterior segment margin ( Figs 2A View Fig , 3F View Fig ); tubes in males are long and well-developed (ca 9 µm); female condition of tubes is unknown. Sensory spots present in subdorsal and ventrolateral positions. Glandular cell outlets type 1 present as two middorsal ones and one pair in ventromedial positions. Tergal plate only with a few cuticular hairs on the most posterior part; sternal plates with cuticular hairs on the lateral halves, but fewer than on preceding segment. The posterior segment margin of the tergal plate is straight, and only interrupted where the laterodorsal tubes attach, whereas the margins of the sternal ones are concave, extending posteriorly near the midventral junction. Pachycycli and pectinate fringe of posterior margin as on preceding segment.
Segment 11 with lateral terminal spines, almost equal to trunk in length ( Fig. 2C View Fig ). Males with three pairs of thin and flexible penile spines ( Figs 2A View Fig , 3G View Fig ); female condition unknown. Sensory spots present in subdorsal positions near the posterior margin of the segment. The segment is completely devoid of cuticular hairs. Tergal extensions narrow gradually into long, slightly flexible tips ( Fig. 3G View Fig ). Sternal extensions are short and rounded. Posterior margin of sternal plates with paired tufts of extended fringe tips ( Fig. 2B View Fig ); they resemble ventrolateral setae, but the structures appear to be formed by long, merged fringe tips from the posterior margin.
Remarks
The new species is very easily recognized by the nature of its cuticular structures in the lateral series of segment 5. The presence of a glandular cell outlet type 2 in lateral accessory positions is rare among congeners and shared with only E. unispinosus ( Yamasaki et al. 2018b) . A second trait, also exclusively shared with E. unispinosus , is the presence of midlateral glandular cell outlets type 2 on segment 1. However, E. unispinosus has only a single middorsal spine (see below in the present contribution and Yamasaki et al. 2018b); hence, the two species are easily distinguished.
The lack of lateroventral tubes on segment 5, combined with the presence of lateroventral spines on segments 6 to 9, is rather rare among species of Echinoderes , and only shared with E. arlis Higgins, 1966 , E. drogoni , E. koreanus Adrianov in Adrianov & Malakhov, 1999 , E. orestauri Pardos, Sánchez & Herranz, 2016 and E. svetlanae Adrianov in Adrianov & Malakhov, 1999 (see Higgins 1966; Adrianov & Malakhov 1999; Pardos et al. 2016a; Grzelak & Sørensen 2018, in press). However, E. anniae sp. nov. is easily distinguished from the latter four, since they all have five middorsal spines, opposed to three in E. anniae sp. nov. The species shows most resemblance with E. arlis . Both species have middorsal spines on segments 4, 6 and 8, and lateroventral spines on segments 6 to 9. Echinoderes arlis does not have lateral accessory glandular cell outlets type 2 on segment 5, but instead has sublateral tubes on this segment, but the attachment points of these tubes could be confused with gland outlets ( Grzelak & Sørensen 2018, in press). It furthermore shows the occasional presence of sublateral tubes on segment 8 (Grzelak & Sørensen in press), and the attachment points of these tubes could also be confused with gland outlets in E. anniae sp. nov., although the gland outlets in E. anniae sp. nov. are considerably larger than the tube attachments in E. arlis . Also the nearly parallel-sided trunks and the relatively welldeveloped pachycycli give the two species a fairly similar appearance. The main differences between the two species appear to be linked to the possession of either tubes or glandular cell outlets type 2. In addition to the differences on segments 5 and 8, the two species can be distinguished in a similar way on segment 2. Both species have glandular cell outlets type 2 in subdorsal and sublateral positions, but they differ in the laterodorsal positions where only E. anniae sp. nov. has glandular cell outlets type 2, and in the ventrolateral positions where E. arlis has tubes instead of glandular cell outlets type 2 ( Grzelak & Sørensen 2018, in press). Furthermore, E. anniae sp. nov. has glandular cell outlets type 2 in midlateral positions on segment 1, which are certainly not present in E. arlis . Hence, it is possible to distinguish the two species in different ways, but the several similarities suggest a close relationship between E. anniae sp. nov. and E. arlis , and potentially also E. unispinosus .


| US |
University of Stellenbosch |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |