
Echinolittorina meleagris (Potiez & Michaud, 1838)
|
publication ID |
https://doi.org/ 10.11646/zootaxa.2184.1.1 |
|
persistent identifier |
https://treatment.plazi.org/id/03D3606F-A505-FF81-FF26-FC99FDD7FBCD |
|
treatment provided by |
Felipe |
|
scientific name |
Echinolittorina meleagris |
| status |
|
Echinolittorina meleagris View in CoL (Beck in Potiez & Michaud, 1838)
( Figures 3 View FIGURE 3 , 4 View FIGURE 4 , 5A, B View FIGURE 5 , 6 View FIGURE 6 )
Phasianella meleagris Beck in Potiez & Michaud, 1838: 311 (no locality; types in Muséum de Douai, destroyed during WWII, P. Bouchet, pers. comm.; neotype, here designated, BMNH 20080962, Fig. 3L View FIGURE 3 , St Vincent).
Littorina (Melarhaphe) meleagris View in CoL — Mörch, 1876: 141 (as Melaraphe View in CoL ). Tryon, 1887: 248, pl. 45, fig. 10 (as Melaraphe View in CoL ). Rosewater, 1970: 424 (as? Melarhaphe View in CoL ). Abbott, 1974: 68, fig. 558. Andrews, 1977: 80, fig. H.E. Vokes & E.H. Vokes, 1983: 14, pl. 4, fig. 6 (as Melaraphe View in CoL ).
Littorina meleagris View in CoL — Aguayo, 1934: 88–89. Bequaert, 1943: 19–21, pl. 6, fig. 17–19. Abbott, 1954a: pl. 19j. Coomans, 1958: 61–62, pl. 8. Lewis, 1960: 416–417, fig. 11A, B (egg capsule), D (veliger) (as maleagris). Coomans, 1963: 125. Kaufman & Götting, 1970: 349, fig. 37. Borkowski, 1971: fig. 2 (egg capsule). Rosewater & Vermeij, 1972: 67–69, map 1, figs 3, 4 (in part, includes E. meleagris View in CoL B sensu Williams & Reid 2004). Flores, 1973a: 13, pl. 2, figs 16–19. Bandel, 1974a: 102, 107, fig. 14B and 17 (egg capsule), 18–22 and 52–53 (radula). Bandel, 1975: 17, pl. 1, figs 11, 12, pl. 2, fig. 4 (larval shell). Borkowski, 1975: 369–377, fig. 1D (radula). García-Talavera, 1983: 42 (in part, includes E. meleagris View in CoL B sensu Williams & Reid 2004). Bandel, 1984: 11, fig. 19 (radula). De Jong & Coomans, 1988: 19, pl. 32, fig. 82.
Littorina (Neritrema) meleagris View in CoL — Warmke & Abbott, 1961: 53, pl. 9n.
Littorina (Melarapha) meleagris View in CoL — Rehder, 1962: 122.
Littorina (Fossarilittorina) meleagris View in CoL — Rosewater, 1981: 29–30, fig. 4J (penis) (in part, includes E. meleagris View in CoL B sensu Williams & Reid 2004). Díaz & Puyana, 1994: 125, pl. 34, fig. 404 (as Fossalittorina).
Fossarilittorina meleagris View in CoL — Bandel & Kadolsky, 1982: table 1. Reid, 1986: 46, fig. 12 (egg capsule). Britton & Morton, 1989: 86, fig. 4-5B.
Nodilittorina (Fossarilittorina) meleagris View in CoL — Reid, 1989: 98, figs 2f, 7i (penis), 10g (pallial oviduct), 11i (egg capsule), 14i (radula). Taylor & Reid, 1990: 208 (shell microstructure).
Nodilittorina meleagris View in CoL — Redfern, 2001: 28, pl. 14, fig. 115. Reid, 2002a: 259–281, figs 1H, 2G (penis), 4 (pallial oviduct), 5B (radula).
Echinolittorina meleagris View in CoL — Williams, Reid & Littlewood, 2003: 60–86.
Echinolittorina meleagris View in CoL A— Williams & Reid, 2004: 2227–2251, fig. 6A (map).
Phasianella punctata L. Pfeiffer, 1840: 255–256 ( Cuba; types not in BMNH, presumed lost; not Turbo punctatus Gmelin, 1791 = E. punctata ).
Litorina guttata Philippi, 1847: 197 View in CoL , Litorina View in CoL pl. 4, fig. 7 (new name for Phasianella punctata L. Pfeiffer, 1840 ). Weinkauff, 1878: 34, pl. 4, figs 3, 4 (pl. 4 by Küster 1856). Weinkauff, 1883: 221.
Litorina (Litorina) guttata View in CoL — Dall, 1889: 146. Dall & Simpson, 1901: 430.
Littorina guttata View in CoL — Johnson, 1934: 102.
Littorina hidalgoi Arango, 1880: 159 View in CoL (La Habana and La Chorrera, Cuba; syntypes in Azpeita Moros Colln, Madrid, not seen; for date of publication see Azpeitia Moros, 1925). Azpeitia Moros, 1925: 165–168, pl. 1, fig. 5.
Taxonomic history: The original description ( Potiez & Michaud 1838) was not accompanied by an illustration, but is sufficiently precise to define unambiguously the species hitherto known as E. meleagris View in CoL ; in particular, the aligned white spots, paler colour of the upper part of the whorls, smooth surface and presence of an umbilicus are together diagnostic. However, it is now believed that the forms of E. meleagris View in CoL in the western and eastern Atlantic are not conspecific ( Williams & Reid 2004), despite their closely similar shells, and the epithet must be attached to one or the other. Therefore a type specimen is required for Phasianella meleagris , and a neotype is here designated.
The identity of Phasianella punctata L. Pfeiffer, 1840 is slightly doubtful because the description is brief and does not mention an umbilicus. Recognizing the secondary homonymy, Philippi (1847) renamed this taxon Litorina guttata ; although he too did not mention the umbilicus, it is visible in his figure.
Being small and of cryptic habit, this species was poorly known in the nineteenth century. Although described three times, in 1838, 1840 and again in 1880, it was brought to wider attention by Philippi (1847) as Litorina guttata . It continued to be known by this name ( Weinkauff 1878, 1883; Dall 1889; Dall & Simpson 1901), until the earlier name L. meleagris became widely accepted ( Mörch 1876; Tryon 1887; Bequaert 1943). It was not included in the western Atlantic monograph of Nodilittorina by Bandel & Kadolsky (1982), because it was considered to belong to the genus Fossarilittorina . On the basis of divergent DNA sequences (12S rRNA gene) Williams & Reid (2004) distinguished ‘ E. meleagris A’ and ‘ E. meleagris B’, from the western and eastern Atlantic respectively, as ‘evolutionarily significant units’ (ESUs).
Diagnosis: Shell small; smooth except for microstriae and one incised peripheral line; pseudumbilicus; brown with quincuncial array of white spots. Penis simple, tapering, no glands, closed sperm duct. Caribbean Sea, Bahamas, E Florida, Bermuda. COI: GenBank AJ622953 View Materials , AJ622954 View Materials .
Material examined: 120 lots (including 11 penes, 5 sperm samples, 11 pallial oviducts, 1 spawn sample, 4 radulae).
Shell ( Fig. 3 View FIGURE 3 ): Mature shell height 3.0– 9.3 mm. Shape high turbinate to elongate succineiform (H/B = 1.43–1.76, SH = 1.50–2.03); spire whorls rounded; suture distinct; spire profile straight; shoulder sometimes thickened; periphery rounded. Columella long, straight or slightly convex at midpoint, narrow, slightly flattened at base; deep pseudumbilicus as wide as columella; no eroded parietal area. Sculpture absent but for fine spiral microstriae over entire surface and single incised line just above periphery (often worn smooth). Protoconch 0.28 mm diameter, 2.4–2.5 whorls. Ground colour brown to black, paler on base, with white spots (or backward-pointing triangles) regularly arranged in prosocline and opisthocline series (i.e. in quincuncial array); spots usually become larger and may anastomose at suture to give paler sutural zone (but not a distinct band); spots may align to create partial (or even complete) oblique opisthocline stripes ( Fig. 3C, D View FIGURE 3 ); rarely entire shell may be whitish with pale grey marbling ( Fig. 3N View FIGURE 3 ); aperture brown with external pattern showing through, pale zone at base (rarely forming a distinct band); columella brown.
Animal ( Fig. 4 View FIGURE 4 ): Head ( Fig. 4H View FIGURE 4 ) black to grey, with unpigmented stripe across snout; tentacle pale around eye and across base, with two short (occasionally long) longitudinal black stripes and a black dot at tip; sides of foot grey to black. Opercular ratio 0.36–0.42 (exceptionally 0.31). Penis ( Fig. 4A–G View FIGURE 4 ): wrinkled base merges into simple narrow tapering filament; vas deferens closed as a superficial duct (running from anterior end of open prostate, across right side of head, to tip of penial filament); penis unbranched and lacking mamilliform glands or penial glandular disc; penis unpigmented except at very base. Euspermatozoa 57–71 µm; paraspermatozoa ( Fig. 4N–P View FIGURE 4 ) containing 1–2 elongate-fusiform rod-pieces 24–30 µm (exceptionally to 43 µm) with pointed to rounded ends, usually projecting from cell, which is packed with round granules. Pallial oviduct ( Fig. 4I, J View FIGURE 4 ): copulatory bursa separates near anterior end of straight section and extends back to three-quarters of length of straight section; posterior third of straight section contains an opaque cream or reddish gland around the egg groove (this does not appear to be an extension of the small translucent capsule gland). Spawn ( Fig. 4K–M View FIGURE 4 ): an asymmetrically biconvex pelagic capsule 240–269 µm diameter by 110 µm high (180 by 100 µm, Borkowski 1971; 200 µm, Bandel 1974a) with broad peripheral rim slightly overhanging base, dome-shaped upper side sculptured by 0–3 concentric rings, containing single ovum 70–80 µm diameter ( Lewis 1960: Barbados; Borkowski 1971: Florida; Bandel 1974a: Colombia; pers. obs.: Florida).
Radula ( Fig. 5A, B View FIGURE 5 ): Relative radula length 1.21–1.90. Rachidian: almost square, length/width 0.83–1.10; major cusp broad, tip pointed. Lateral and inner marginal: 4 cusps, tip of major cusp rounded to slightly truncate. Outer marginal: 4–6 cusps; no flange on outer side of base.
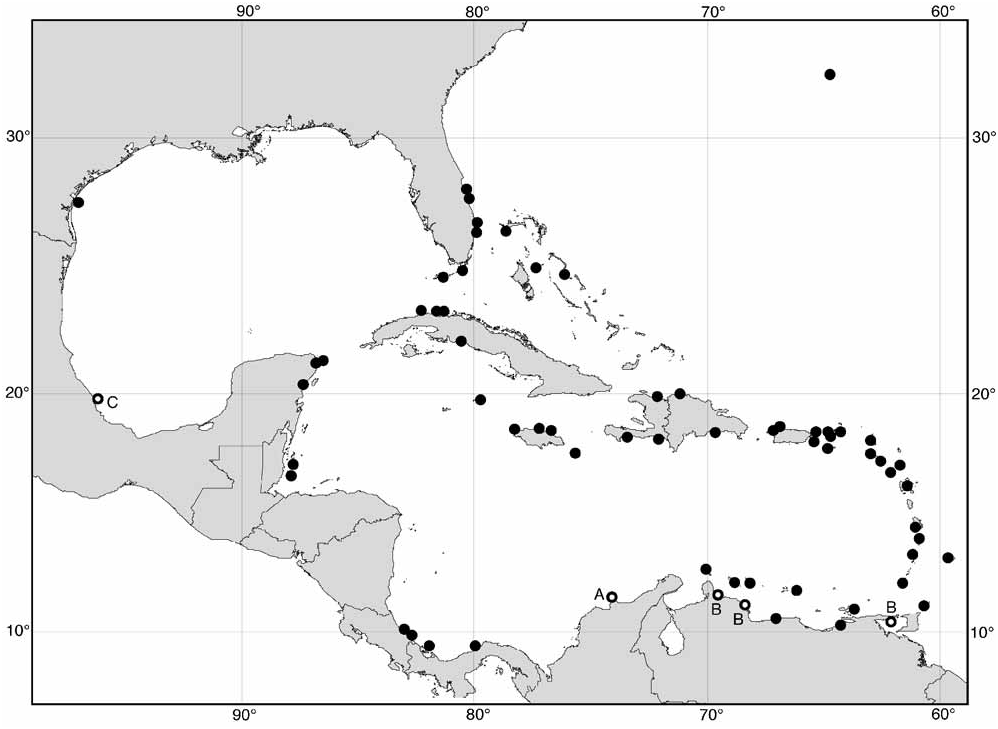
Range ( Fig. 6 View FIGURE 6 ): Throughout Caribbean Sea, northern Bahamas, E Florida, Bermuda, rare in Gulf of Mexico. Range limits: Washington Oaks State Park , Flagler Co, Florida (H.G. Lee, pers. comm.); Sebastian Inlet, Brevard Co., Florida ( BMNH 20080958 ) ; Boca Chica Key, Florida ( USNM 820431 View Materials ) ; Rocky Bay , Bermuda ( USNM 773845 View Materials ) ; Eight-mile Rock, Grand Bahama I., Bahamas ( BMNH 20080960 ) ; Jacks Bluff , Cotton Bay, Eleuthera I., Bahamas ( AMS) ; Les Trois Pavillons, Dept. NW, Haiti ( USNM 439443 View Materials ) ; Virgin Gorda, British Virgin Is ( BMNH 20080964 ) ; Nenton Point , Antigua ( BMNH) ; Silver Sands , Christ Church, Barbados ( BMNH) ; Arnos Vale Bay , Tobago ( BMNH 20080956 ) ; Cumana, Venezuela ( USNM 709134 View Materials ) ; Caracas , Venezuela ( MNHN) ; Bonaire, Netherlands Antilles ( ZMA) ; Santa Marta, Colombia ( Bandel 1974a); Fort Sherman , Colon, Panama ( USNM 620507 View Materials ) ; Portete, Limón Prov., Costa Rica ( USNM 706437 View Materials ) ; South Water Cay , Belize ( BMNH 20080957 ) ; Isla Mujeres, Quintana Roo, Mexico ( USNM 662938 View Materials ) ; Punta del Morro , Veracruz, Mexico ( Britton & Morton 1989); Lucea Bay, Jamaica ( BMNH 20080959 ) ; Cienfuegos, Cuba ( BMNH 20080963 ) ; La Habana, Cuba ( BMNH) ; Port Aransas , Texas ( USNM 664275 View Materials ; Andrews 1977) .
This species is common in eastern Florida, western Cuba and throughout most of the Caribbean Sea where there is suitable hard substrate and conditions are not too turbid, estuarine or eutrophic. There are only three records from the Gulf of Mexico, two from Port Aransas, Texas, and one from Veracruz, presumably because conditions there are too continental. However, E. meleagris is also uncommon in the Bahamas (only four records, all from localities in the north, compared with 18 for the similarly small-shelled E. mespillum ) and there are no records from eastern Cuba (cf. 3 for E. mespillum ). Except for the archipelago of Las Aves, Flores (1968, 1973a) reported records in Venezuela only from continental localities, although other offshore localities are reported here. This suggests that very oceanic conditions are also unsuitable for E. meleagris . Abbott & Jensen (1967) were the first to report this species from Bermuda.
This has been considered to show an amphiatlantic distribution ( Rosewater & Vermeij 1972; Rosewater 1981; García-Talavera 1983), but the West African form is not conspecific ( Williams & Reid 2004; see Remarks below).
Habitat: Recorded from coral limestone and sandstone; mid-eulittoral, in lower barnacle zone and in algal turf; sheltering in pits on exposed shores, also in stagnant algal pools in upper eulittoral and littoral fringe on sheltered coasts. Absent from continental shores with turbid water and from strongly oceanic shores.
Many ecologists have observed that E. meleagris occupies the lowest shore zone of all littorinids in the tropical western Atlantic ( Flores 1973b; Vermeij 1973a; Yipp & Carefoot 1988; Britton & Morton 1989). At Santa Marta, Colombia, it is most common in the eulittoral in shallow warm pools, but is also found up to several centimetres below low water level among dense algae; it is preyed upon by Thais and Leucozonia ( Bandel 1974a; Brattström 1980). It occupies the mixed algal zone of the upper eulittoral in Panama (Brattström 1986) and both the eulittoral yellow zone and pools in the black zone higher on the shore in the Bahamas ( Newell et al. 1959; G.L. Voss & N.A. Voss 1960; Brattström 1992, 1999). In wave-exposed situations it shelters among algal rocks in the low eulittoral ( Bingham 1973; Redfern 2001). On beachrock in Barbados it grazes indiscriminately, but was found to move upward when other littorinids were removed, suggesting competitive effects ( Yipp & Carefoot 1988). McLean (1967) measured the erosion of beachrock by the grazing activity of this species.
Corresponding with its low-level and moist habitats, this species is the most sensitive of co-occurring littorinids to desiccation, suffering 100% mortality after 3 days in air. It has a significantly larger area of ciliated gill filaments (as opposed to unciliated folds extending across mantle roof) than other Echinolittorina species ( Yipp & Carefoot 1988). It is also intolerant of high temperatures; 50% survived 1 hour at 42–43°C ( Fraenkel 1968). However, it is reported to be moderately euryhaline, tolerating salinity in the range 26–36 ppt, as may be encountered in high-level pools ( Flores 1973b).
Lewis (1960) reported eggs in November and December at Barbados; capsules were released singly or in strings, but the latter were probably cases of abnormal premature release. Capsules were released in June in Florida (pers. obs.).
Remarks: Members of the Littorininae typically occupy the littoral fringe and may extend down into the barnacle zone of the upper eulittoral. The mid or even low eulittoral habitat of E. meleagris is unusual in the subfamily. Correspondingly, it is less tolerant of desiccation and high temperature than its congeners.
This species has been recorded in the eastern Atlantic, from Ghana to Angola ( Rosewater & Vermeij 1972; Rosewater 1981; García-Talavera 1983). West African specimens do not differ anatomically, but the white subsutural band on the shell is usually more distinct (pers. obs.). On the basis of sequence data from the mitochondrial 12S gene Williams & Reid (2004) suggested that the eastern and western Atlantic forms were distinct ESUs, although their phylogenetic relationships were not resolved. Sequence data for the COI gene have since been added, showing them as sister ESUs in a three-gene analysis ( Williams & Duda 2008; Fig. 1 View FIGURE 1 ), and gives an average K2P genetic distance for COI of 4.53% ( Fig. 37 View FIGURE 37 ). The widely disjunct distribution, limited genetic data and small difference in shell colour are provisionally considered sufficient to recognize the two as distinct species. By a parsimony argument (outgroup comparison with sister taxon E. mespillum ), dispersal across the Atlantic took place from west to east ( Vermeij & Rosenberg 1993), and may have occurred around the same time as that of E. ziczac , in the upper Pliocene or early Pleistocene ( Williams & Reid 2004; Williams & Duda 2008).




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Echinolittorina meleagris
| Reid, David G. 2009 |
Echinolittorina meleagris
| Williams, S. T. & Reid, D. G. 2004: 2227 |
Echinolittorina meleagris
| Williams, S. T. & Reid, D. G. & Littlewood, D. T. J. 2003: 60 |
Nodilittorina meleagris
| Reid, D. G. 2002: 259 |
| Redfern, C. 2001: 28 |
Nodilittorina (Fossarilittorina) meleagris
| Taylor, J. D. & Reid, D. G. 1990: 208 |
| Reid, D. G. 1989: 98 |
Fossarilittorina meleagris
| Britton, J. C. & Morton, B. 1989: 86 |
| Reid, D. G. 1986: 46 |
Littorina (Fossarilittorina) meleagris
| Diaz, J. M. & Puyana, M. 1994: 125 |
| Rosewater, J. 1981: 29 |
Littorina (Melarapha) meleagris
| Rehder, H. A. 1962: 122 |
Littorina (Neritrema) meleagris
| Warmke, G. L. & Abbott, R. T. 1961: 53 |
Littorina meleagris
| De Jong, K. M. & Coomans, H. E. 1988: 19 |
| Bandel, K. 1984: 11 |
| Garcia-Talavera, F. 1983: 42 |
| Bandel, K. 1975: 17 |
| Borkowski, T. V. 1975: 369 |
| Bandel, K. 1974: 102 |
| Flores, C. 1973: 13 |
| Rosewater, J. & Vermeij, G. J. 1972: 67 |
| Coomans, H. E. 1963: 125 |
| Lewis, J. B. 1960: 416 |
| Coomans, H. E. 1958: 61 |
| Bequaert, J C 1943: 19 |
| Aguayo, C. G. 1934: 88 |
Littorina guttata
| Johnson, C. W. 1934: 102 |
Litorina (Litorina) guttata
| Dall, W. H. & Simpson, C. T. 1901: 430 |
| Dall, W. H. 1889: 146 |
Litorina guttata
| Weinkauff, H. C. 1883: 221 |
| Weinkauff, H. C. 1878: 34 |
Littorina (Melarhaphe) meleagris
| Vokes, H. E. & Vokes, E. H. 1983: 14 |
| Andrews, J. 1977: 80 |
| Abbott, R. T. 1974: 68 |
| Rosewater, J. 1970: 424 |
| Tryon, G. W. 1887: 248 |
| Morch, O. A. L. 1876: 141 |
Phasianella punctata L. Pfeiffer, 1840: 255–256
| Pfeiffer, L. 1840: 256 |