
RHINATREMATIDAE, Nussbaum, 1977
|
publication ID |
https://doi.org/ 10.1111/j.1096-3642.2012.00838.x |
|
persistent identifier |
https://treatment.plazi.org/id/03DB87B7-FFE2-FFAE-FEFA-9668FE776180 |
|
treatment provided by |
Marcus |
|
scientific name |
RHINATREMATIDAE |
| status |
|
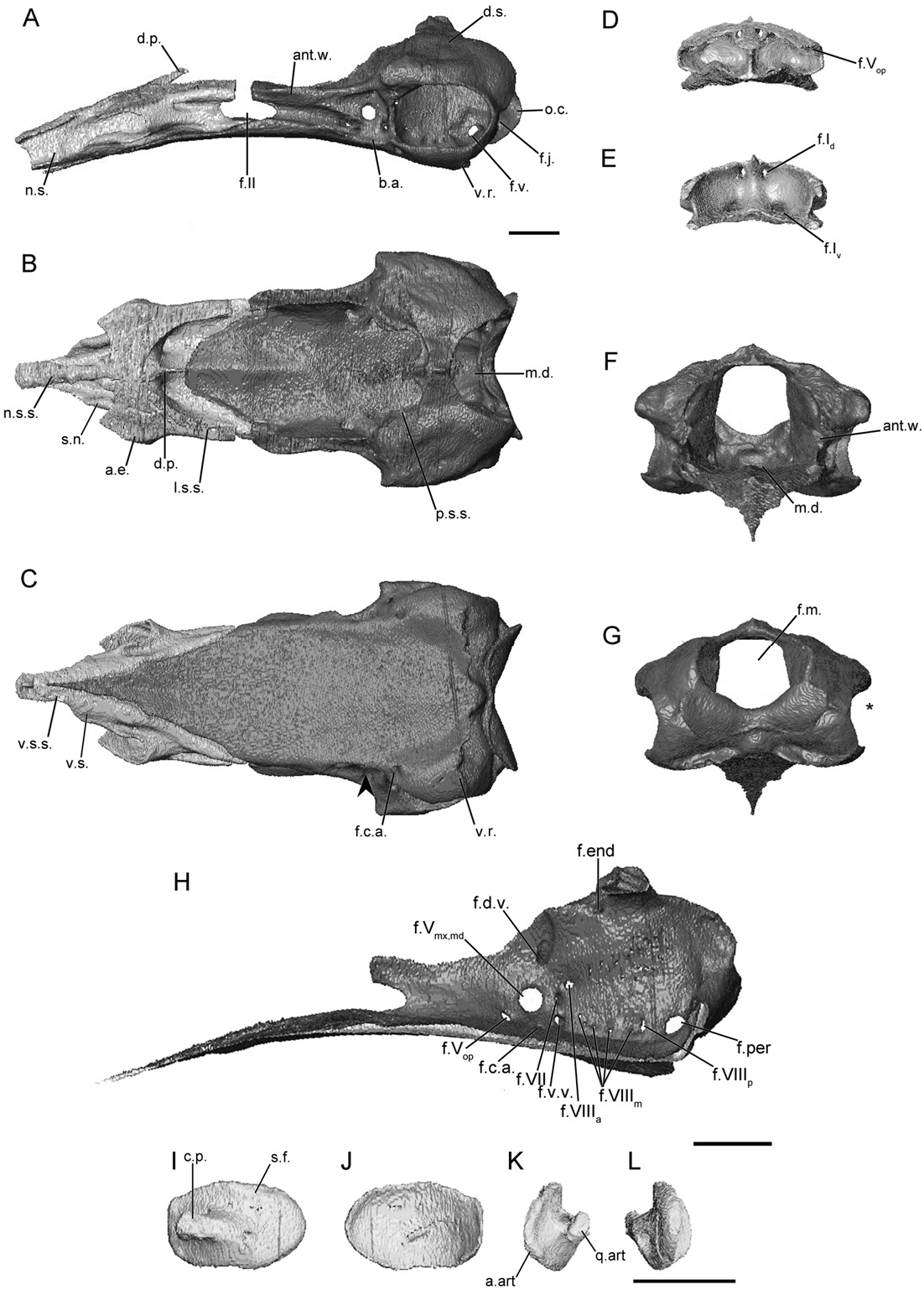
RHINATREMATIDAE View in CoL ( FIGS 7 View Figure 7 , S15)
The main body is relatively long, accounting for just under half the total length of the sphenethmoid ( Fig. 7A View Figure 7 ). The lateral wall of the main body is long, and is capped by a broad sutural surface that receives the parietal ( Fig. 7A View Figure 7 ). A broad ossified anterolateral process is present, and in E. bicolor it is connected medially to the anterior dorsal sutural surface ( Fig. 7B View Figure 7 ). The dorsomedial process is thin and rod-like ( Fig. 7B View Figure 7 ), and is completely covered dorsally. The posterior margin of the lateral walls is deeply incised (less so in R. bivittatum ) by the anterior margin of the optic foramen ( Fig. 7A View Figure 7 ). The posterior margin of the floor is also deeply concave and U-shaped in outline ( Fig. 7B View Figure 7 ).
The nasal septum is moderately long, terminating posterior to the external naris. It remains uniform in height distally ( Fig. 7A View Figure 7 ). The dorsal sutural surface is narrow near its base and expands towards the tip ( Fig. 7A View Figure 7 ). The ventral surface also maintains a broad sutural surface as far as the tip of the nasal septum. A broad sheet of bone forms a shelf ventral to the exit points of the ventral branch of the olfactory nerve, connecting the sola nasi to the nasal septum ( Fig. 7B,C View Figure 7 ).
The foramina in the anterior wall are paired, and the foramina of the ventral pair are located slightly more laterally from the midline than the dorsal pair. A single anterolateral foramen is present in the base of each expanded anterolateral process ( Fig. 7E View Figure 7 ).
The wall is parallel to subparallel to the contralateral wall when viewed dorsally ( Fig. 7B View Figure 7 ). In anterior view the antotic wall is vertically orientated ( Fig. 7F View Figure 7 ). Posteriorly the dorsal sutural surface is expanded slightly, and is continuous with a portion of the surface on the anterior margin of the otic capsule. The anterior margin of the antotic wall is deeply incised by the posterior margin of the oval optic foramen ( Fig. 7A View Figure 7 ), less so in R. bivittatum (Fig. S15A). Five foramina pierce the antotic wall ( Fig. 7H View Figure 7 ). These have the same configuration, and transmit the same structures as those seen in the dermophiid species (referred to here as Pattern 1; Maddin, 2011). An additional foramen just ventral to that for the dorsal vein is present in E. bicolor . Histological data reveal that this additional foramen transmits a vein. This foramen is absent on the right side of the head in BM 1946.9.5.66 ( Fig. 7H View Figure 7 ).
The dorsal surface of the otic-occipital complex is nearly horizontal, and forms the entire occipital surface (the parietal does not contribute; Fig. 7A View Figure 7 ). It tapers dramatically in width towards the midline ( Fig. 7A View Figure 7 ). A sutural surface that receives the parietal is present along the anterior margin of the surface, but narrows substantially at the midline and is absent at the midline in R. bivittatum (Fig. S15A). A process extending laterally, just anterior and dorsal to the fenestra vestibuli, is present but poorly developed in E. bicolor ( Fig. 7B View Figure 7 ). The region occupied by the inner ear is large and fills almost the entire extent of the otic-occipital complex in lateral view. The fenestra vestibuli is large and slightly ovoid ( Fig. 7A View Figure 7 ). In posterior view the lateral edges of the otic-occipital complex are incised by the fenestra vestibuli ( Fig. 7G View Figure 7 ). The occipital condyle is small, and in lateral view it is almost level with the outline of the posterior margin of the otic-occipital complex ( Fig. 7A View Figure 7 ). The jugular foramen in the base of the condyle is only partially visible in lateral view. The foramen magnum is large and subcircular in outline ( Fig. 7G View Figure 7 ).
The medial wall of the otic capsule is perforated by eight foramina ( Fig. 7H View Figure 7 ). The four foramina common to all species are present in their usual locations (endo- and perilymphatic foramina, anterior and posterior vestibulocochlear nerve foramina). Four small foramina conducting components of the medial branch of the vestibulocochlear nerve occur along the ventral margin ( Maddin, 2011).
The anterior portion of the floor of the os basale is triangular in outline. It is relatively very long, extending to the region of the anterior half of the nasal septum. A thin medial ridge that fits into a groove in the ventral surface of the sphenethmoid is present. The floor of the os basale is nearly flat and the median posterior depression is only weakly defined ( Fig. 7B View Figure 7 ). A well-developed basicranial articulation is absent from rhinatrematids ( Fig. 7A, C View Figure 7 ), and only a narrow shelf lateral to the antotic wall is all that represents the articulation. In ventral view there is no constriction of the lateral margin of the floor posterior to the basicranial articulation (arrowhead; Fig. 7C View Figure 7 ). A very weak ridge is present instead of a wing-like ventral projection on the ventral surface of the otic capsule ( Fig. 7A View Figure 7 ). Posteriorly the ventral surface of the floor of the os basale terminates in a rounded point that closely approaches the ventral margin of the foramen magnum ( Fig. 7C View Figure 7 ).
The foramen that leads to the canal carrying the carotid artery enters the ventral surface of the floor of the os basale, ventral to the anterior limit of the otic capsule ( Fig. 7C View Figure 7 ). The canal terminates at lateral and medial foramina located just ventral to the trigeminal foramina ( Fig. 7H View Figure 7 ).
The footplate of the stapes is large and ovoid in rhinatrematids ( Fig. 7I, J View Figure 7 ), and its anterior margin is closely applied to the anterior margin of the fenestra vestibuli. The remaining margins nearly fill the fenestra. The medial surface of the footplate is strongly concave. The columellar process is short, rod-like, and extends anteriorly in roughly the horizontal plane. The base of the columellar process is pierced by a foramen ( Fig. 7L View Figure 7 ). This foramen is variably present in one specimen ( FMNH 152310 View Materials ), being present on the right and absent from the left .
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |