
Eurycleonus talamellii, Meregalli, Massimo, 2005
|
publication ID |
https://doi.org/ 10.5281/zenodo.170025 |
|
DOI |
https://doi.org/10.5281/zenodo.6266164 |
|
persistent identifier |
https://treatment.plazi.org/id/C451E476-EB15-6E0A-2138-FE34FD52FA14 |
|
treatment provided by |
Plazi |
|
scientific name |
Eurycleonus talamellii |
| status |
sp. nov. |
Eurycleonus talamellii View in CoL new species
Figs 1–7 View FIGURE 1 View FIGURES 2 – 8
Type locality: Morocco, Tan Tan province, 2 km NW Tan Tan, 28°27’N 11°07’W
Type material. Holotype %. MOROCCO: “SW Morocco, 28°27’N 11°07’W, 2 km NW Tan Tan, 80 m, 16–18.xii.2003, J. Kaláb legit” (coll. Meregalli).
Paratypes: same data, 5 % 22 Ψ (5 % 6 Ψ coll. Meregalli; 16 Ψ coll. Talamelli); “SW Morocco, 28°27’N 11°07’W, 2 km NW Tan Tan, 80 m, 28.xi / 2.xii.2003, J. Kaláb legit”, 1 % 8 Ψ (3 Ψ coll. Meregalli; 1 % 5 Ψ coll. Talamelli); “SW Morocco, 28°27’N 11°07’W, 2 km NW Tan Tan, 80 m, 29.xii.2004, Meregalli legit”, elytra of 5 specimens (coll. Meregalli); “S Morocco, 30 km W Es Smara, 28. IV.2004, Gianasso legit” [26°39’N 11°57’W] 3 Ψ (found dead and partly incomplete) (coll. Meregalli); “Tandalt, Bas Dra, Thami, 3.63 / Eurycleonus gigas, L. Kocher det.” [= Tadalt, 28°45’N 09°46’W] 1 % (Museo de Ciencias Naturales, Madrid).
Specimens not examined: MOROCCO: “TizguielHarratine” [29°13’N 08°32’W] (cited in Kocher 1961).
Diagnosis. A species of Eurycleonus characterized by the rostrum with middle line straight in lateral view, with upper margin of the scrobe sinuate and nearly reaching the lower part of the eye, the elytra with whitish and brownish scales in irregular patches, the aedeagus with apex short, not acutely pointed.
Measurements. Holotype: Body length including rostrum 24.10 mm. Rostrum: length 5.40 mm, width 2.21 mm. Pronotum: length 5.36 mm, width 6.00 mm (ratio 0.89). Elytra: length 13.70 mm, width 8.15 mm (ratio 1.68). Ratio of elytral to pronotal width 1.36.
Description. Body very robust, broadly elliptical, integument black, with dense bifid scales, antennae and tarsi dark reddish ( Fig. 1 View FIGURE 1 ).
Rostrum stout, sides slightly convergent from base to antennal insertion, apical part of upper margin of scrobe, above the antennal insertion, moderately and regularly constricted; median line distinct from base to antennal insertion, convex, moderately raised, not keeled, slightly broadened anteriorly, interrupted at antennal insertion, extended towards apex where it forms a broad central triangular plate extended beyond antennal insertion and joined apically to epistome; median line in lateral view nearly rectilinear, scarcely concave near base; dorsum with two paramedian shallow depressions, which are narrowed at antennal insertion and reach apex on either side of the broad triangular central plate; dorsolateral margins usually rounded, seldom slightly more prominent; sides glossy, with a deep narrow furrow above upper margin of scrobe and elongate rough punctures in front of eyes; upper margin of scrobe slightly keeled, sinuate, directed basad under lower margin of eye ( Fig. 7 View FIGURES 2 – 8 ). Vestiture: bifid lanceolate glossy white scales, cleft up to near base, densely covering dorsum except on median line, anterior dorsal plate and epistome; scales on sides sparse above scrobes, only dense inside the furrow above scrobes, relatively dense on pregenae and sides below scrobes; scales on underside sparse, missing from the glossy, nearly completely glabrous triangular prementum; a few hyaline setae appressed to integument are present on sides and underside, mainly near apex. Scape of antennae dark reddish, lighter at base, weakly curved, slender and narrow, only shortly thickened at apex, with some bi or trifid scales and a few setae near apex, mainly on the front side; funicle of relatively uniform thickness; segment 1 short, subglobose, barely longer than broad; 2nd conical, twice longer than wide and twice longer than the 1st; 3rd 5th subconical, isodiametric, 5th slightly smaller; 6th longer than wide; 7th conical, wider than the previous segments, appressed to, and apparently part of, club; club elongateoboval, segments 1 and 2 together slightly shorter than the 3rd, annulus of segment 3 distinct ( Fig. 2 View FIGURES 2 – 8 ); funicle segments 1 to 6 with few short scales not covering integument; 7th and club with elliptical whitish bifid scales, cleft up to their mid length, covering integument.
Head globose, eyes acutely oval, slightly convex, surrounded by dense white scales, shorter than those on rostrum.
Pronotum with base very weakly arched at middle; sides subparallel up to midlength, regularly and gently curving anteriorly, subsinuate apicad, apical margin widely rounded over head; surface smooth, finely micropunctured and with few scattered barely larger punctures, hardly distinct, delimited by a semicircular, slightly raised glossy margin on their outer side; central line present, extremely narrow, not raised, glossy; curvature of dorsum in lateral view regular, with maximum height at middle. Vestiture: white glossy bifid scales, lanceolate on sides and shorter towards dorsum, densely covering integument on dorsolateral and lateral parts; small bare spots are present in correspondence of the punctures; these bearing one narrow hyaline reclined seta as long as the scales; dorsum lacking scales, excepting towards the apical part.
Elytra elliptical, base sinuate, as wide as pronotal base, gently broadened beyond humeri, subparallel up to apical quarter; maximum width slightly beyond mid length of elytra, with a scarcely distinct compression at basal third, very regularly curved at apex; dorsum moderately and very regularly convex; intervals nearly flat, with a low convexity in the central part of their width; striae very narrow, with small, seriate, shallowly impressed punctures. Vestiture composed of white, light brown and brown bifid glossy lanceolate scales cleft up to near base; scales on suture light brown, darker towards the external margin; even intervals with usually white or very light brown scales; odd intervals with whitish or light brown scales at centre and irregular darker spots toward the striae; brown scales usually present along the striae and on the marginal part of the intervals, often alternate with patches of whitish scales; a white dense round patch is present on base of intervals 3 and 4; another white round patch is present on declivity, at junction of intervals 456; intervals 2, 4 and 6 with vague round whitish spots, alternate to less dense light brown scales.
Legs. Femora regularly and moderately thickened at middle, with dense bifid scales covering integument, excepting some small scattered bare maculae, bearing a reclined seta; fore tibiae rectilinear, apex perpendicularly truncate, with a strong apical inner tooth and a small expansion directed outwards; middle tibiae straight; hind tibiae curved outwards; fore tibiae with dense bifid white scales and moderately erect hyaline setae, mainly present on the inner side; middle and hind tibiae with denser coating of lanceolate, mainly undivided glossy white scales and numerous setae moderately projecting above integument. Tarsi slender, scaly; segments 1 and 2 triangular, isodiametric; 3 bilobed, longer than wide; onychium very long, slender; claws connate at middle, weakly divaricated.
Underside with dense coating of white to yellowish trifid scales, cleft up to near base and with very thin teeth; ventrites with dense coating of mainly bifid scales, white except for a narrow brown stripe at base of ventrites 3–5, with very small, inconspicuous bare dots bearing one hyaline seta.
Aedeagus: see figs 3–4.
Variation. Specimens examined of Eurycleonus talamellii are quite uniform for the majority of their morphologic traits. The females are larger and have broader elytra, with maximum width distinctly beyond mid length of elytra, as is typical of this genus; the size varies between 23.6 and 26.3 mm in males and between 26.8 and 30.7 mm in females, with a length/width ratio of elytra comprised between 1.60 and 1.75. In some specimens the rostrum is slightly more depressed longitudinally and with slightly raised dorsolateral margins; although the distribution of white and brownish scales on the elytra is variable, the appearance is always irregularly maculate; the two more distinct white patches at the base of intervals 4–5 and at the apex of intervals 4–6 are always present. Female genitalia: see figs. 5–6.
Remarks. Eurycleonus talamellii is a geographic and apparently taxonomic vicariant of E. gigas . The latter differs by several traits: body larger, elytra with more rounded sides, broadened towards apical third also in male, with a length/width ratio comprised between 1.4 and 1.5; rostrum with upper margin of scrobe curved inwards at antennal insertion; upper margin of scrobe, in dorsal view, often slightly expanded before antennal insertion; in lateral view dorsum more curved, particularly apicad, median line at base distinctly concave and upper margin of scrobe not sinuate, regularly curved and directed below lower margin of the eye ( Figs 7–8 View FIGURES 2 – 8 ); segments 1 and 2 of funicle shorter, segment 1 distinctly globose (cf. Meregalli 2000: 155, Fig. 13); sides of pronotum nearly subparallel, or very slightly convergent, from base to apical third, maximum width at base; scales on elytra nearly uniformly ivorywhite, or greywhite, very light brown only on odd intervals but not forming a distinct, although irregular, pattern of alternate light and dark dots; lacking the white round patches on base of intervals 3–4 and on apex of intervals 4–6; aedeagus more regularly curved in lateral view and with narrower, slightly more acute apex (cf. Meregalli 2000: 155, Fig. 10 View FIGURE 10 ). Eurycleonus amon differs by the smaller pronotum, with base sinuate, arched towards the elytra (cf. Meregalli 2000: 155, Fig. 4 View FIGURES 2 – 8 ), and, in lateral view, with maximum height near base. The new species lives together with Ammocleonus hieroglyphicus (Olivier 1807) and with Conorhynchus relictus Meregalli 2001 . Both species are smaller; the first differs by being winged, by the base of pronotum triangularly protruding towards elytra, by the longer rostrum, with high and sharp keel, and by the simple scales. Conorhynchus relictus is immediately recognized by its very short, conical rostrum.
Etymology. This species is named after my friend Fabio Talamelli, with whom I share the pleasure of studying one of the most fascinating subfamilies of Curculionidae , the Lixinae .
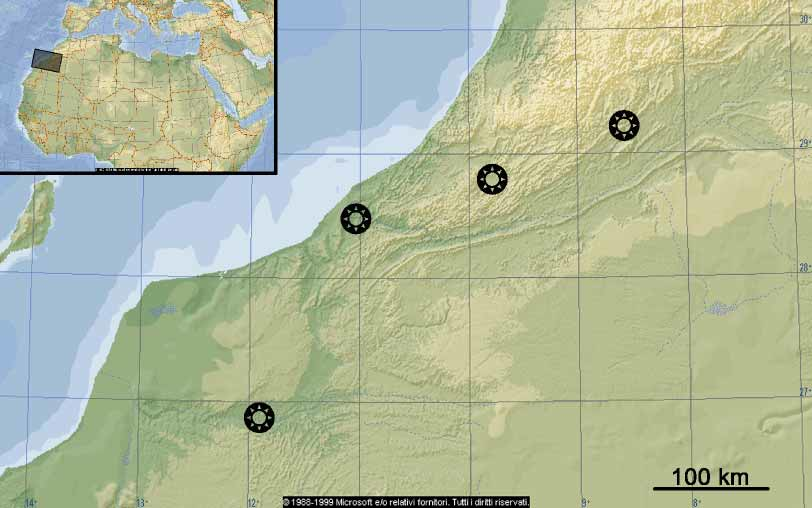
Distribution. The range so far known of E. talamellii includes the lower part of the Drâa valley, and nearby regions, and extends to the south until at least the surroundings of Es Smara, that is, between 26°39’N and 29°13’N. It is apparently endemic to Morocco ( Fig. 9 View FIGURE 9 ), although a marginal presence in northwestern Mauritania, in habitats similar to that of Es Smara, is probable.
Ecology. The species of Eurycleonus are strictly associated with sandy desert habitats ( Meregalli 2000), and E. talamellii is no exception. The type locality is a semidesert, gently sloping sandy hill, very near to the town of Tan Tan. Vegetation is comparatively dense and rich, covering 30%–40% of the surface, and is mainly composed of Zygophyllum gaetulum Emb & Maire (Zygophyllaceae) , Euphorbia echinus Hook. f. & Coiss. and E. regisjubae Webb & Berth. (both Euphorbiaceae ), with sporadic presence of several other plants, among which are Centaurea incana Desf. (Asteraceae) and Ver b en a supina L. ( Verbenaceae ) ( Fig. 10 View FIGURE 10 ). The climate is typical of a steppedesert, with low rainfall (102 mm /year), exclusively distributed in late fall and winter ( Fig. 11 View FIGURE 11 ) (data from New et al. 2002, reported in DIVAGIS 2004).
No living specimens were seen in this area in late December 2004 and early January 2005. However, several fragments, mainly elytra, were found buried in the sand below the larger shrubs of Z. gaetulum . Some woody rootstocks of these shrubs showed signs of apparent larval feeding, but no larvae were found in the roots or at the base of the larger stalks. No fragments of Eurycleonus were found below other plants. Data on the biology of the species of Eurycleonus are scarce, and always refer to Chenopodiaceae . Specimens of Eurycleonus amon (as E. gigas ) were found on Haloxylon schweinfurthii Ashers. and Anabasis articulata Moq. ( Kneucker 1909) and on Hammada elegans (Bunge) Botsch ( Alfieri 1976, sub Haloxylon salicornicum Boiss. ) whilst E. gigas was collected in the Grand Erg, Gassi Touil, on Cornulaca monacantha Del. ( Peyerimhoff 1931) . However, the host plants of the preimmaginal stages are not known. Chenopodiaceae have been recorded as the host plant of various species of Cleonini ( Hoffmann 1950; TerMinasyan 1988; Meregalli 2000), usually at the immaginal stage. Zygophyllaceae have been reported by Kneucker (1909) as being the host plant, in Sinai, of the preimmaginal stages of another Cleonine weevil, Porocleonus candidus (Olivier 1807) , a species whose adults are often found below Chenopodiaceae (Meregalli, personal observations in Tunisia). Zygophyllaceae dominate at Tan Tan, where only few and small plants of goosefoot are present. The occurrence of several fragments of Eurycleonus in the sand below shrubs of Zygophyllum gaetulum and the detection of signs of larval feeding in the woody rootstocks may suggest that, at the type locality, this is the host plant of E. talamellii , at least at the larval stages. Furthermore, the association of adults of Eurycleonus with Chenopodiaceae may indicate that the genus is linked to both Zygophyllaceae and Chenopodiaceae , in analogy to the apparent biology of Porocleonus candidus . However, the possibility that E. talamellii usually develops in Chenopodiaceae and that at the type locality Z. gaetulum should be regarded as a refuge plant (defined as in Colonnelli & Osella 1998) cannot be ruled out. More field researches are required to detail biology of Eurycleonus . In particular, it should be determined whether preimmaginal and immaginal stages are monophagous or if they can develop indifferently on Zygophyllaceae and Chenopodiaceae , and, also, if each species has a specialized biology and feeds upon different plant families.
The site 30 km west of Es Smara is a broad wadi, with vegetation mainly composed of scattered trees of Acacia raddiana Savi (Fabaceae) , shrubs of Pergularia tomentosa L. ( Asclepiadaceae ), several plants of Citrullus colocynthis (L.) Schrad. ( Cucurbitaceae ) and again Chenopodiaceae and Zygophyllaceae , covering not more than 5% of the surface. This sparsely vegetated stripe interrupts a sandy desert area nearly completely devoid of vegetation. Soil between the shrubs and trees is bare whitish sand ( Fig. 12 View FIGURE 12 ). The three specimens collected in this area were found dead in the sand (Gianasso, personal communication). No living specimen or fragments were found in January 2005. The climate of Es Smara is typical of a desert, with an extremely scarce annual rainfall (26 mm /year), occurring in fall and winter (data from New et al. 2002, reported in DIVAGIS 2004).
All living specimens collected at the type locality were found walking on the sandy soil, after a period of uncommon heavy rainfalls (Kaláb, personal communication). As previously reported, no living adults, notwithstanding very accurate investigations, were found at the type locality in December 2004 and January 2005, when no rain had previously fallen. However, it is unlikely that the life cycle of E. talamellii depends entirely on rainfalls, which are too scarce and unpredictable in these areas. All the other specimens of Eurycleonus examined, including those of E. gigas and E. amon , were found between March and early May; the specimens collected in late April and early May have usually vestiture of scales very damaged or missing (Meregalli, personal observations and Peyerimhoff 1931). These data suggest that these weevils may complete their larval cycle and metamorphose by the beginning of winter; an occasional rainfall may then trigger emergence, particularly in the areas which receive comparatively more rain; however, adult activity may be normally determined by other biotic or abiotic parameters, such as temperature, daylength, phenology of the host plants; it mainly occurs in late winter and early spring, when temperature is not too hot and the plants start vegetation and flowering. The life span of the adults is probably quite short, and by the time that day temperature gets too high they die, and only fragments can be found in the sand or below the host plants.
Discussion. The genus Eurycleonus has an apparently disjunct distribution, with one species in Sinai and two species in the centralwestern Sahara, between southern Tunisia and southern Morocco. Notwithstanding Peyerimhoff’s (1931) indication, for E. gigas , of “Tout le Sahara, du Sinai à l’Erg Iguidi, Cyrénaïque”, species of this genus seem to be absent from the central and eastern Sahara, in the Libyan and Egyptian deserts. Alfieri (1976) cited Eurycleonus only for Sinai, not for other territories of Egypt, and no specimens from Libya are reported in literature (Erg Iguidi is in central Algeria, not in Cyrenaica) or were found in several Museums personally visited. This apparent gap in distribution may, however, be due to scarcity of collections in the favourable habitats, and to the ephemeral appearance of the adult specimens, which renders their finding very difficult: excepting the exceptional record by Kneucker of about 300 exs of E. amon collected in two days in March 1902 in Sinai ( Kneucker 1909: 122, as E. gigas ), only a few tens of specimens belonging to this genus have been found since the description of E. gigas in 1868. The two species living in the western Sahara differ by relatively minor characters, and can be regarded as sister species, whereas the Sinaitic E. amon is more distinct; this suggests that two evolutionary lineages have developed, one in the eastern part of the deserts of north Africa and now probably restricted to Sinai, the second in the centralwestern Sahara. This second line may have undergone a speciation event by vicariance, giving rise to E. gigas and E. talamellii , when the habitats favourable to the persistence of populations of these weevils contracted and fragmented. Dryland vegetation communities developed and expanded in the middlelatitudes of the old world from mid Miocene to early Pliocene, and in early Pliocene the Sahara desert became fully established ( Tallis 1991). Diversification and / or dispersal of taxa linked to arid habitats in northern Africa may date back to this age. During the Pliocene dry and moist, sometimes very humid, phases alternated. In the drier periods a very high percentage of pollen of steppe and desert plants was recorded ( Dupont & Leroy 1994), whereas in the more humid phases the deserts contracted, and were at least in part replaced by a dense vegetation, even including rainforests ( Adams 2004). This alternation of climatic phases may have contributed to creating a fragmentation of the habitats suited for the survival of populations of Eurycleonus , with a subsequent speciation event by vicariance between the extremes of the range. These wingless weevils, highly adapted to peculiar environmental conditions, have obviously a limited dispersal capability, so that gene flow is reduced, particularly if the favourable habitats become fragmented and isolated, as it may have occurred during the Plio and Pleistocenic climatic oscillations. A significant comparison is offered by the winged Ammocleonus hieroglyphicus , a related weevil sympatric and syntopic with Eurycleonus , equally associated with steppe and deserts and usually found below Chenopodiaceae . Ammocleonus hieroglyphicus has a high dispersal capability by flight, so that, as opposed to the species of Eurycleonus , it is distributed throughout all of northern Africa and the Middle East region, without any particular morphological differences across its broad range.





No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.