
Eutyphlus spiralis Owens and Carlton, 2016
|
publication ID |
https://doi.org/ 10.1649/072.070.0102 |
|
persistent identifier |
https://treatment.plazi.org/id/038FA12D-FFB2-580D-FF51-382DFE23FD55 |
|
treatment provided by |
Diego |
|
scientific name |
Eutyphlus spiralis Owens and Carlton |
| status |
sp. nov. |
5. Eutyphlus spiralis Owens and Carlton View in CoL , new species
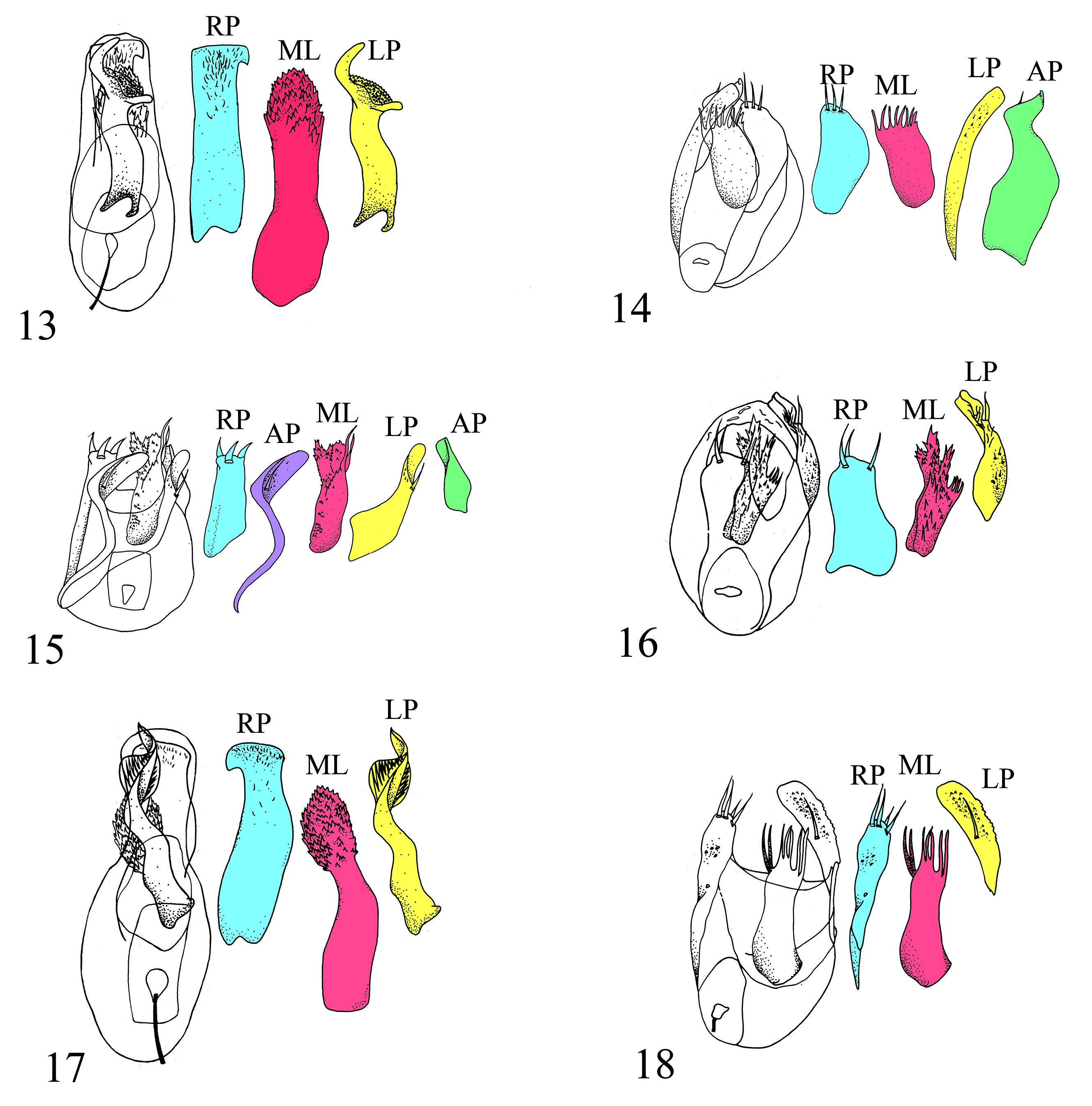
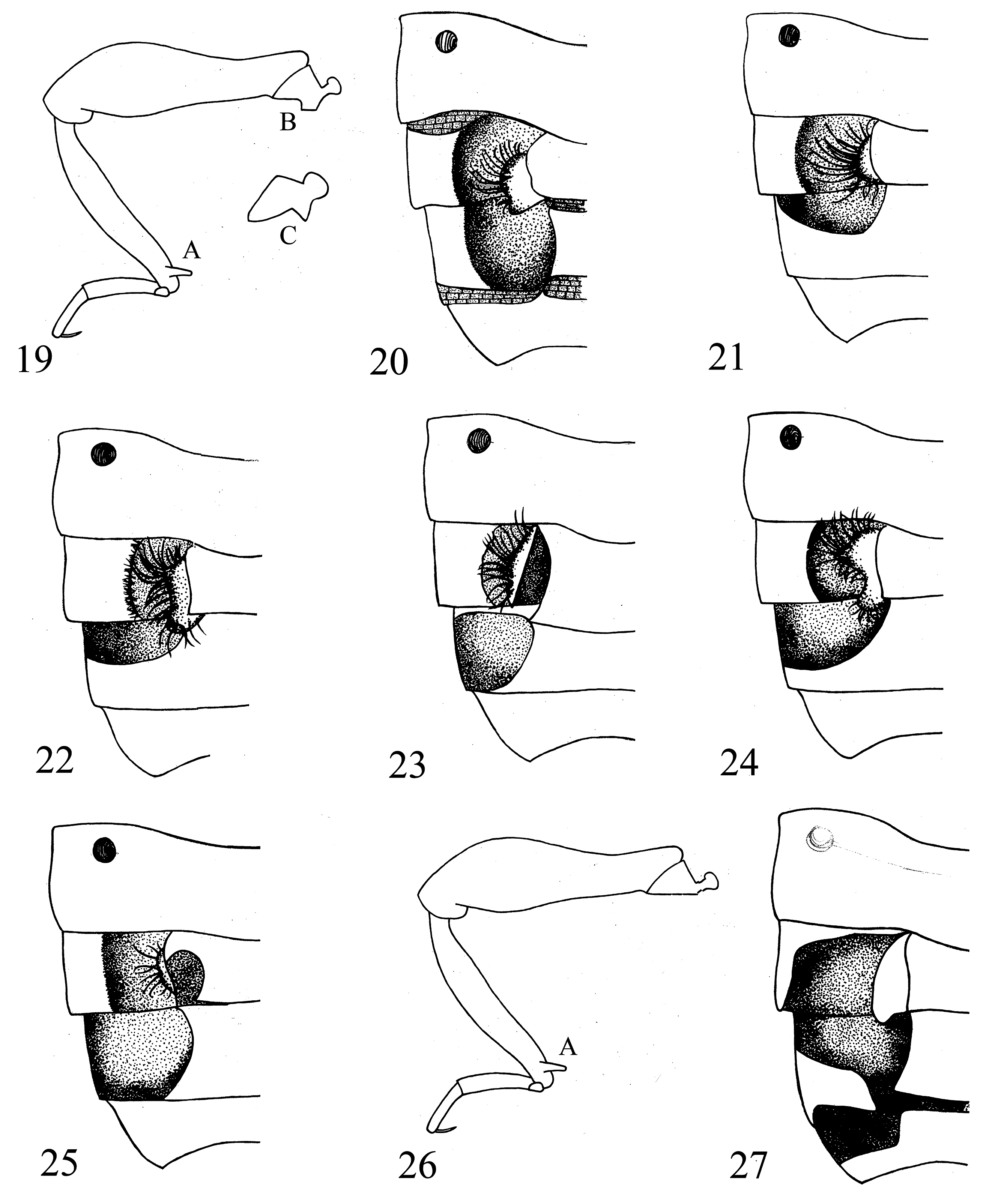
( Figs. 17 View Figs , 19A, 19C, 26 View Figs , 32 View Figs )
Holotype. Male. USA: NC. Macon Co. 10 mi SW Franklin; Back Country info center; V-22- 1991; Berlese oak & Rhododendron ; S.O’ Keefe ( FMNH) (dissected and slide-mounted in Euparal).
Diagnosis. Eutyphlus spiralis is similar to E. dybasi based on the absence of anteroprosternal foveae, presence of secondary sexual characters on the legs of males ( Figs. 19A, C, 26 View Figs ), quadrifoveate elytra, concave metaventrite, and larger, broadly ovate seventh ventrite of males. In E. spiralis , the right paramere of the aedeagus is ventrally located and expanded distally to the left (the right paramere of E. dybasi is expanded distally to the right), and the left paramere is dorsally located, twisted counterclockwise, and bears a series of spines along both sides of the process ( Fig. 17 View Figs ) (the left paramere of E. dybasi is expanded distally into a process that bears ctenidial spines along one face).
Description. MALE. Measurements: HL 0.20 mm, HW 0.29 mm; PL 0.35 mm, PW 0.33 mm; EL 0.56 mm, EW 0.56 mm; A1-6 respectively 0.02, 0.16, 0.15, 0.12, 0.11, 0.3 mm. An1–11 respectively 0.10, 0.05, 0.03, 0.02, 0.02, 0.02, 0.03, 0.03, 0.04, 0.04, 0.11 mm. MP1–4, respectively, 0.01, 0.05, 0.01, 0.05 mm. GL 0.23 mm, GW 0.14 mm. ML 1.63 mm. Integument: Typical for genus. Head: Head slightly narrower than pronotum. Eyes present and well-developed with 27 facets, slightly emarginate posteriorly, ocular canthus weakly developed, projecting weakly laterally and sloping to gula. Ventral surface flat to slightly rounded at gula, gular sulcus well-developed anteriorly and projecting to margin of posteriorly located, paired, gular foveae. Thorax: Prothorax with basolateral margin slightly crenulate, median longitudinal sulcus present, area behind basal sulcus with thin, longitudinal sulcus extending from basal bead to basal sulcus. Prosternum lacking anteroprosternal foveae. Protibia with apical spine. Mesotrochanter with tubercle, mesotibia with apical flange. Metaventrite slightly concave baso-medially. Elytra bearing 3 basal foveae, as well as subhumeral foveae. Flight wings well-developed. Abdomen: Tergites unmodified. Seventh ventrite broadly oval. Other ventrites unmodified. Genitalia: Aedeagus asymmetrical. Broad, membranous lobe dorsal and associated with left paramere that is twisted counter-clockwise distally and bears a row of ctenidial spines on both sides. Median lobe with distal fimbriated process. Right paramere more ventral, flattened, distally expanded to left, with apical patch of very sparse, scattered, short, suberect setae.
FEMALE. Unknown.
Variation. Unknown.
Bionomics. Seasonality: The type was collected during May. Elevation: The elevation at the type locality is 646 m. Habitat/Microhabitat: The holotype was collected from oak and rhododendron litter. Collecting methods: The holotype was collected by sifting forest litter and extracting via a Berlese funnel.
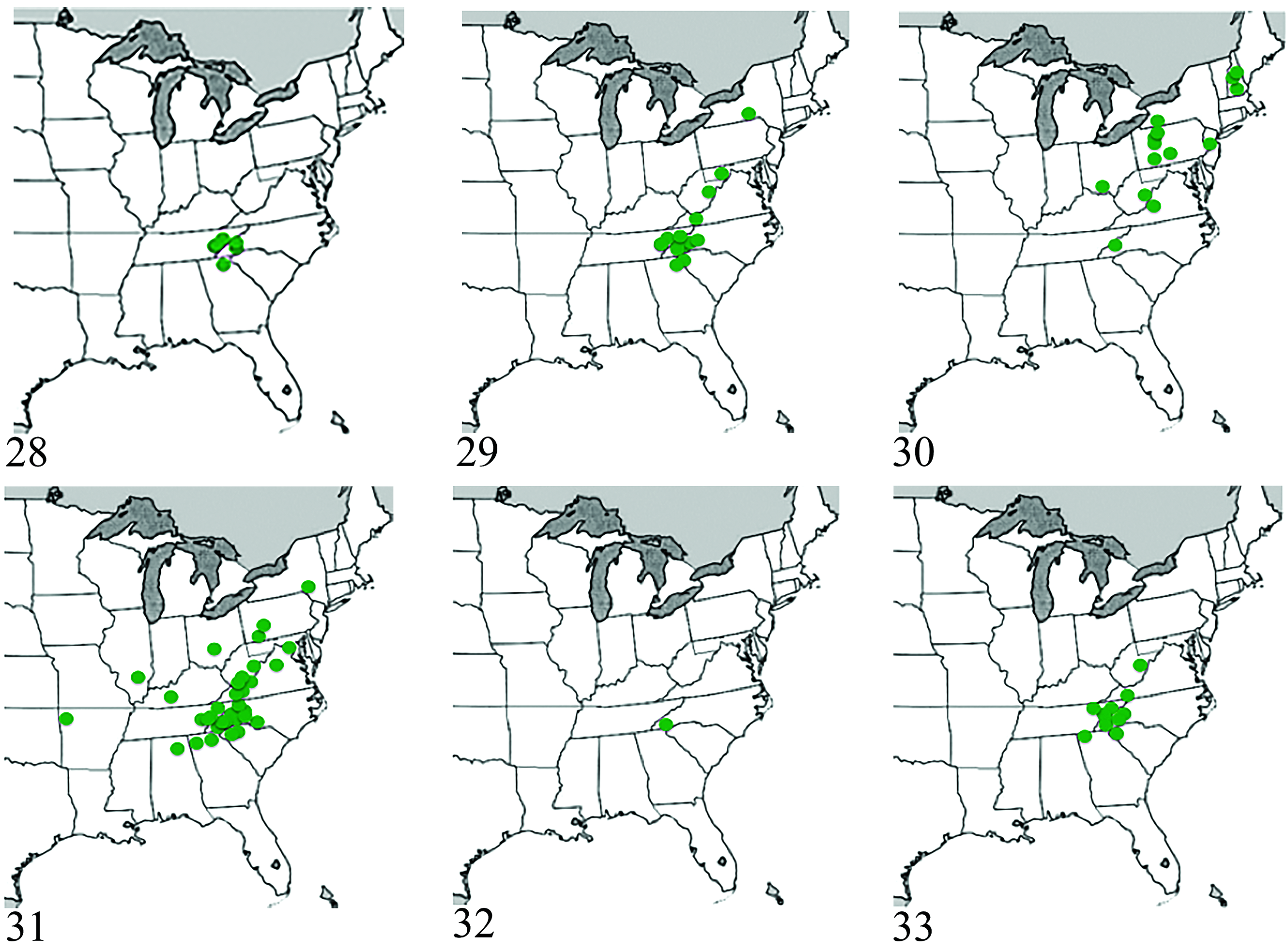
Distribution. This species is known from Macon County, North Carolina ( Fig. 32 View Figs ).
6. Eutyphlus thoracicus Park, 1956 ( Figs. 11, 12 View Figs , 18 View Figs , 33 View Figs )
Diagnosis. Eutyphlus thoracicus is distinguished from all known Eutyphlus species by the smaller body size, absence of a longitudinal pronotal sulcus, the left paramere of the aedeagus forming a sclerotized bar that bears small spicules and setae, and the ventrally located flattened right paramere ( Fig. 18 View Figs ).
Redescription. MALE. Measurements: HL 0.14 mm, HW 0.20 mm; PL 0.21 mm, PW 0.20 mm; EL 0.34 mm, EW 0.38 mm; A1–6 respectively 0.03, 0.11, 0.10, 0.09, 0.09, 0.02 mm. An1–11 respectively 0.05, 0.03, 0.01, 0.01, 0.01, 0.01, 0.02, 0.02, 0.02, 0.03, 0.09 mm. MP1–4 respectively 0.01, 0.03, 0.01, 0.03 mm. GL 0.05 mm, GW 0.04 mm. ML 1.20 mm. Integument: Typical for genus. Capitate genal setae sparse. Head: About as wide as pronotum. Vertex bearing small, longitudinal depression extending dorsally from posterior margin of head to about the margin of the eye. Eyes present, well-developed with 27 facets, convex, about as long as wide. Ocular canthus weakly developed, projecting laterally and sloping to gula. Ventral surface of head rounded at gula, gular carina or sulcus absent or weakly developed anteriorly, posteriorly located paired, gular foveae present. Thorax: Pronotum with lateral margin smooth, median longitudinal sulcus absent, area behind basal sulcus smooth. Prosternum bearing anteroprosternal foveae. Paired median mesoventral fovea present. Metaventrite flat. Elytra bearing 2 basal foveae as well as subhumeral foveae. Flight wings well-developed. Abdomen: Tergites unmodified. Fourth ventrite bearing small basolateral foveae. Seventh ventrite small and strap-like. Other ventrites unmodified. Genitalia: Aedeagus asymmetrical. Left paramere most dorsal, oblique, forming a sclerotized bar bearing small spicules and setae, and possessing a single, elongate, median seta. Median lobe modified into internal, fimbriated process associated with internal, membranous sac. Right paramere attached ventrally, flattened, and bearing 4 distal setae.
FEMALE. Measurements: HL 0.16 mm, HW 0.16 mm; PL 0.23 mm, PW 0.21 mm; EL 0.30 mm, EW 0.29 mm; A1–6 respectively 0.02, 0.11, 0.10, 0.10, 0.08, 0.04 mm. An1–11 respectively 0.04, 0.02, 0.01, 0.01, 0.01, 0.01, 0.01, 0.01, 0.02, 0.03, 0.07 mm. MP1–4 respectively 0.01, 0.03, 0.01, 0.03 mm. ML 1.20 mm. Integument, head, thorax, and abdomen: Similar to male except eyes completely absent, ocular canthus weakly developed. Tempora rounded and nearly parallel. Pronotum more quadrate. Seventh ventrite small and transverse. Genitalia: Completely membranous. Produced into somewhat elongate, median membranous structure and billowy right sac.
Variation. Although generally smaller than members of other species, size is variable (± approximately 0.1 mm total length). The degree of pronotal convexity varies slightly. The aedeagus of E. thoracicus varied little among the individuals examined, but three distal setae were present on the right, ventrally located paramere of a few specimens instead of the usual four.
Bionomics. No previously published information on the biology of E. thoracicus is available. Seasonality: Specimens of E. thoracicus were all collected from late winter/early spring to early fall (earliest date 21 February; latest 20 October). Elevation: Specimens were obtained from between 870m and 1,920 m elevation. Habitat/Microhabitat: Specimens were collected from hardwood forest litter, Rhododendron litter, oak litter, spruce litter, fir litter, and old-growth forest mixed litter. Several individuals were also collected from moss mats. Collecting methods: The majority of individuals were collected via litter sifting and Berlese extraction. Several individuals were collected in flight intercept traps.
Distribution. The species is known from Georgia (Dade County), North Carolina (Haywood, Macon, Sway, and Yancy Counties), Tennessee (Blount, Cocke, and Sevier Counties), Virginia (Washington County), and West Virginia (Pocahontas County) ( Fig. 33 View Figs ).
PHYLOGENETIC ANALYSIS
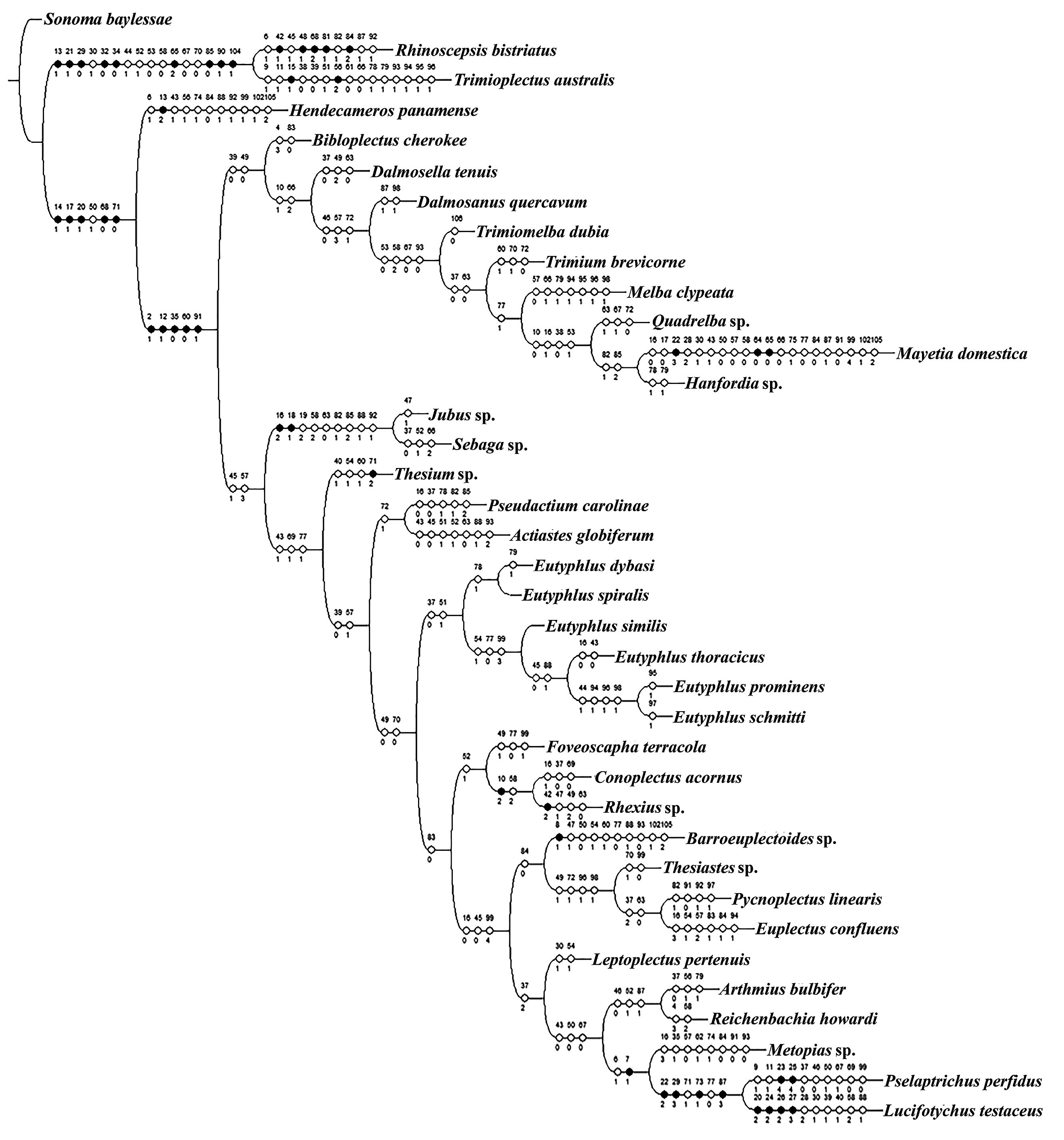
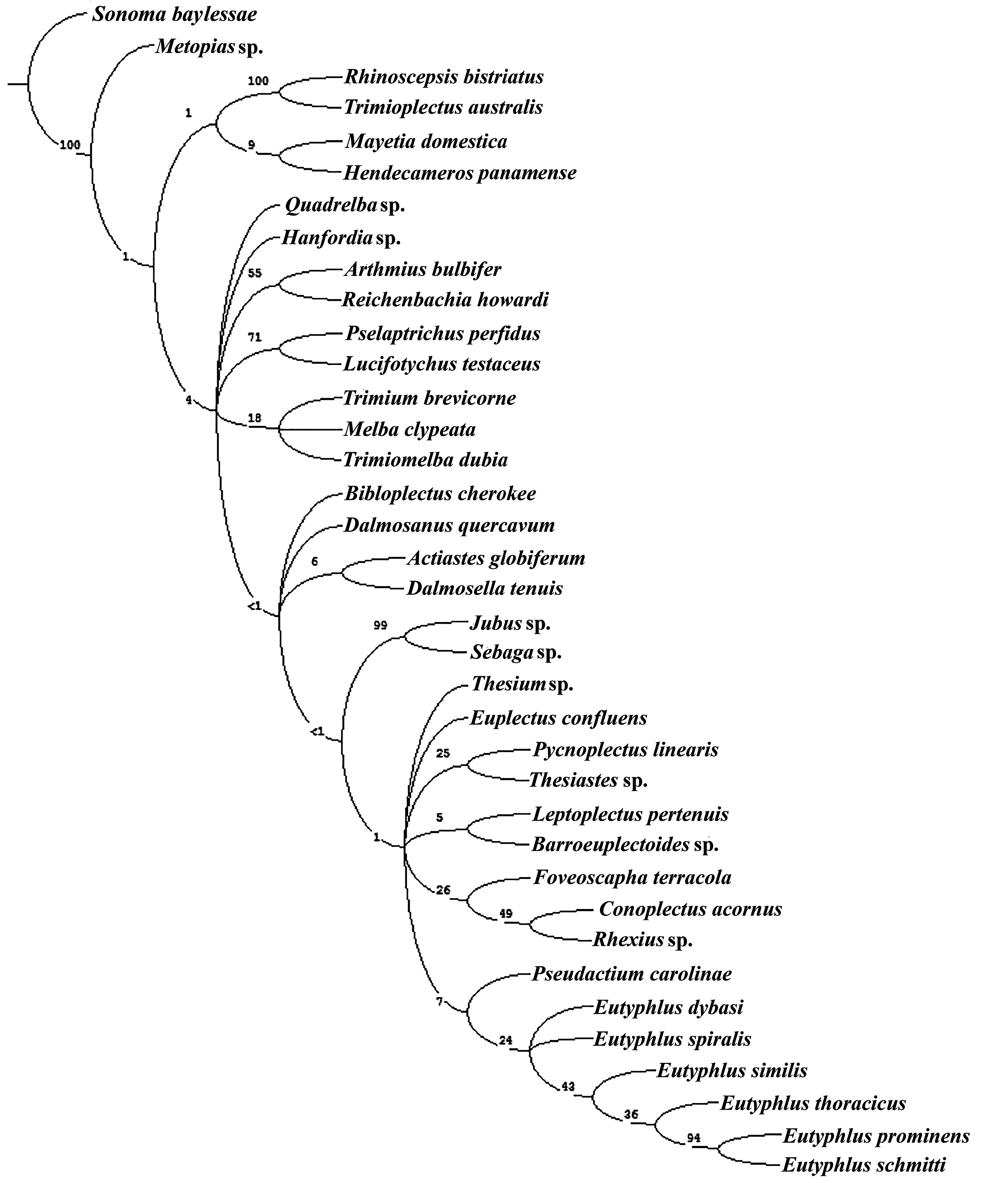
Results. Cladistic analysis of the data matrix resulted in 26 equally parsimonious trees (length = 347, CI = 0.37, RI = 0.52). The preferred tree selected for illustration ( Fig. 34 View Fig ) was rooted with Sonoma baylessae Ferro and Carlton (tribe Faronini ). A clade comprising one metopiasine genus, Rhinoscepsis LeConte , and one panaphantine genus, Trimioplectus Brendel , were recovered as sister to the remaining taxa. Remaining taxa were supported by five synapomorphies (14-1, 17-1, 20-1, 68-0, 71-0) and one homoplasious character (50-1). Hendecameros Comellini , the single representative of the tribe Bythinoplectini , diverged based on a number of homoplasies and a single autapomorphy (13-2). The clade containing the remaining taxa was supported by five synapomorphies (2-1, 12-1, 35-0, 60-0, 91-1). Two homoplasies (39-0, 49-0) supported a clade comprising all representatives of the Trimiina , plus members of the genus Bibloplectus Reitter (subtribe Panaphantina ) and Mayetiini . Bibloplectus was sister to Trimiina , and Mayetia Mulsant and Rey was nested within the trimiine taxa close to the genus Hanfordia Park. The second branch, linked by the homoplasies (45-1, 57-3), contained Eutyphlus and a number of trichonychine genera and genera from the remaining Euplectitae tribes and supertribes, with the exception of the basal Faronitae . The two genera of the tribe Jubini were recovered as sister taxa based on a number of synapomorphies and homoplasies. Thesium Casey was not recovered with other panaphantines. Actiastes Casey was not recovered with the other trimiine genera but was paired with Pseudactium Casey.
Eutyphlus was recovered as a monophyletic group alongside other members of the Panaphantina , excluding Thesium , based on two homoplasies. Within the genus, E. dybasi and E. spiralis formed a sister clade to the other four species based on a single character (78-1). The remaining species were grouped as E. similis sister to E. thoracicus and E. schmitti sister to E. prominens .
Other clades included a mixture of trichonychine genera on trees that differed in topology among various equally parsimonious trees. Non-trichonychine genera generally segregated on the trees with batrisite and goniacerite genera as sister taxa, and the two representatives of the goniacerite tribe Tychini recovered as sister taxa.
Bootstrap support was low for many of the internal branches of the tree ( Fig. 35 View Fig ). Four branches on the tree had a branch support above 90%. The branch with Sonoma Casey diverging as sister to all other taxa, the Rhinoscepsis - Trimioplectus clade, the Jubini clade, and the branch linking E. prominens and E. schmitti had branch supports of 1.0, 1.0, 0.99, and 0.94, respectively. The next highest branch support was 0.71 for the goniacerite clade containing Pselaptrichus Brendel and Lucifotychus Park and Wagner. The branch containing Arthmius LeConte and Reichenbachia Leach had support of 0.55. The rest of the branches in the tree had support below 0.50.
Discussion. The main objective of the phylogenetic analysis was to test for the monophyly of Eutyphlus . Although branch support was low, monophyly of the genus was supported by several homoplasious character states. These results, in addition to the relatively straightforward diagnosis of the genus using easily observable external characters, argue for retaining genus status for this group of species.
Several tribes and subtribes were recovered as paraphyletic among different branches of the tree. Only the Bythinoplectini , Jubini , Trogastrini , and Dimerini formed monophyletic groups, and only the Bythinoplectini and Jubini were not nested within other tribes. Limited taxon sampling of these genus groups in this analysis makes assertions regarding monophyly problematic.
The supertribe Euplectitae is a collection of tribes and subtribes that have been taxonomically defined on the basis of regional treatments, with previously untreated genera sometimes placed into these taxa arbitrarily ( Newton and Chandler 1989). The subjective nature of these higher classifications has resulted in downstream issues for the classification of large genus groups that share a number of dis- tinct morphological features, yet cannot be separated in a consistent manner under the current system. In addition, a number of characters exhibited by the Euplectitae , such as foveae and sulci, have proven to be highly variable within tribes and supertribes, sometimes differing among species within the same genus. This results in a large number of homoplasious characters and requires inclusion of a large number of outgroups within the Euplectitae and other supertribes to polarize characters for tree rooting.
Homoplasy is a particularly vexing problem with the external characters available for use in morphological phylogenetic analysis, and this may reflect conserved developmental genes that may be turned on and off in phylogenetically remote taxa. Thus, these characters are poor indicators of relationship at genus group nodes (tribe and subtribe), but may provide useful data for defining phylogenetically shallow nodes within genera, as required in the present study for testing monophyly of Eutyphlus . The low support values at the majority of nodes do not inspire confidence in the phylogenetic hypothesis proposed, but they are consistent with results of other species-level pselaphine phylogenies (e.g., Carlton 2003 that featured a 16-species polytomy of Reichenbachia species ).

| FMNH |
Field Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |