
Hyainailouros sulzeri BIEDERMANN , 1863
|
publication ID |
https://doi.org/ 10.2478/if-2017-0019 |
|
persistent identifier |
https://treatment.plazi.org/id/03FB2F7E-F405-FFD5-FF70-F99E5846FA83 |
|
treatment provided by |
Diego |
|
scientific name |
Hyainailouros sulzeri BIEDERMANN , 1863 |
| status |
|
Hyainailouros sulzeri BIEDERMANN, 1863
Text-fig. 6 View Text-fig
1965 Pterodon nyanzae sp. nov.; Savage, p. 274.
L o c a l i t y. Arrisdrift, Namibia. Morales et al. (1998b) and Morales et al. (2003) referred an M1 ( Text-fig. 5c View Text-fig ), an upper canine and a mandible with m1 and unerupted molar ( Text-fig. 6 View Text-fig ) to this species.
A g e. Basal Middle Miocene.
Hyainailouros osteothlastes ( SAVAGE, 1973)
1920 Hyaena sp. indet.; Fourtau, p. 91.
1947 Hyaenaelurus fourtaui ; Koenigswald, p. 292.
1973 Megistotherium osteothlastes ; Savage, pp. 485–486.
1989 Megistotherium osteothlastes Savage ; Rasmussen et al., p. 443. 2007 Hyainailouros bugtiensis Pilgrim ; Morlo et al., p. 147. 2010 Megistotherium osteothlastes Savage ; Lewis and
Morlo, p. 550.
H o l o t y p e. Skull (NHMUK M 26173).
T y p e l o c a l i t y. Gebel Zelten, Libya.
D i a g n o s i s. See Savage (1973).
A g e. Basal Middle Miocene.
O t h e r l o c a l i t i e s. Wadi Moghara, Egypt ( Fourtau 1920, Koenigswald 1947, Rasmussen et al. 1989, Morlo et al. 2007); Cheparawa, Muruyur Formation (ca. 14.5 Ma), Tugen Hills, Kenya (Morales and Pickford 2008); Fort Ternan, Kenya (ca. 13.7 Ma) ( Savage 1973); Bartule, Ngorora Formation, Member A (ca. 13–12.5 Ma) and Kabarsero, locality 2/10, Ngorora Formation, Member D (ca. 12 Ma), both Kenya ( Morales and Pickford 2005).
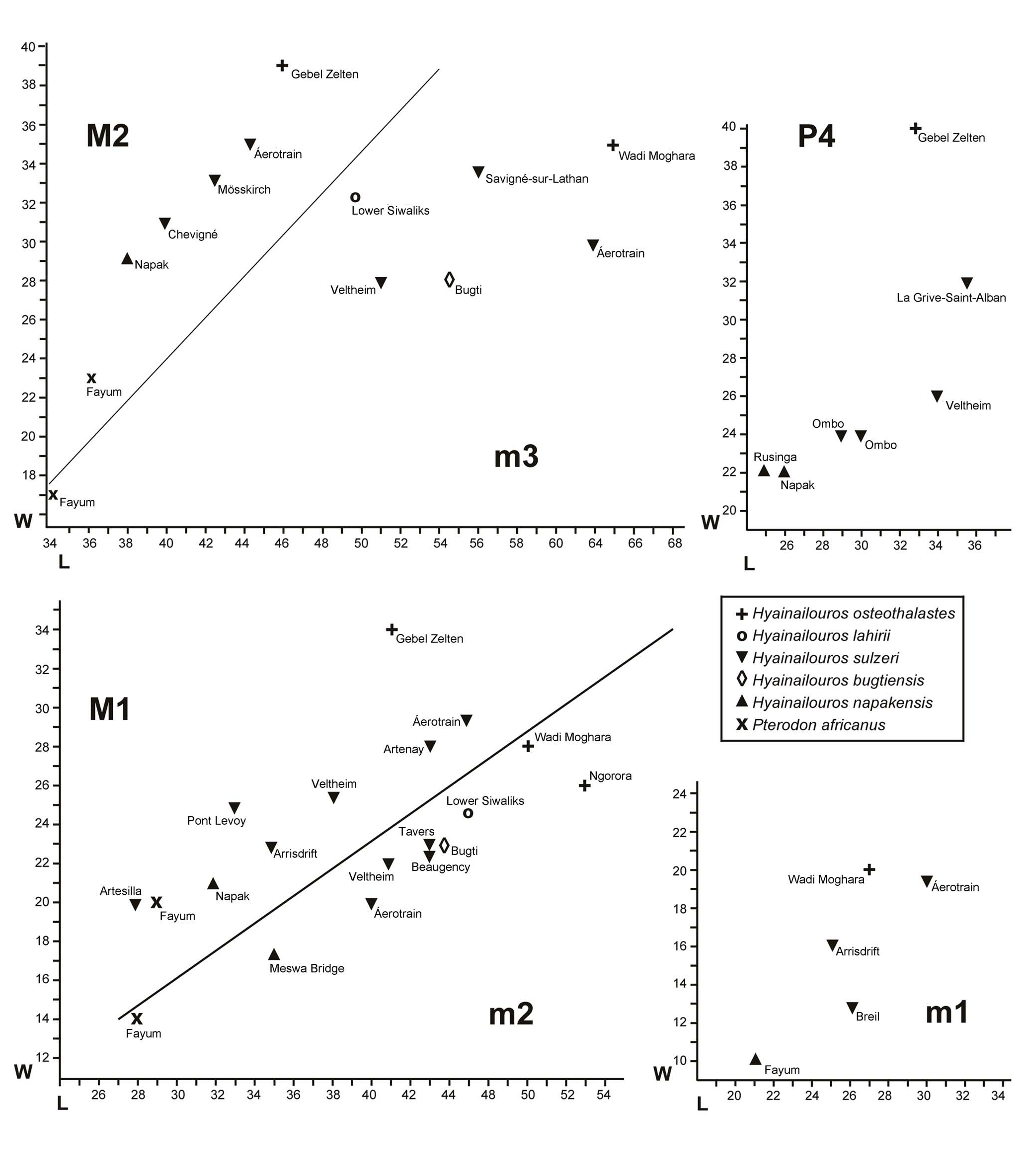
D i s c u s s i o n. Hyainailouros has a long and complicated history of study, influenced by two opposing aspects; on the one hand, it is quite rare in the fossil record, and on the other, its gigantic size makes it easily recognisable even with little material. The sample of the type species described from the Swiss locality at Veltheim by Biedermann (1863) was revised by Helbing (1925), to which he added other fossils from Europe and Asia. Later, Beaumont (1970) revised the fossils from the type locality, and fossils from Chevilly Aérotrain, France, were added to the hypodigm ( Ginsburg 1980) which are the most complete dental remains associated with elements of the postcranial skeleton. Two additional Miocene genera were attributed to Hyainailouros on the basis of their large dimensions and their resemblance to this genus; the first was Megistotherium SAVAGE, 1973 and the second was Sivapterodon erected by Ginsburg (1980) for the species Hyainailouros lahirii PILGRIM, 1932 . The latter species is known only by a fragment of mandible with m2 – m3 close in size to Hyainailouros sulzeri . According to its creator, the genus is based on the greater reduction of the paraconid in the molars (m2 – m3). However, the two molars are poorly preserved “both of them considerably worn and damaged in their anterior halves” ( Pilgrim 1932: 170) which prevents us from taking this character seriously, and the same applies to the apparent robusticity of the m3, which could have been overestimated due to the damage to the anterior part of the crown. The great reduction of the talonid of the m3, which is almost obsolete, is comparable to that in the fossils of H. sulzeri from Chevilly Aérotrain. In the current state of our knowledge, it is difficult to sustain the validity of Sivapterodon .
Megistotherium was erected by Savage (1973) for cranial and post-cranial material collected at Gebel Zelten, Libya. The holotype is a magnificent skull which unfortunately lacks most of the dentition, preserving only the almost complete right P2 and parts of the two M2s. As such, comparisons with Hyainailouros are limited. The dimensions of the molars (M1 – M2) estimated from the roots clearly exceed those of the largest specimens of Hyainailouros sulzeri ( Text-fig. 7 View Text-fig ). Despite the greater dimensions, there are no other criteria which support of generic distinction between Megistotherium osteothlastes and the type species of Hyainailouros . In addition, Ginsburg (1980) realised that some of the post-cranial elements attributed to Megistotherium by Savage (1973) belong in fact to a large species of Amphicyon . Nevertheless, the metatarsal UB 20758 ( Savage 1973: fig. 17) corresponds closely in morphology and dimensions to the specimen from Chevilly Aérotrain illustrated by Ginsburg (1980). Morlo et al. (2007) and Lewis and Morlo (2010) maintained the validity of these two genera, but unconvincingly.
The scarcity of material renders it difficult to obtain a reliable estimate of the range of metric variation of the dentition ( Text-fig. 7 View Text-fig ). The variation, with exceptions, is not very great especially when compared to other large hyaenodonts such as Pterodon dasyuroides ( Lange-Badré 1979) .Thesmallerformcomprises Hyainailourosnapakensis, plus an M1 from the Spanish site of Artesilla ( Azanza et al. 1993) and an m2 (KNM ME 12) from Meswa Bridge, Kenya Rasmussen and Gutiérrez (2009). Hyainailouros napakensis is also the oldest known species in the genus, to which can be attributed an m1 from Meswa Bridge (KNM ME 12) classified as Hyainailouros sp. by Rasmussen and Gutiérrez (2009: fig. 15C) which, in our opinion, corresponds better with an m2, the dimensions of which accord well with the Napak species ( Text-fig. 5b View Text-fig ). It is also possible that a posterior fragment of an m3 from Losodok attributed by the same authors to Hyainailourus sp. can be included in H. napakensis . The maxilla of H. napakensis is only slightly smaller than the specimen of Hyainailouros sulzeri from Arrisdrift, Namibia, comparisons between the samples being limited to the M1, which are close in morphology, with the metacone and paracone well separated from each other, with a strong parastyle and the protocone not reduced. However, two significant differences are highlighted; 1) in H. napakensis the protocone is located in a very anterior position, in front of the parastyle, whereas, in the Arrisdrift fossil, the protocone is more distally positioned in front of the protocone; 2) in H. sulzeri from Arrisdrift the sectorial part of the M1, formed of the paracone+metacone-metastyle is more compressed transversely than in H. napakensis ( Text-fig. 5c View Text-fig ). In both these features, the M1 from Arrisdrift approaches the M1 from Artesilla ( Azanza et al. 1993). Furthermore, the juvenile mandible from Arrisdrift (GSN AD 106’99) with the unerupted molar ( Morales et al. 2003) indicates the presence of a large m3, approaching 50 mm in length, a size which corresponds well with Hyainailouros sulzeri . It is notable that the locality of Artesilla has yielded mammals with clear African affinities ( Azanza et al. 1993, Pickford and Morales 1994, Morales et al. 2001).
The opposite extreme of the size spectrum is occupied by the huge species Megistotherium osteothlastes . The upper dentition is known only from the holotype skull, which, as mentioned previously, is incomplete and damaged, measurements being obtained from the roots or alveoli. However, the specimen indicates that the teeth were large, much longer than other specimens of Hyainailouros ( Text-fig. 7 View Text-fig ). Lower teeth attributed to this species were published by Rasmussen et al. (1989) and Morales and Pickford (2005) from Wadi Moghara, Egypt, and Ngorora, Kenya, respectively. The differences in dimensions from the maximal values of H. sulzeri are not great, but they fall outside the range of variation of this species. It is possible that the metric differences could correspond to extreme intra-specific variability, within which the specimens of H. osteothlastes would be the largest (perhaps males with hypertrophic canines), but it is also possible that, without denying the existence of dimorphism or great variability, there could be two or three species with overlapping dimensions. Morphological comparisons do not resolve the problems because of the meagre quality of the teeth of H. osteothlastes , impossible to observe in the upper dentition, and very limited for the lower teeth. The mandible from Wadi Moghara described by Rasmussen et al. (1989) has an m3 missing the posterior part, and an m2 with a reduced talonid, similar to the m2 of H. sulzeri and to an m2 from Ngorora ( Morales and Pickford 2005).


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Genus |
Hyainailouros sulzeri BIEDERMANN , 1863
| Morales, Jorge & Pickford, Martin 2017 |
Hyainailouros napakensis
| Ginsburg 1980 |
H. napakensis
| Ginsburg 1980 |
H. napakensis
| Ginsburg 1980 |
H. napakensis
| Ginsburg 1980 |
H. napakensis
| Ginsburg 1980 |
Megistotherium
| SAVAGE 1973 |
Megistotherium
| SAVAGE 1973 |
Hyainailouros
| BIEDERMANN 1863 |
Hyainailouros sulzeri
| BIEDERMANN 1863 |
Hyainailouros
| BIEDERMANN 1863 |
Hyainailouros sulzeri
| BIEDERMANN 1863 |
H. sulzeri
| BIEDERMANN 1863 |
Hyainailouros sulzeri
| BIEDERMANN 1863 |
Amphicyon
| Lartet 1836 |