
Jasmineiricola mackiei, Boxshall, Geoff A., O’Reilly, Myles, Sikorski, Andrey & Summerfield, Rebecca, 2015
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4018.3.6 |
|
publication LSID |
lsid:zoobank.org:pub:B60BAB3C-DA63-47BB-BBF1-FBC5B9D55EC |
|
DOI |
https://doi.org/10.5281/zenodo.6094152 |
|
persistent identifier |
https://treatment.plazi.org/id/5A07EE17-FFEE-DC5C-FF01-A1A4160CFC13 |
|
treatment provided by |
Plazi |
|
scientific name |
Jasmineiricola mackiei |
| status |
gen. nov. |
Jasmineiricola mackiei n. gen. et n. sp.
Type material. Holotype ♀ plus 4♀♀ paratypes from single specimen of Jasmineira caudata, Huldra, Stn 8-2 (60.8463ºN, 2.616804ºE), depth 123 m, 0 5 June 1999; collected by A. Sikorski; NHMUK Reg. No. Holotype ♀ 2015.447, paratypes ♀♀. 2015.448-451.
Northern North Sea. 1♀ paratype from J. caudata , North Sea, Conoco Lyell Field (60o 53' 56.62"N, 0 1o 16' 17.23"W), depth 140 m; July 1991; collected by S. Hamilton. 10♀♀ paratypes from 7 specimens of J. caudata , northern North Sea from Statfjord Field (Blocks 33/34); 1996: collected by S. Hamilton. 1♀ paratype from J.caudata, Oseberg Sør, Stn 13-3 (60.3965ºN, 2.784167ºE), depth 99 m, 0 4 May 1998; collected by A. Sikorski. 2♀♀ paratypes from 2 specimens of J. caudata, Nordøstflanken, Stn 2-2, (61.35ºN, 1.9475ºE), depth 152 m, 17 May 1998; collected by A. Sikorski. 3♀♀ paratypes from J. caudata, Nordøstflanken, Stn 1-2, (61.33333ºN, 1.9605ºE), depth 150 m, 17 May 1998; collected by A. Sikorski. 1♀ paratype from J. caudata, Nordøstflanken, Stn 6-2, (61.36917ºN, 1.942166ºE), depth 170 m, 17 May 1998; collected by A. Sikorski. 2♀♀ paratypes from 2 specimens of J. caudata, Nordøstflanken, Stn 9-2, 4, (61.361ºN, 1.92ºE), depth 153 m, 18 May 1998; collected by A. Sikorski. 1♀ paratype from J. caudata, Nordøstflanken, Stn 10-4, (61.3705ºN, 1.928666ºE), depth 161 m, 18 May 1998; collected by A. Sikorski. 1♀ paratype from J. caudata, Nordøstflanken, Stn 12-6, (61.40317ºN, 1.879166ºE), depth 161 m, 18 May 1998; collected by A. Sikorski. 1♀ paratype from J. caudata, Vigdis, Stn 9-3 (61.3782ºN, 2.104756ºE), depth 279 m, 30 May 1999; collected by A. Sikorski. 2♀♀ paratypes from J. caudata, Vigdis, Stn 15-3 (61.37692ºN, 2.094748ºE), depth 276 m, 30 May 1999; collected by A. Sikorski. 1♀ paratype from J. caudata, Tordis, Stn 4-3 (61.2751ºN, 2.120787ºE), depth 202 m, 30 May 1999; collected by A. Sikorski. 2♀♀ paratypes from 2 specimens of J. caudata, Huldra, Stn 9-2, 3, (60.8564ºN, 2.652572ºE), depth 123 m, 0 4 June 1999; collected by A. Sikorski. 1♀ paratype from J. caudata, Huldra, Stn 1-1 (60.85328ºN, 2.650849ºE), depth 123 m, 0 4 June 1999; collected by A. Sikorski. 2♀♀ paratypes from J. caudata, Huldra, Stn 11-1 (60.85974ºN, 2.664455ºE), depth 123 m, 0 4 June 1999; collected by A. Sikorski, [Specimens used for SEM]. 1♀ paratype from J. caudata, Huldra, Stn 16-1 (60.93292ºN, 2.555287ºE), depth 125 m, 0 5 June 1999; collected by A. Sikorski. 2♀♀ paratypes from J. caudata, Statfjord Øst , SFEK Stn 8-2 (61.37767ºN, 1.9095ºE), depth 156 m, 13 June 1999; collected by A. Sikorski. 3♀♀ paratypes from J. caudata, Gullfaks, Stn 1-8 (61.09441ºN, 2.19321ºE), depth 133 m, 17 June 1999; collected by A. Sikorski. 2♀♀ paratypes from J. caudata, Gullfaks, Stn 12-3 (59.2054ºN, 2.212357ºE), depth 218 m, 19 June 1999; collected by A. Sikorski, [Specimens used for SEM]. 2♀♀ paratypes from J. caudata, Regional IV, Stn 9-3 (61.12257ºN, 2.397248ºE), depth 188 m, 19 June 1999; collected by A. Sikorski, [Specimens used for SEM]. 1♀ paratype from J. caudata, Veslefrikk, Stn 7-1 (60.78652ºN, 2.9121ºE), depth 177 m, 24 May 2004; collected by A. Sikorski. 1♀ paratype from J. caudata, Veslefrikk, Stn 1 (60.7432ºN, 2.942ºE), depth 168 m, 24 May 2004; collected by A. Sikorski, [Specimen used for SEM]. 1♀ paratype from J. caudata, Vigdis F 2005 , Stn 5-3 (61.31358ºN, 2.077643ºE), depth 222 m, 14 June 2005; collected by A. Sikorski. Registration numbers NHMUK 2015.452-461.
High Latitude Norwegian waters (non-type). 2♀♀ from J. caudata, Resi Stangnes 0 6, Stn 5-1 (68º 48.459’N, 16º 36.753’E), depth 74 m, 28 June 2006; collected by A. Sikorski. 1♀ from J. caudata, Vega 2006 , Stn 2-1 (65.70004ºN, 12.133338ºE), depth 130 m, 0 4 December 2006; collected by A. Sikorski. 9♀♀ from 7 specimens of J. caudata, Mainstram F-07, Forsan Stn G4-1 (67.95466ºN, 15.626ºE), depth 33 m, 11 May 2007; collected by A. Sikorski. 2♀♀ from 2 specimens of J. caudata, Mainstram F-07, Stn 2-1 (67.95734ºN, 15.63483ºE), depth 32 m, 11 May 2007; collected by A. Sikorski. 4♀♀ from 4 specimens of J. caudata, Narvik Kom, Taraldsvik, Stn T 4 (68.45305ºN, 17.43757ºE), depth 90 m, 19 July 2009; collected by A. Sikorski. 2♀♀ from J. caudata, Ellingsen, Stn T 3-2 (67.8961ºN, 16.22328ºE), depth 166 m, 14 October 2009; collected by A. Sikorski. 1♀ from J. caudata, Oseberg Sør, Stn OSS 16 (60.61029ºN 02.777862ºE), depth 104 m, 26 May 2013; collected by A. Sikorski. 3♀♀ from 2 specimens of J. caudata, Storvika, Stn 3A (67º32.222’N, 15º17.995’E), depth 19 m, 21 May 2014; collected by A. Sikorski. 3♀♀ from 2 specimens of J. caudata, Storvika, Stn 4B (67º32.272’N, 15º18.420’E), depth 32 m, 21 May 2014; collected by A. Sikorski. Registration numbers NHMUK 2015.452-461.
Scotland (non-type). 2♀♀ from J. caudata , NW Scotland, Loch Fyne, Meall Mhor, SEPA Stn 9; depth 25 m; August 1993; collected by M.O’Reilly; NHMUK Reg. No. 2015.452-461. 1♀ from J. caudata , NW Scotland, Loch Linnhe, Gorston Stn 2, depth unknown, 7 July 2004; collected by P. Garwood; NHMUK Reg. No. 2015.452-461. 1♀ from J. caudata , NW Scotland, Loch Hourn, Stn 3a, depth unknown, 11 May 2005; collected by P. Garwood. 1♀ from J. caudata , NW Scotland, Sound of Mull, Funiary Stn 5E REF (56o 33.280’N, 0 5o 64.620’W) depth 19 m; 17 August 2006; collected by J. Hunter & S. Hamilton; NHMUK Reg. No. 2015.452-461. 1♀ from J. caudata , NW Scotland, Ullapool, Ardmair, Fish Farm Stn AC1 b; depth unknown; 29 June 2010; collected by J. Hunter/S. Hamilton. 1♀ from J. caudata , (Unico. sample 47557) NW Scotland, Western Isles, Gardline Survey 843510, Dev Site 20-MFB, depth unknown; 15 July 2010.
Ireland (non-type). 1♀ from J. caudata , Ireland, Dunmore 07-1 UCL, depth unknown, 2008; collected by P. Garwood; NHMUK Reg. No. 2015.452-461. 1♀ from J. caudata , (Unico. sample 43870) Irish Sea, CEFAS Stn G05, ADJSED, (53º 30.600’N, 05º 14.400’W), depth unknown; 2008.
Additional non-type material. 2♀♀ from 2 specimens of J. caudata , Sweden, Kosterfjord, SW of Yttre Vattenholm, depth unknown, 27 August 1986; collected by Andy Mackie (National Museum of Wales), Reg. No. NMWZ 1986.108. 1♀ from J. caudata , Sweden, Kosterfjord, SW of Yttre Vattenholm, depth unknown, 28 October 1989; collected by Andy Mackie (National Museum of Wales).
1♀ from Jasmineira sp. (posterior missing), North Sea, Osprey Oilfield, Stn 4A (61º 10’N, 01º 10’E), depth 150-180 m, 1990; collected by Brian Cleator. 2♀♀ from J. candela , Veslefrikk-98, Stn 2-2 (60.7645ºN, 2.920833ºE), depth 174 m, 16 May 1998; collected by A. Sikorski; NHMUK Reg. No. 2015.462-463. 1♀ from J. elegans , (Unico. sample 34024), Northern Ireland, Belfast Lough, NIEA Stn SDCS, (54o 50.526'N, 0 5o 42.852'W), depth unknown; 31 March 2004; collected by Tim Mackie. 1♀from J. elegans , (Unico. sample 46849), North Sea, Arundel/Farragon Oilfield, Gardline Survey GDLARUFA10, Stn A09-24-a, depth unknown, 0 9 September 2009. 1♀ damaged, ectosoma broken off from host Jasmineira sp. fragment; (Unico. sample 40611); North Sea, Brent Oilfield, Gardline Survey GDL 211290, Stn BRA GR11FA, depth 140 m; 17 April 2007. 2♀♀ from Jasmineira sp. fragment, (Unico. sample 41005), North Sea, Brent Oilfield, Gardline Survey GDL 211290, Stn BRA GR18FA, depth 141 m; 20 June 2007. 1♀ on Jasmineira sp., NW Scotland, Little Loch Broom, Ardessie Fish Farm site ABC 2b, depth unknown, 26 December 2010; collected by J. Hunter/S. Hamilton. 1♀ on Jasmineira sp., NW Scotland, Little Loch Broom, Ardessie Fish Farm site AA 1b, depth unknown, 12 July 2011; collected by J. Hunter/S. Hamilton. 1♀ from Jasmineira sp. fragment, (Unico. sample 43872) Irish Sea, CEFAS Stn G05 ADJSED, (53º 28.800’N 05° 16.800’W), depth unknown; 2008. 1♀ from Jasmineira sp. fragment, (Unico. sample 43879) Irish Sea, CEFAS Stn G18 MDAC, (53º 28.800’N 05° 16.800’W), depth unknown; 2008.
Etymology. The name of the new species honours Dr. Andy Mackie (National Museum of Wales) who found material of this parasite in 1986.
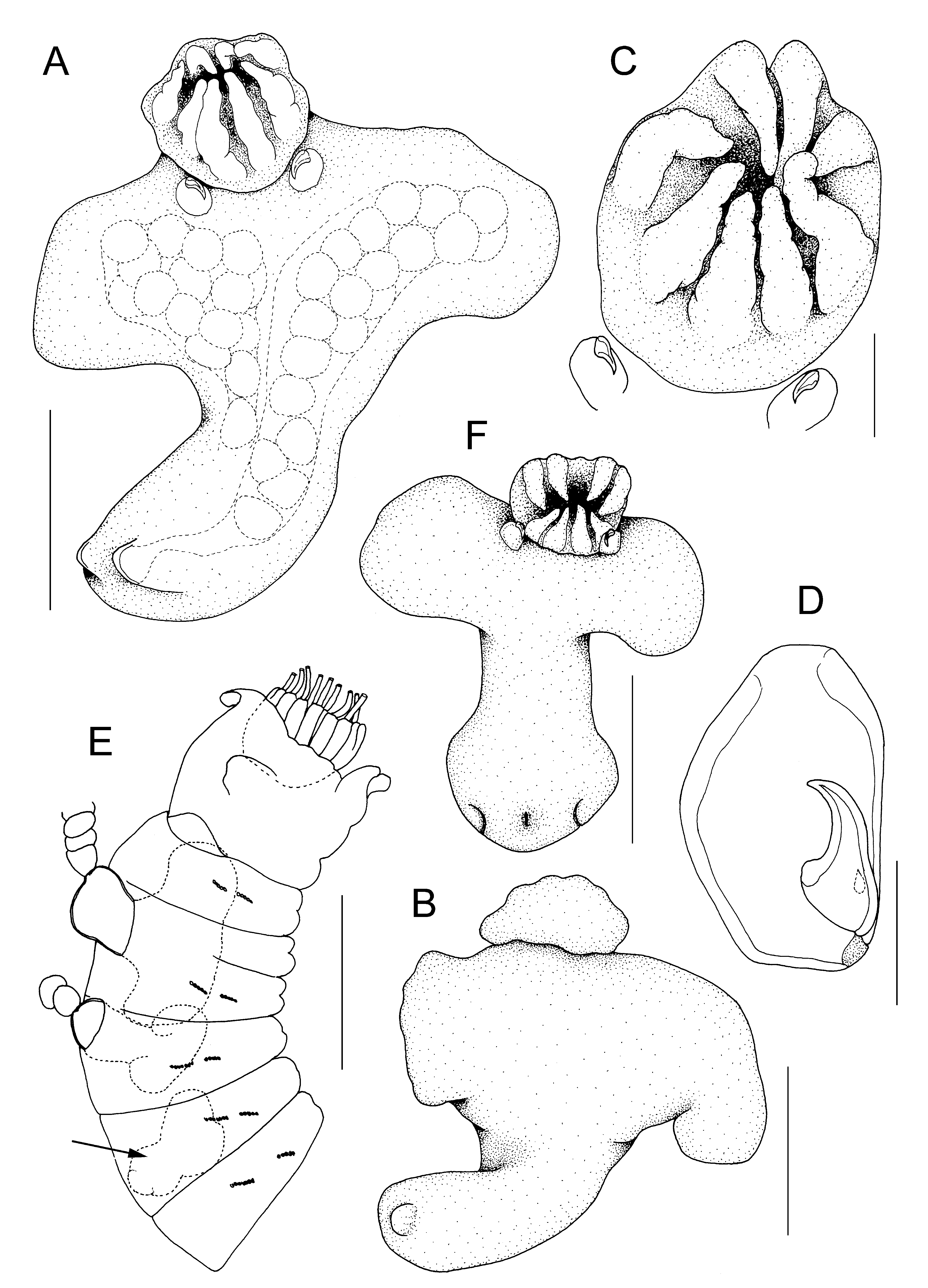
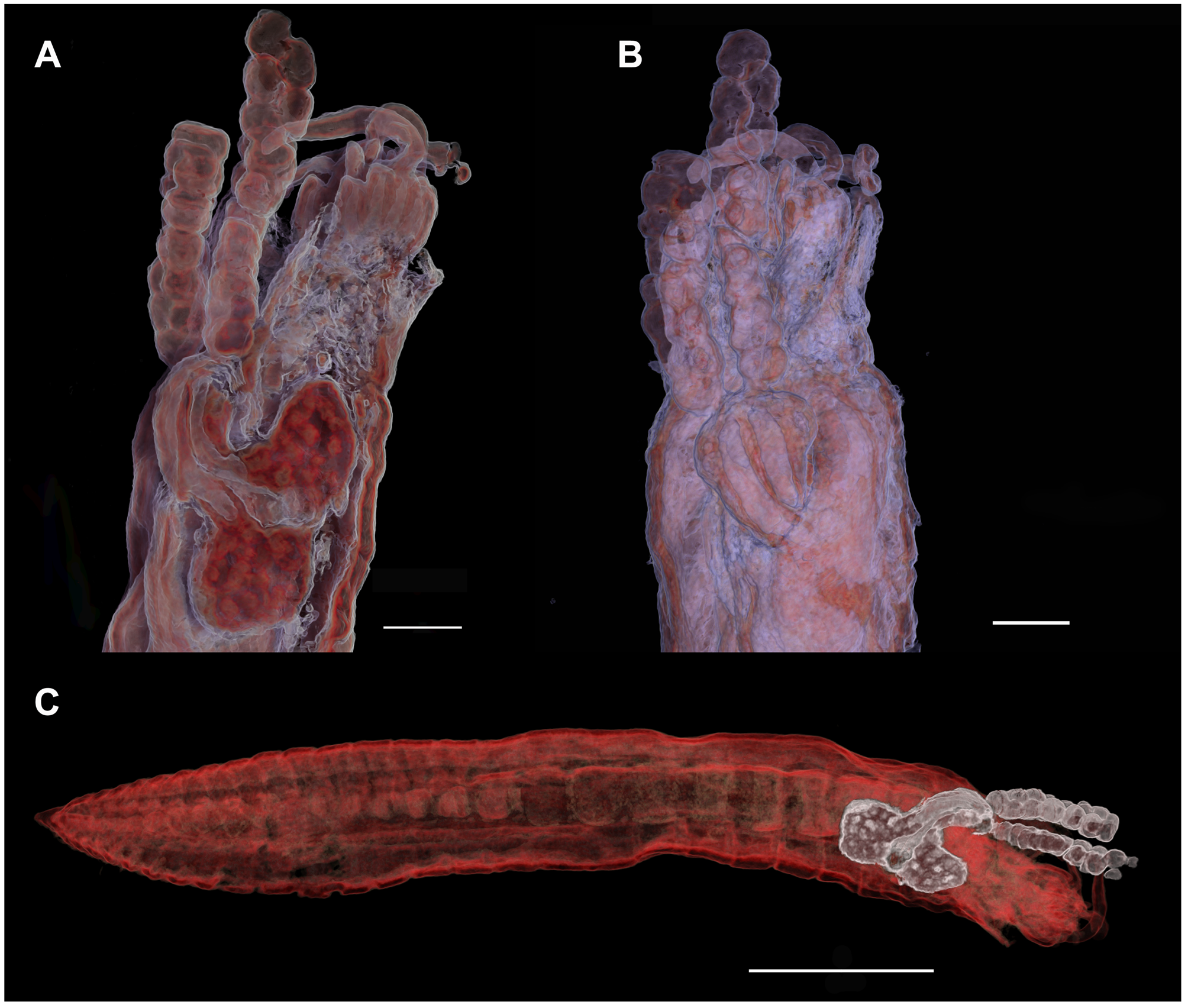
Description. The adult female is highly transformed and lacks any trace of external segmentation ( Fig. 1 View FIGURE 1 A, B). The body comprises a well defined head region carried anteriorly on the trunk which is expanded transversely to form paired anterolateral lobes and extends posteriorly to a genito-abdominal lobe. The head and anterior part of the trunk bearing the anterolateral lobes are collectively referred to as the endosoma which is embedded within the body of the host ( Figs 1 View FIGURE 1 E, 2A–C) while the posterior genitoabdominal lobe, the ectosoma, protrudes through the body wall of the host and carries the paired egg sacs ( Figs 2 View FIGURE 2 A, B, 3A, B). The adult female body exhibits torsion, twisting through 90º in the region between the endosoma and the ectosoma, so the posterior genitoabdominal lobe is directed towards the anterior end of the host ( Figs 2 View FIGURE 2 A, B, 3A, B). This torsion is not shown by developing females that have not yet erupted through the body wall of the host ( Fig. 1 View FIGURE 1 F).
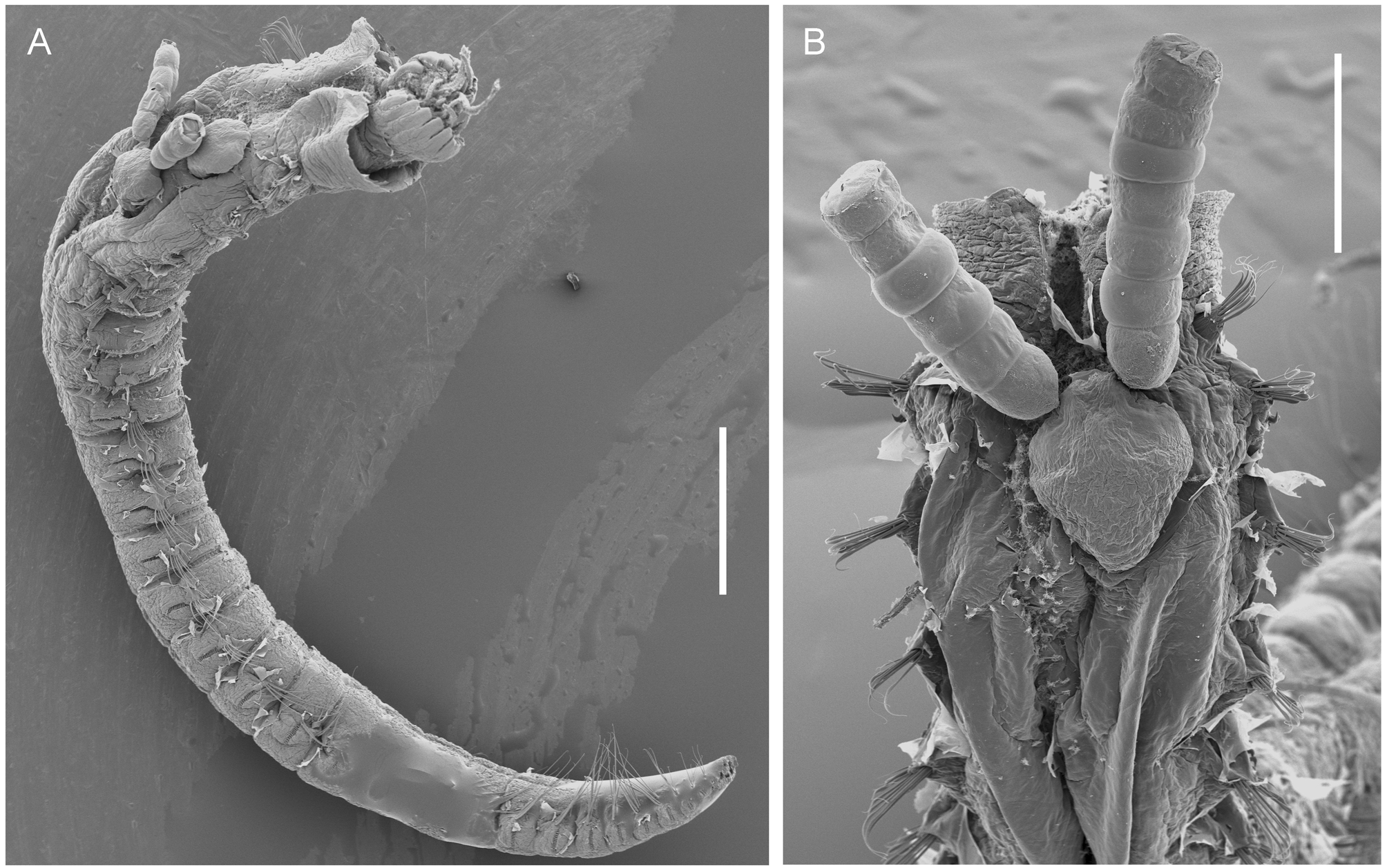
The head region is clearly defined and its ventral surface bears a rosette-like array of eight slender, tapering lobes ( Fig. 1 View FIGURE 1 C), which are arranged as four pairs and probably represent modified mouthparts. Immediately posterior to the junction of the head region and trunk are the paired maxillipeds. The maxillipeds are well developed, subchelate appendages ( Fig. 1 View FIGURE 1 D) comprising a robust proximal segment and a curved distal claw. The claw is armed with a short stout spine proximally. No trace of swimming legs was detected.
The anterolateral trunk lobes are typically dorso-ventrally flattened but are variable in shape, depending partly on their position within the host, and also on their state of development. They contain the ovaries and in mature females ( Figs 1 View FIGURE 1 A, 2A) the lobes are relatively larger than in developing females ( Fig. 1 View FIGURE 1 F). The lobes can be symmetrical or asymmetrical.
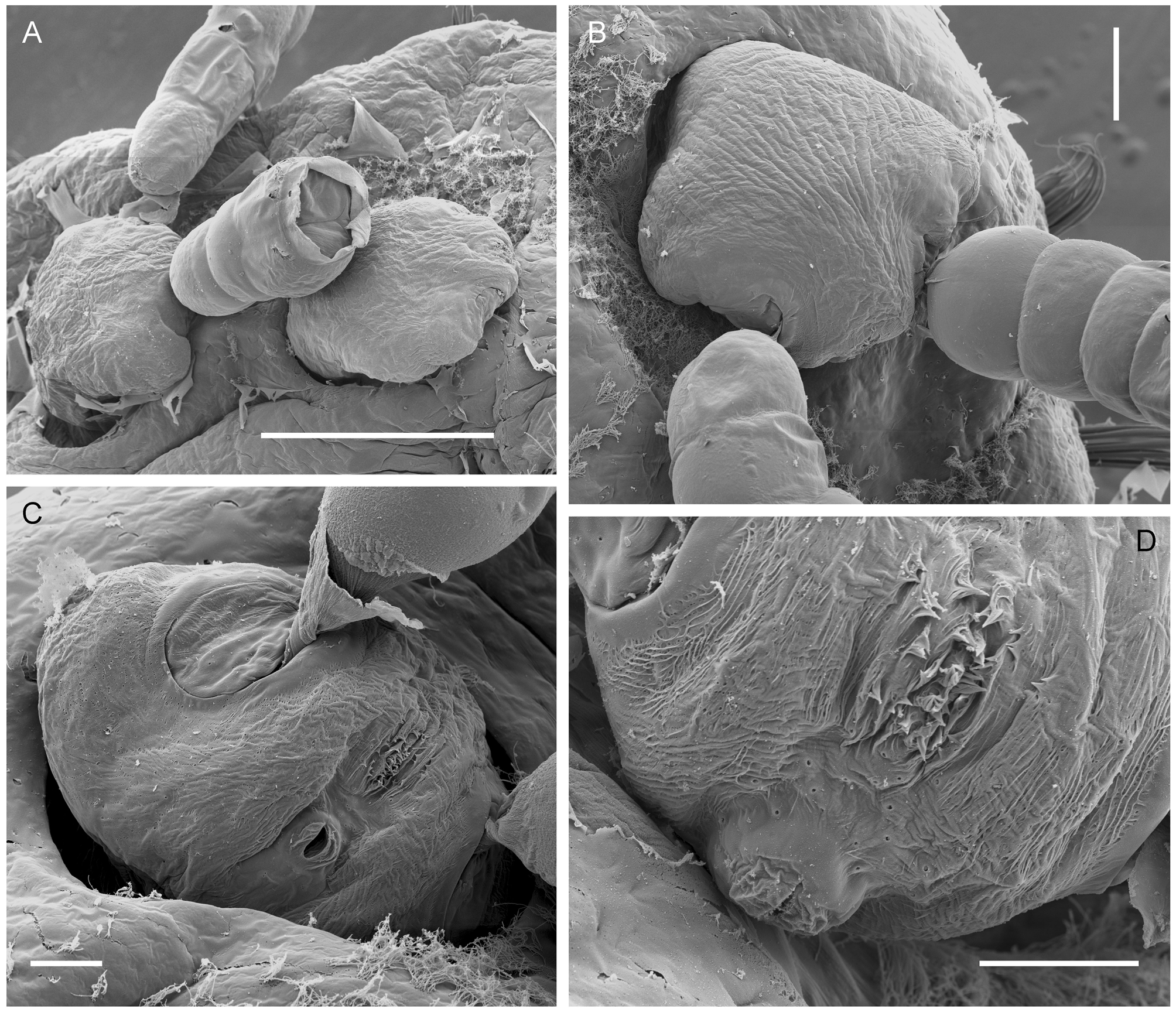
The posterior genitoabdominal lobe (= ectosoma) visible externally on the host ( Fig. 4 View FIGURE 4 A–D) is a rounded diamond-shape, approximately 180 Μm long by 190 Μm wide, and carries paired genital apertures posterolaterally and a median anus posteriorly ( Fig. 4 View FIGURE 4 C, D). Each genital aperture is rounded and closed off by an unarmed genital operculum ( Fig. 4 View FIGURE 4 C). The anus is slit-like and carried on a small anal prominence ( Fig. 4 View FIGURE 4 D). The integument of the dorsal surface is slightly ridged and in the mid-line, dorsal to the anal prominence, there is a patch where the epicuticular ridges are raised into spiniform microstructures ( Fig. 4 View FIGURE 4 D). Around the base of the anal prominence several paired integumental pores are present. No vestiges of caudal rami were found. The ectosoma contains paired cement glands which extend through into the endosoma ( Fig. 2 View FIGURE 2 B).
The egg sacs are paired and the arrangement of eggs within the sac may be linear, biseriate, or multiseriate: the mean number of eggs per sac was 24.3 (±10.2), with a range of 11 to 47 (N = 15). The egg sacs are often uniseriate near their origin at the female gonopore but most sacs contain two ( Fig. 2 View FIGURE 2 B) or more irregular rows of eggs. The maximum number of eggs observed in a single sac was 47, and this sac had four irregular rows of eggs over much of its length.
Male. Unknown.
Hosts. The parasite is specific to the sabellid genus Jasmineira and has been reported from three host species. The majority of records are from the type host J. caudata Langerhans, 1880 , but it was also found on J. candela ( Grube, 1863) in Norwegian waters and on J. elegans Saint-Joseph, 1894 in UK waters.
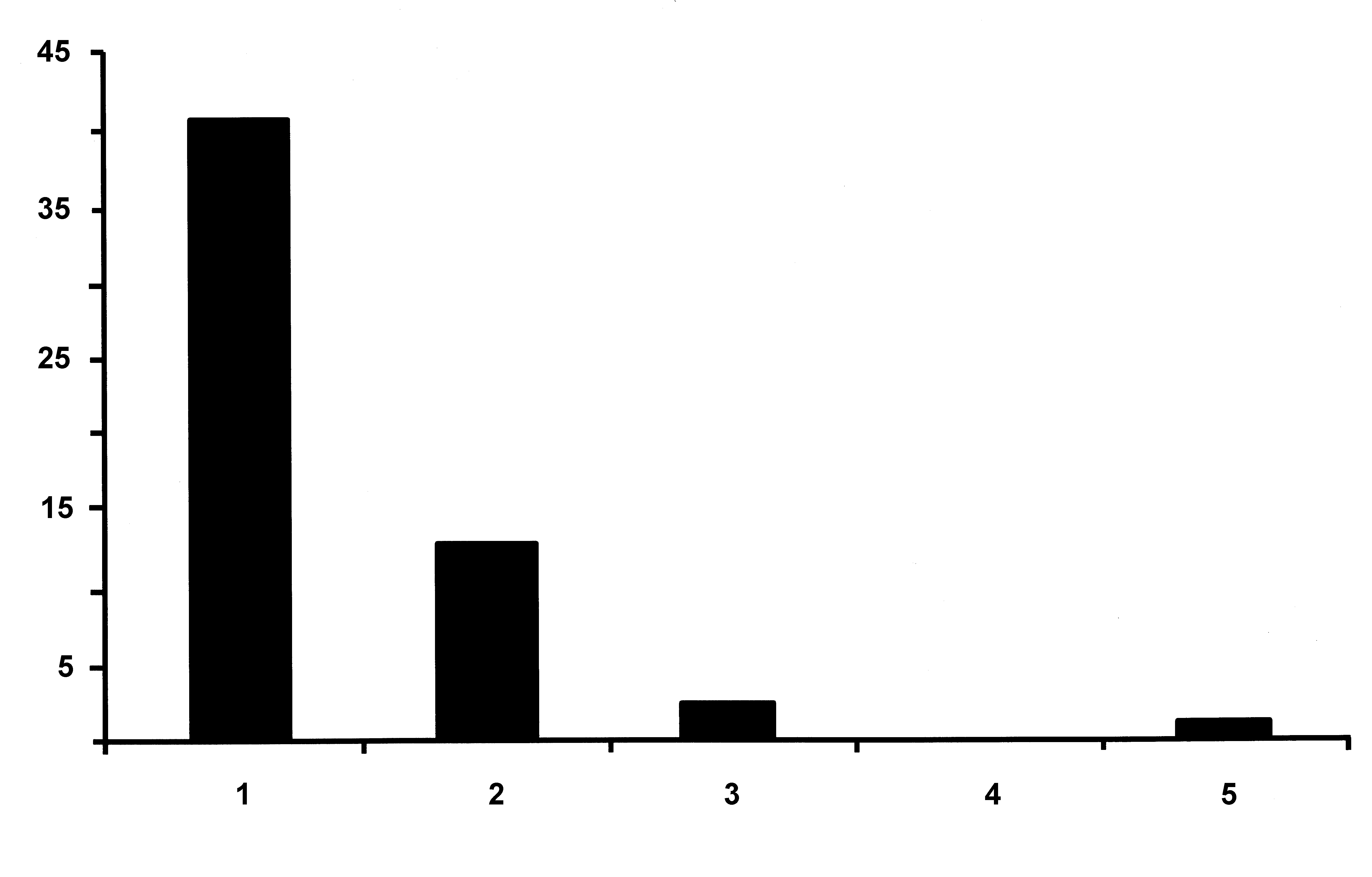
Prevalence and intensity of infection. The number of parasites present on 58 infected J. caudata was 81, giving a mean intensity of 1.40 parasites per worm, and the number of adult females per infected host ranged from one to five ( Fig. 5 View FIGURE 5 ). The maximum number of adult females found on a single host was five: on this maximally infected host three females were positioned on the left side of the worm ( Fig. 1 View FIGURE 1 E), one each in setigers 2, 3 and 4, and two females were on the right side in setigers 2 and 3. The ectosoma of the female in setiger 2 was erupted further than that of the female on setiger 3, and the female on setiger 4 was only detected when the worm was cleared in lactic acid because it had not yet erupted through the body wall of the worm. This young female ( Fig. 1 View FIGURE 1 F) did not exhibit the torsion in the region between the endosoma and ectosoma. We infer from this that an earlier infective stage in the life cycle must have penetrated the host, commenced metamorphosis within the host, and that the ectosoma erupts through the body wall of the host as the developing female approaches maturity.
Few data are available on the prevalence rate of Jasmineiricola mackiei n. gen. et n. sp., as the numbers of uninfected hosts in the samples are generally unavailable, however in one sample from Loch Fyne in Scotland, just a single J. caudata was infected out of a total of 11 examined, a prevalence rate of 9.1%. The discovery of an infected host containing a parasite that had not erupted through the body wall, indicates that such early stages might be difficult to detect and that prevalence rates might be underestimated.
Position on host. Post-metamorphic adult females are embedded in the anterior part of the host, usually on setiger 2 ( Figs 1 View FIGURE 1 E, 2B, 3A–B). Viewed from the outside the posterior end of the ectosoma is directed towards the anterior end (head) of the host. The egg sacs are therefore directed towards the distal opening of the host’s tube ( Fig. 3 View FIGURE 3 A–B) and lie within the space between the worm and its tube. When multiple infections occur, one or more of the specimens may be positioned more posteriorly, on setigers 3 or 4, but the typical position is on setiger 2.
Geographical and depth distribution. Records are reported here from numerous localities off the northwestern coast of Europe extending northwards from the Irish Sea and the sea lochs on the west coast of Scotland, through the northern North Sea, as far east as the Swedish coast at Kosterfjord, and up into Norwegian waters as far north as 68.5°N, beyond the Arctic Circle.
The known depth range extends from 19 to 279 m, and the mean depth of occurrence was 136 m, based on all records for which depth data were available (39 stations).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |