
Leucochrysa (Nodita) prisca, ENGEL & GRIMALDI, 2007
|
publication ID |
https://doi.org/ 10.1206/0003-0082(2007)3587[1:TNFODA]2.0.CO;2 |
|
persistent identifier |
https://treatment.plazi.org/id/038D8787-FFD3-FF82-FD4B-3E9CC909FE3E |
|
treatment provided by |
Carolina |
|
scientific name |
Leucochrysa (Nodita) prisca |
| status |
sp. nov. |
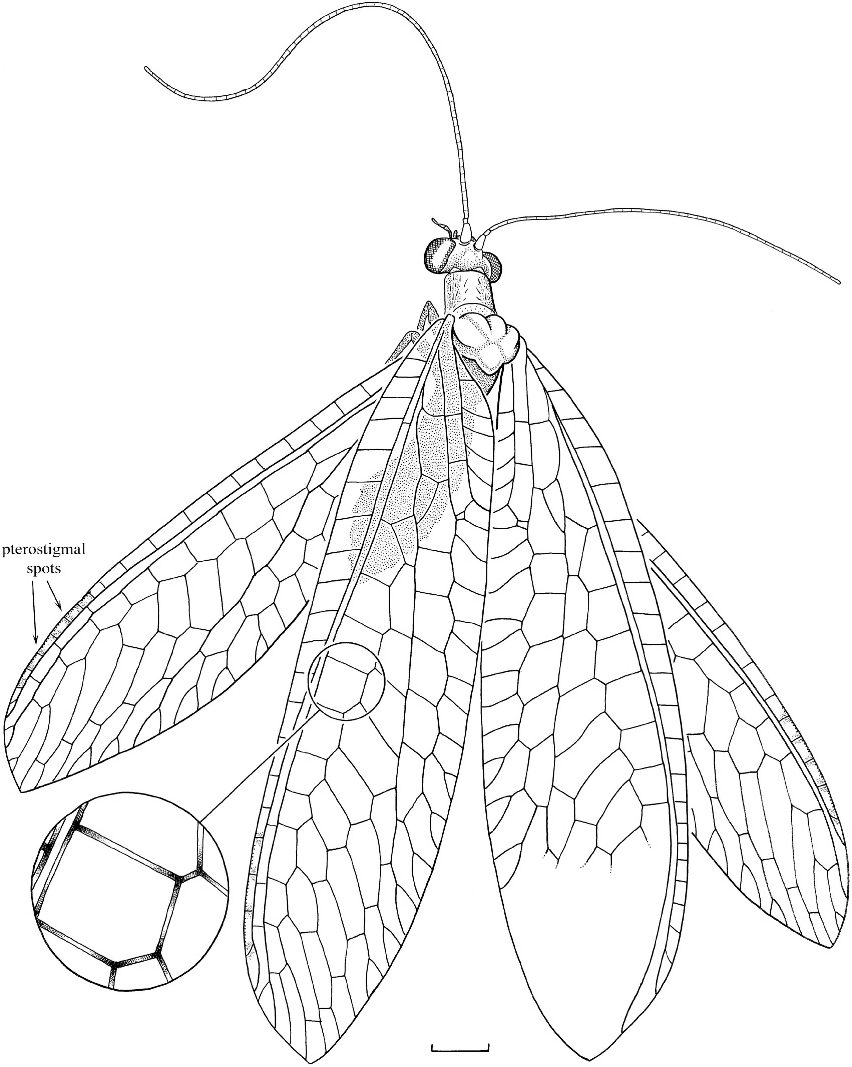
Leucochrysa (Nodita) prisca View in CoL , new species figures 32 View Fig , 33 View Fig
DIAGNOSIS: A relatively small species of Leucochrysa (at a forewing length of 10.5 mm) lacking the red or brown markings of the head.
DESCRIPTION: Forewing length 10.5 mm; total body length 6.6 mm; pronotum length 0.63 mm, width 0.73 mm. Head apparently light brown, antennae uniformly light brown, face darker brown below level of antennae, markings apparently absent; integument apparently imbricate. Palpi tapering at apices; labrum emarginate; flagellum about as long as forewing.
Pronotum apparently unmarked, slightly longer than wide, anterior margin broadly rounded; remainder of body apparently light brown, perhaps with metallic highlights.
Forewing oval and narrow, longer than abdomen, with sparse setae except along margins; 16 costal crossveins (c-sc); costal area narrows progressively toward apex; Sc and R deviate slightly near apex, Sc joins C at apex; R straight, not sinuous; single sc-r crossvein toward wing base (vein bsc), three sc-r crossveins below pterostigma; pterostigma elongate and marked with faint clouds of smoky brown; cell m1 much smaller than m2; cell im ovate, broad, first rs-m crossvein meets im subapically; no radial crossvein (r-m) basal to origin of Rs, radial crossveins straight; cell c1 as long as c2; cell dcc narrowly open along posterior wing margin, longitudinally divided anteriorly to form small cell c3; six closed cells between PsM and PsC; two gradate series, inner gradate series complete, gradates parallel; 1A forked ( fig. 33 View Fig ). Hindwing with two gradate series. Wing membranes hyaline; major longitudinal veins light brown, although darkened at points where crossveins form; crossveins distinctly darker than longitudinal veins, darker where two or more veins meet each other.
Abdomen uniformly colored; sternum two without stridulatory apparatus.
HOLOTYPE: Female ; MACT-544 ( fig. 32 View Fig ), Miocene amber of the Dominican Republic.
ETYMOLOGY: The specific epithet is taken from the Latin word priscus (meaning ‘‘old’’).
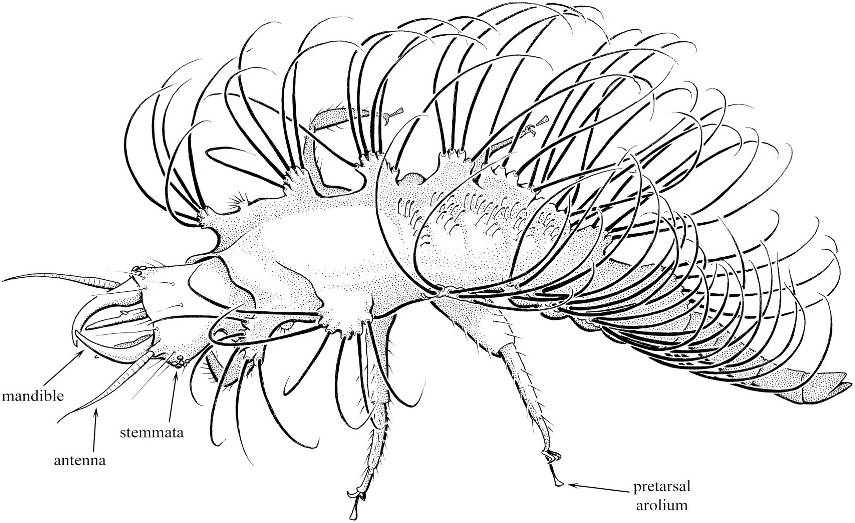
Chrysopidae larva figure 34 View Fig
Chrysopidae View in CoL larva: Grimaldi and Engel, 2005: 352,
fig. 9.31. View Fig
COMMENTS: Integument reddish-brown throughout, without apparent markings and largely lacking microtrichiae. Head distinctly flattened and roughly quadrate, slightly longer than wide; antennae longer than mandibles and slightly longer than head capsule; mandibles nearly as long as head capsule; mandibles simple, without dentition. Lateral tubercles on thorax and abdominal segments I–VII well developed ( fig. 34 View Fig ), tubercles spherical and nodulate at apices at setal origins; prothoracic segment with secondary, smaller, lateral tubercles posterior to main lateral tubercles, secondary tubercles with short, simple, erect setae; main lateral tubercles of each segment with 4–6 simple, greatly elongate setae forming a dorsal space or corbicula for the accumulation of debris on the dorsal surface of the thorax and abdomen. Thoracic and abdominal dorsa relatively flattened except slightly upcurved and flared dorsolaterally to form dorsal corbicula; dorsum without setae except metathoracic and first four abdominal segments with short, erect, setae with spatulate apices medially ( fig. 34 View Fig ). Abdomen tapering in width to relatively narrow apical segment; apical segment tapered to rough point at apex.
MATERIAL: Larva; AMNH DR- 10-1454 ( fig. 34 View Fig ), Miocene amber of the Dominican Republic.
COMMENTS: Like modern chrysopid larvae, the fossil exhibits the flattened dorsum and elongate, curved, lateral setae designed to form a dorsal corbicula. The larva fills the corbicula with debris so as to disguise itself not only from predators but from prey (the socalled ‘‘sheep in wolf’s clothing’’). Given the distribution of this behavior and such morphological adaptations across the diversity of chrysopids, it is not surprising to find a larva exhibiting this suite of features as along ago as the Miocene. Indeed, such design and behavior may stretch back into the Cretaceous.
FAMILY MYRMELEONTIDAE LATREILLE
The antlions are notable predators, familiar for the conical pits constructed by larvae of some species. Larvae dwell in the ground or on trees and it seems that eggs are laid singly, sometimes scattered in dry soil or sand. Most larvae are subterranean and some construct conical pits that are used as traps for prey. The larva lies buried at the bottom of the pit with only the open jaws protruding. In other groups, the larvae burrow freely in sand, and move under the sand or on the surface at night. Still others appear to live on trees and closely resemble bark. Larvae can often be found under overhangs where they are protected from rain. Adults are typically predatory although some are known to feed on pollen.
Several fossils of the family are known as compressions from the Early Cretaceous of Brazil and China (e.g., Martins-Neto, 1990, 1992; Martins-Neto and Vulcano, 1989a, 1989b). Many of these species have been assigned to a variety of new, extinct subfamilies but the validity of these requires careful reevaluation (e.g., the Araripeneurinae is a synonym of Palparinae ). The only other Cretaceous records of the antlions are the fossils Samsonileon fragmentus Ponomarenko from Israel (Lower Cretaceous; Dobruskina et al., 1997) and Palaeoleon ferrogeneticus Rice from Labrador (Upper Cretaceous; Rice, 1969), both sometimes placed in a separate family, Palaeoleontidae (e.g., Dobruskina et al., 1997; Martins-Neto, 2000). Additionally, Whalley (1980) reported an incomplete, unnamed specimen of a putative myrmeleontid in Lebanese amber but we have not been able to confirm its identity. Aside from the fossils discussed below, Dendroleon septemmontanus Statz is the only compression of a myrmeleontine from the Tertiary ( Statz, 1936). Only a single, definitive myrmeleontid species is known in amber from any deposit, this being the recently described Porrerus dominicanus from Dominican amber ( Poinar and Stange, 1996). Below we make minor corrections to the original description, present new figures of the holotype, and report on additional specimens.
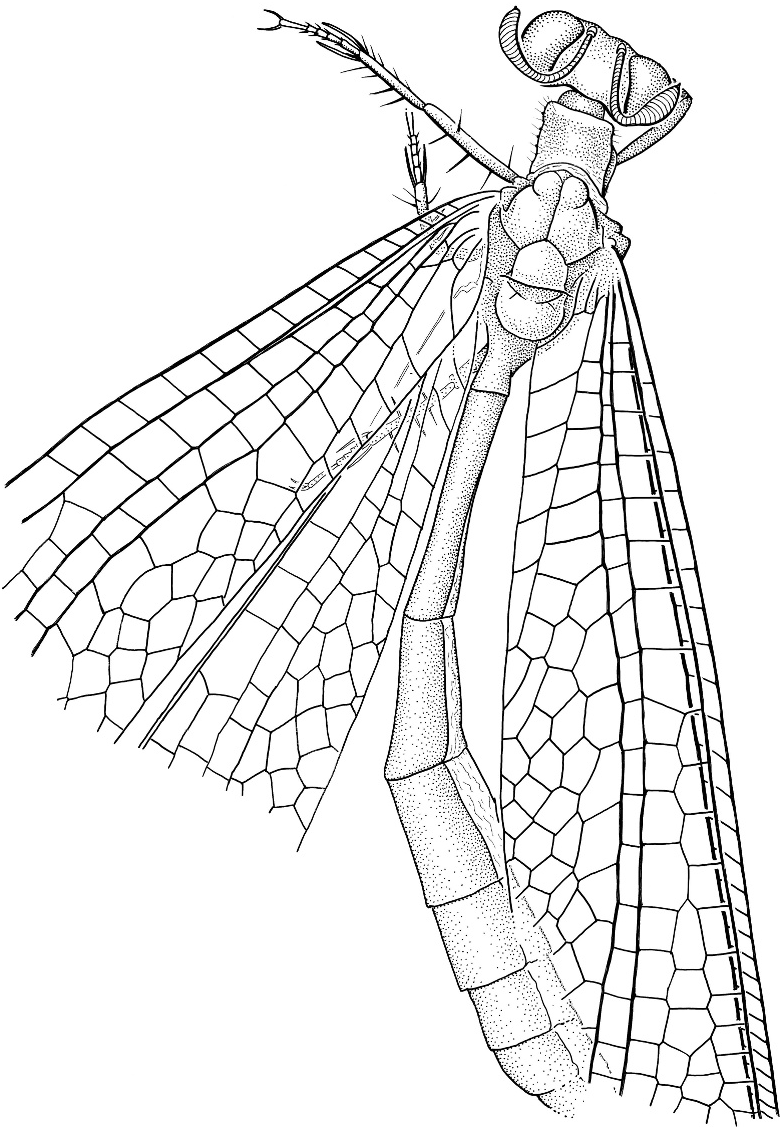
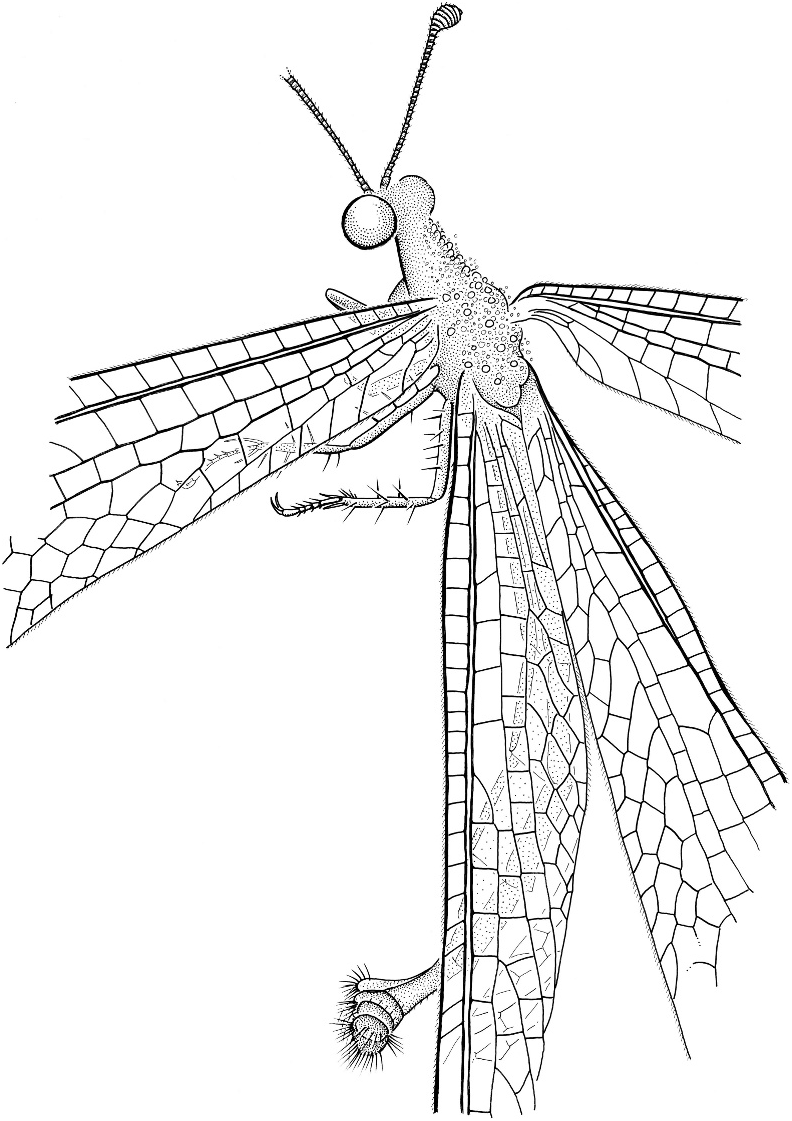
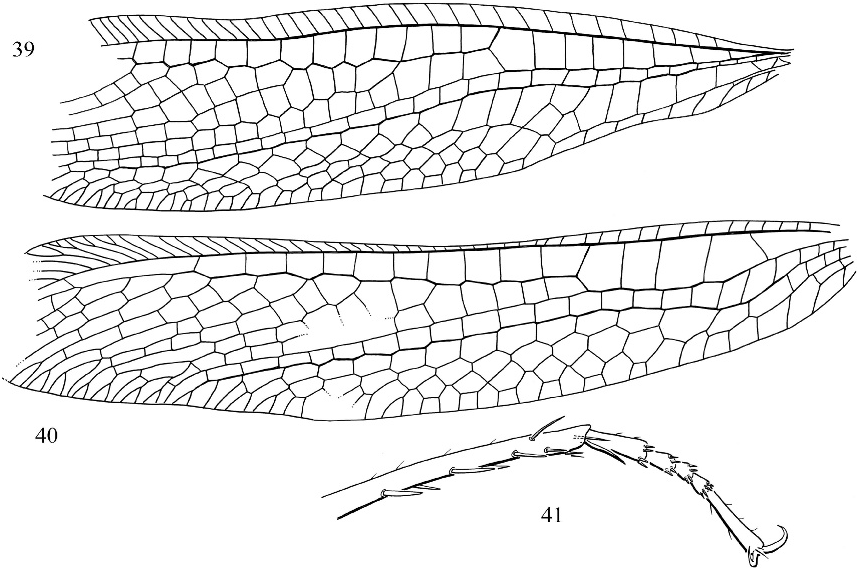
Porrerus dominicanus Poinar and Stange figures 35–41 View Fig View Fig View Fig View Fig View Figs




| AMNH |
American Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Leucochrysa (Nodita) prisca
| ENGEL, MICHAEL S. & GRIMALDI, DAVID A. 2007 |
Chrysopidae
| Grimaldi, D. & M. S. Engel 2005: 352 |