
Malacoceros jirkovi Sikorski, 1992
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3786.3.1 |
|
publication LSID |
lsid:zoobank.org:pub:3388BBBF-B3C6-4F34-9E76-E287CD335933 |
|
DOI |
https://doi.org/10.5281/zenodo.5630118 |
|
persistent identifier |
https://treatment.plazi.org/id/03D4B72B-614A-FFAB-F7D1-A116FDDC46DD |
|
treatment provided by |
Plazi |
|
scientific name |
Malacoceros jirkovi Sikorski, 1992 |
| status |
|
Malacoceros jirkovi Sikorski, 1992 View in CoL
Malacoceros jirkovi Sikorski, 1992: 105 View in CoL –108, Figure 810. Sikorski, 1994: 26–27, Figure 2 View FIGURE 2 .
Material examined. Paratype: NE Atlantic Ocean, southern Norwegian Sea: 61°00’N 1°36’W, station 1100, grab, depth 140 m, coarse sand and pebbles and shells, temperature 9.04°C, salinity 35.39 psu, 18-Jul-1957, 1 af, 1 mf (ZMUC-POL-000946).
Non– type material. Expedition DIVA 3, cruise METEOR ME 79-1, NE Atlantic Ocean, Great Meteor Seamount, 30°00.04'N 028°30.05'W, station 662, grab, depth 287.6 m, 19-Aug-2009, formalin, 1 af (ZMH- P27552).
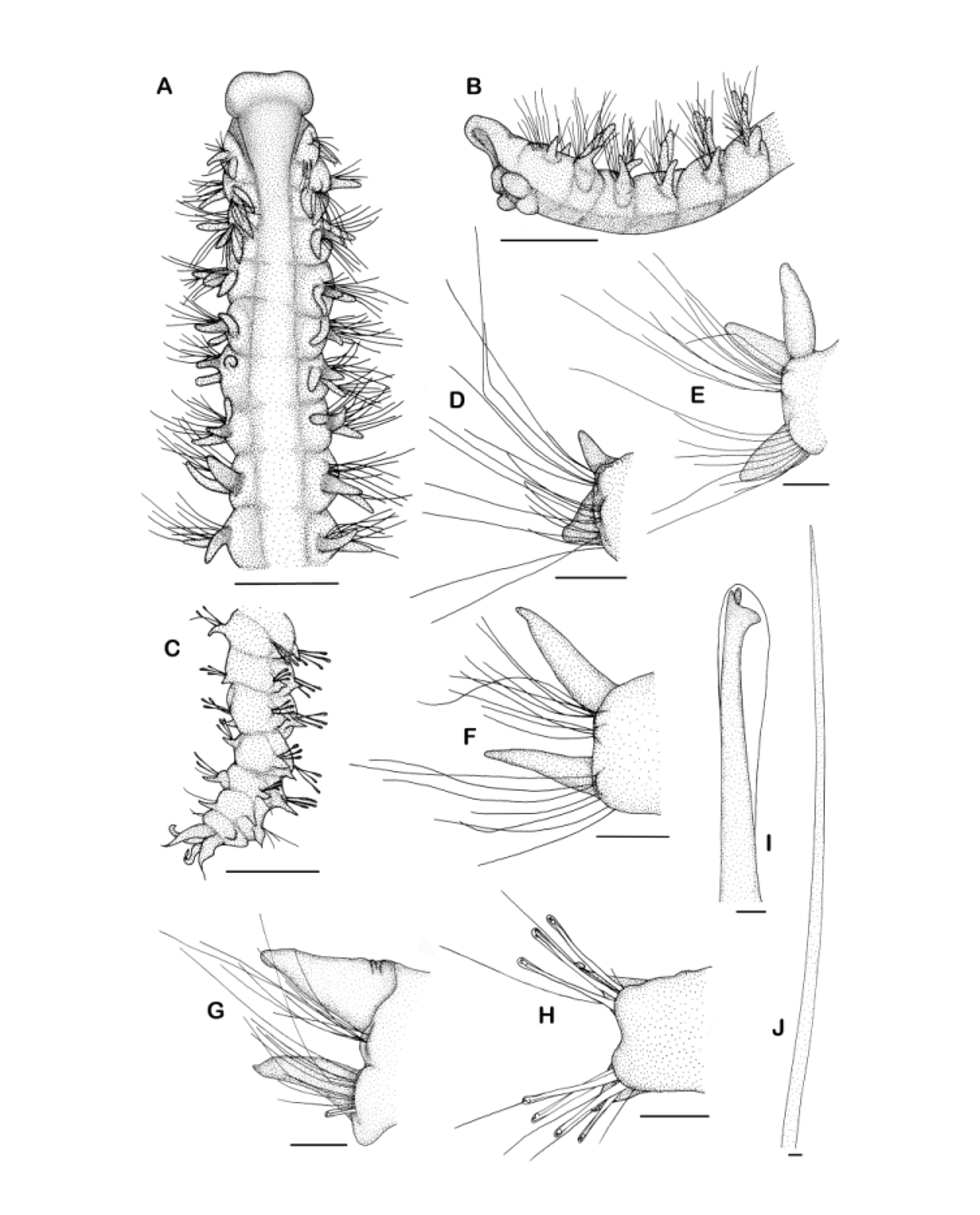
Description (of specimen from the Atlantic seamounts, expedition DIVA 3). Specimen is an anterior fragment of 41 chaetigers, 0.87 mm wide (measured at chaetiger 7), and about 8 mm long (difficult to determine because specimen curled up).
Prostomium bell-shaped to sub-triangular with distinct anterolateral horns; prostomium posteriorly extended into short caruncle which terminates at the level of the notopodia of chaetiger 2, laterally separated from peristomium by furrow. Prostomium with one pair of minute eyes positioned on the posterior half of the prostomium where the maximum width is reached, more or less at the level of the first notopodium. Occipital antenna absent. Peristomium not well developed. Palps lost, but scar of palp insertion discernable anterior to nuchal organs. Nuchal organs as small elongate grooves lateral to the posterior tip of the prostomium. Metameric dorsal ciliated organs not discernable. Dorsal crests absent but transverse ciliary bands across the dorsum present until about chaetiger 20.
Dorsal branchiae present from chaetiger 1 until the end of the fragment; cirriform with elongate, slender tip, ciliated on outer margins, separate from notopodial lamellae throughout; first branchiae about the same length as notopodial lamellae, from second chaetiger longer than notopodial lamellae, on third chaetiger reaching full length; from chaetigers 3 to ~24 branchiae almost reaching midline dorsally but not touching each other, on posterior chaetigers branchiae still distinctly longer than notopodial lamellae but thinner and shorter in comparison.
Interparapodial lateral pouches absent.
Parapodia on first chaetiger positioned slightly more dorsolaterally than on following chaetigers, there in lateral position. Notopodial lamellae subulate until about 25th chaetiger, with rounded outer margin and straight inner margin, becoming more fusiform and distinctly smaller on posterior chaetigers; neuropodial lamellae subulate and much longer than wide at first chaetiger, from second to about 25th chaetiger of rounded shape with small pointed tip on outer margin, after chaetiger 25 upper part of the lamella more strongly developed than lower part which becomes more and more reduced. Both parapodial rami without prominent praechaetal lamellae.
Chaetae in anterior and middle chaetigers capillaries with narrow sheaths, smooth but very fine granulations present near the tip, arranged in two rows, anterior row of capillaries shorter than posterior row; few very long smooth capillaries present in uppermost position in the notopodia. From chaetiger 26 posterior row of capillaries replaced by row of hooks in neuropodia, hooks hooded, bidentate or perhaps tridentate with additional minute apical teeth (difficult to observe due to size), 6–8 hooks present in each neuropodium; capillaries in anterior row of hook-bearing neuropodia thin, smooth and without sheaths; notopodia with bundle of capillaries of different length, capillaries smooth and without sheaths, very long capillaries present in notopodia of chaetigers 29–35. Sabre chaetae as stout granulated chaetae with short elongate tip in inferior position in neuropodia, present from chaetiger 6 (in holotype as well as present material), in posterior chaetigers sabre chaetae well separated from hooks and capillaries of the neuropodium, further anteriorly positioned in immediate vicinity of capillaries or even slightly posterior to them, 2–3 sabre chaetae in each neuropodium.
Pygidium unknown.
Pigmentation. Pigment not discernable in formalin preserved specimen.
Methyl green staining pattern. No distinct staining pattern.
Ecology. Malacoceros jirkovi was found in a sample from the Great Meteor Seamount in coral and crushed shell gravel in about 300 m of depth. Material studied by Sikorski (1992) was found in sand, in sand mixed with shells, and in muddy sand in water depths ranging between 140– 235 m.
Geographical distribution. Southern Norwegian Sea, Great Meteor Seamount.
Remarks. The genus Malacoceros is one of the less speciose spionid genera (12 species according to Delgado- Blas & Díaz-Díaz 2013). The most recent publication on this genus is by Delgado-Blas & Díaz-Díaz (2013) providing the description of the new species M. longiseta from Venezuela, and also an overview about important characters of Malacoceros species. Based on the prostomial shape, the morphology of the first parapodia, and the beginning of neuropodial hooks between chaetigers 26–32, our examination of the Malacoceros from the Great Meteor Seamount led to an identification as M. jirkovi Sikorski, 1992 . Differences to the original description included, most importantly, the beginning of sabre chaetae on chaetiger 6 instead of chaetigers 23–27 as stated by Sikorski (1992), and furthermore the shape of neuropodial lamellae with a minute pointed tip in the seamount specimen which is obviously not present in specimens from the southern Norwegian Sea, the type locality of this species. However, a check of the paratype of M. jirkovi available from the ZMU collections in Copenhagen revealed that sabre chaetae are actually present from chaetiger 6, as they were in the present material and this erroneous statement by Sikorski (1992) has to be corrected for M. jirkovi . Such an early start of sabre chaetae is unusual for Malacoceros spp. and has been reported only for two other species, both from Venezuela: M. cariacoensis Delgado-Blas & Díaz-Díaz, 2010 and M. longiseta Delgado-Blas & Díaz-Díaz, 2013 , in which sabre chaetae start on chaetiger 5. In all other Malacoceros species they are present at the earliest from chaetiger 15–20 but most often from about chaetigers 25–30 ( Delgado-Blas & Díaz-Díaz 2013).
The observed differences concerning the shape of the neuropodial lamellae might be due to the smaller size of the seamount specimen; in the considerably larger paratype the neuropodial lamellae are of rounded shape. Sikorski (1992) observed the presence of tridentate neuropodial hooks in M. jirkovi . Opposed to this we can only state the presence of bidentate hooks with certainty for both the paratype and the seamount specimen. The observation of the number of apical hooks is difficult due to their small size and we cannot rule out that a third tooth is sometimes detectable. The conspicuously long capillaries in posterior notopodia observed in the seamount specimen are also present in the paratype.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Malacoceros jirkovi Sikorski, 1992
| Bick, Andreas, Guggolz, Theresa & Götting, Miriam 2014 |
Malacoceros jirkovi
| Sikorski 1992: 105 |