
Metadynomene tuamotu, Ng, Peter K. L. & Mclay, Colin L, 2010
|
publication ID |
https://doi.org/ 10.5281/zenodo.294087 |
|
DOI |
https://doi.org/10.5281/zenodo.6203107 |
|
persistent identifier |
https://treatment.plazi.org/id/039B87DC-FFC6-A54D-F991-F88950323A1A |
|
treatment provided by |
Plazi |
|
scientific name |
Metadynomene tuamotu |
| status |
sp. nov. |
Metadynomene tuamotu View in CoL sp. nov.
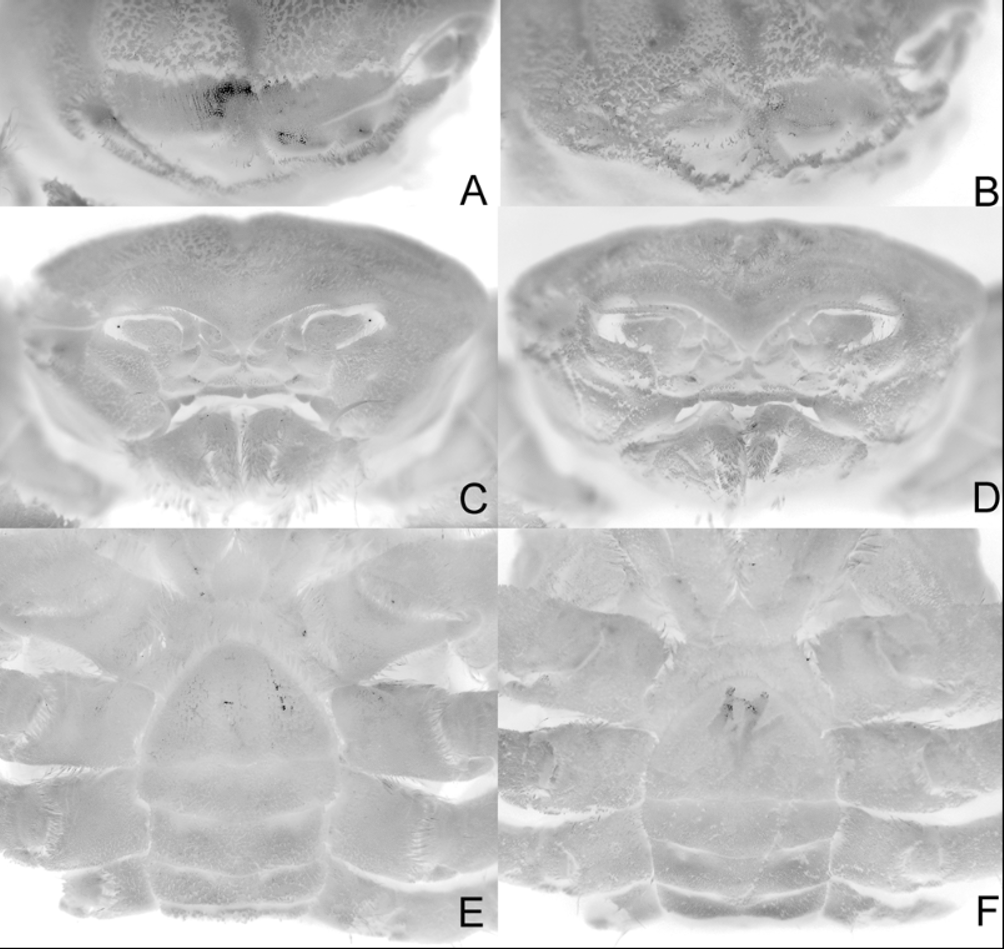
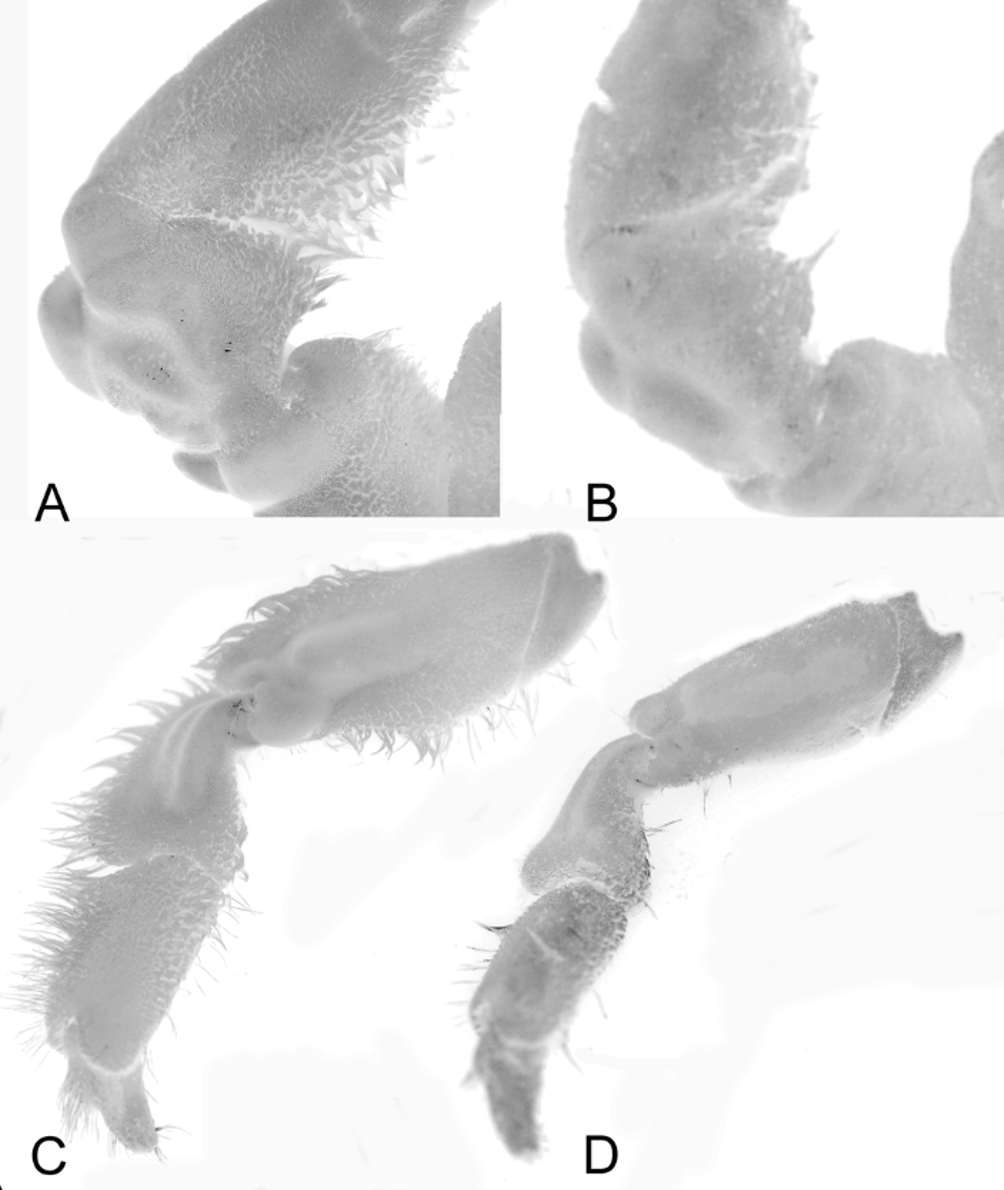
( Figs. 1A View FIGURE 1. A ; 2A, C, E; 3A, C; 4A)
Metadynomene tanensis View in CoL — McLay 1999: 521 (part) (not Dynomene tanensis Yokoya, 1933 View in CoL ).
Material examined. Holotype male (29.1 × 27.6 mm) (MNHN-B26566), Fangatafu, Tuamotu Archipelago, French Polynesia, station 487, 22°14.1'S 138°47.2'W, 310 m, coll. J. Poupin, 25 April 1995.
Diagnosis. Carapace width slightly greater than length; dorsal, lateral surfaces densely, uniformly covered with short, soft setae outlining regions, grooves on carapace, actual surface being almost completely obscured; suborbital margin shelf-like, projecting, visible from dorsal view; anterolateral margin with 4 low but distinct teeth (including external orbital tooth) before end of branchial groove; start of posterolateral margin (after end of branchial groove) with small low tubercle; cheliped carpus with prominent lobe on outer margin; cheliped dactylus gently curved with subspatuliform distal part; inferior margin of P2–P4 with less than 5 spines; merus, carpus of P2–P4 subfoliaceous with distinct cristae dorsal margin separated from rest of article by prominent longitudinal groove; male telson subtriangular.
Etymology. The species is named after its type locality, the Tuamotu Archipelago. The name is used as a noun in apposition.
Remarks. Guinot (2008: 10) noted that a suite of characters argued for the recognition of a separate subfamily, the Metadynomeninae , for Metadynomene . These include the uropod being as long as abdominal somite 6 (see also Guinot & Bouchard 1998), the presence of a small process on the coxa of P3, the basisischium of P5 not fused to the merus and the coxa of the P5 covers the penis. The present new species does not add any new characters that define the genus or sub-family.
The more ovoid carapace shape of M. tuamotu sp. nov. ( Fig. 1A View FIGURE 1. A ) is closer to M. devaneyi , with M. tanensis being more quadrate ( Fig. 1 View FIGURE 1. A B). This is because the posterolateral margin of M. tuamotu sp. nov. is relatively longer than that of M. tanensis . Significantly, the third anterolateral tooth of M. tuamotu sp. nov. ( Fig. 2A View FIGURE 2. A, C, E ) is relatively smaller than that of M. tanensis ( Fig. 1 View FIGURE 1. A B) even though the overall size of the type is larger. Other differences include the relatively deeper grooves and more prominent lobes on the outer margin of the carpus of the cheliped on M. tuamotu sp. nov. ( Fig. 3A View FIGURE 3. A, C ) (less prominent in M. tanensis , Fig. 3B View FIGURE 3. A, C ), the proportionately stouter P2-P 4 in which the dorsomarginal cristae of the merus and carpus are more developed ( Fig. 3C View FIGURE 3. A, C ) (relatively more slender with much lower dorsal cristae in M. tanensis , Fig. 3 View FIGURE 3. A, C D) as well as the relatively more triangular male telson ( Fig. 2 View FIGURE 2. A, C, E E) (more semicircular in M. tanensis , Fig. 2 View FIGURE 2. A, C, E F). Although the type male of M. tuamotu sp. nov. is larger than any known M. tanensis s. str., the suite of differences observed here suggest they are not conspecific. In addition, the kind of pubescence of the two species appears to be slightly different. In M. tuamotu sp. nov., the setae are relatively finer and the pubescence generally more velvety. In M. tanensis , the pubescence is generally of a coarser texture ( Figs. 1A View FIGURE 1. A , 2A, C View FIGURE 2. A, C, E versus Figs. 1 View FIGURE 1. A B, 2B, D) (see Ahyong et al. 2008: 72, Figs. 43, 44). The live colours of the two species also appear to differ. Metadynomene tuamotu is generally yellowish-orange overall ( Fig. 4 View FIGURE 4 A) while M. tanensis is brown to orange-brown ( Fig. 4 View FIGURE 4 B, C). The male first gonopod of M. tuamotu sp. nov. is not diagnostically different from M. tanensis .
The four known species of Metadynomene have non-overlapping distributions: M. crosnieri McLay, 1999 , is known only from the type locality, the Glorieuse Islands, south of the Seychelles, Indian Ocean; M. tanensis ( Yokoya, 1933) is widespread in the Western Pacific, from Japan to New Zealand ( McLay 2009); M. devaneyi (Takeda, 1977) is known from the Hawaiian and Marquesas Is., French Polynesia ( McLay 1999); and M. tuamotu sp. nov. is only known from the Tuamotu Archipelago in French Polynesia, thus far. The two species from French Polynesia are recorded from island groups separated by approximately11°, the Marquesas and Tuamotus. The taxonomy of the most widespread species, M. tanensis , will also need further attention. The specimens on hand ( McLay & Ng 2005; Ahyong et al. 2009; McLay 2009) show some differences in carapace form and colour, and we have doubts if these can all be explained by variation.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Metadynomene tuamotu
| Ng, Peter K. L. & Mclay, Colin L 2010 |
Metadynomene tanensis
| McLay 1999: 521 |