
Naupactus dissimulator Boheman, 1840
|
publication ID |
https://doi.org/ 10.1080/00222933.2017.1346715 |
|
publication LSID |
lsid:zoobank.org:pub:051587DD-37C2-4216-AA61-0E563BB44D64 |
|
persistent identifier |
https://treatment.plazi.org/id/3C7887C4-756E-CE00-539A-FF30AE3AB3EC |
|
treatment provided by |
Felipe |
|
scientific name |
Naupactus dissimulator Boheman |
| status |
|
Naupactus dissimulator Boheman View in CoL
( Figures 2 View Figure 2 (a, b), 3i, 4(i, j), 5c)
Naupactus dissimulator Boheman 1840, p. 8 View in CoL . Type material: male labelled as typus, Buenos Aires, at NHRS, herein designated as lectotype . Body length: 10 mm.
Naupactus fallax Boheman 1840, p. 9 View in CoL . Type material: female labelled as allotype (error), Brazil, at NHRS, herein designated as lectotype . Synonymy by Kuschel in Wibmer and O ’ Brien 1986, p . 59. Body length: 11.5 mm .
Naupactus dissimulator: von Dalla Torre et al. 1936, p. 19 View in CoL (catalogue); Blackwelder 1947, p. 794 (checklist); Hustache 1947, p. 39 (in key); Wibmer and O ’ Brien 1986, p. 59 (checklist).
Redescription. Female ( Figure 2a View Figure 2 )
Body length 10 – 12.5 mm. Vestiture brown, with pattern of white stripes on pronotum and elytra, and elytral disc usually rubbed; pronotum with distinct white stripe along midline, extended along whole elytral suture; middle half of 3° interval with white maculae (female); 7° interval with white stripe on anterior third, obliquely curved towards 5° interval in posterior half. Rostrum about as long as wide at apex; lateral carinae very strong, denuded, subparallel. Eyes round, convex. Scape reaching anterior margin of pronotum; funicular article 2, 1.5 – 1.9× as long as article 1. Pronotum subcylindrical, 1.20 – 1.25× as wide as long; disc flat, rugose. Elytra oval, 1.60 – 1.70× as long as wide; disc flat and usually rubbed; sides moderately curved and narrowed towards apex; apical tubercles large; intervals about 3× as wide as punctures of striae. Front femora 2.60 – 2.70× as long as wide and 1.40 – 1.45× as wide as hind femora. Front tibiae with large mucro and about 10 large denticles on inner margin; middle tibiae with smaller mucro and lacking denticles; hind tibiae lacking mucro and denticles; corbel of hind tibiae lacking. Ventrite 5 slightly shorter than ventrite 2.
Female genitalia
Ovipositor about ¾ as long as abdomen, with rows of 3 – 5 setae on posterior third, on external sides of baculi (see Figure 3d View Figure 3 ). Spermatheca ( Figure 3i View Figure 3 ) subcylindrical, large (0.7 mm); nodulus tubular, long, and ramus incipient; spermathecal duct 2 – 3× as long as spermatheca.
Male ( Figure 2b View Figure 2 )
Body length 8 – 11 mm. Rostrum 1.15 – 1.20× as long as wide. Pronotum about as wide as long; disc slightly convex. Elytra 1.80 – 190× as long as wide; maximum width similar to maximum width of pronotum; apical tubercles very small, almost indistinct. Ventrite 5 about 1.20× as long as ventrite 2, apex rounded.
Male genitalia ( Figure 4i, j View Figure 4 )
Body of penis about twice as long as penis apodemes; apex acute, with waved sides, but not arrow-pointed; ostium about 4 – 5× as long as wide; endophallus with spiny area and large, strongly sclerotized internal armature, consisted on a pyriform central piece and two lateral wing-shaped pieces.
Other material examined
ARGENTINA. Buenos Aires: Delta, 1945, M Viana (1f MLP); Delta Paraná, Río Luján, Monrós (2f FIML); Delta, 6 April 1927, on orchids, Kisliuk (1f USNM); Delta, Carapachay, October 1940, Monrós (1m BMNH); Delta, Paraná Guazú , December 1952 (3f 4m BMNH); Dique Luján, November 1938, Daguerre (3f 2m USNM); Isla Martín García, June 1936, M Viana (2f 3m MLP); idem, January 1938, M Viana (2f 1m MLP, 1f MNHN); idem, April 1938, M Viana (1m MLP); Isla Paulino, 22 December 1977, Cicchino (3f 2m MLP); idem, 18 December 2009, G del Río (1f MLP, IBOL); Isla Santiago, 21 April 1945, Olivier (1m MLP); idem, 18 January 1919 (1f MNHN); La Plata, (2f 3m MLP); idem, AR Bezzi (2f 2m MLP); idem, 1939, Costa (1m MLP); idem, C Bruch (1m MLP); Palermo, 20 December 1915, J Bosq (1m MLP); idem, 18 March 1922, Marshall (1f USNM); Punta Lara, December 1931, Bosq (1f 3m MLP); idem, 15 March 1958, Aguirre (4f USNM); idem, January 1958, Aguirre (5f 1m USNM); idem, December 1931, J Bosq (4m MLP); idem, 30 December 1969 (1f BMNH); San Fernando, 25 November 1932 (1f 2m MLP); idem, December 1939 (1m MLP); idem, 8 January 1950, JB Daguerre (1f 1m USNM); San Ignacio, Villa Lutecia, 1910, ER Wagner (1f 3m MNHN); San Isidro, 26 March 1927, on peach, Kisliuk (1f USNM); idem, 3 January 1932, J Bosq (2f MLP); idem, 6 January 1932, J Bosq (1f 3m MLP); March 1932 (2f 2m MLP); Tigre, 1935, M Viana (1f 1m MLP); idem, November 1958, Daguerre (2f 1m USNM); no loc. 20 March 1906, C Bruch (4m MLP); no loc., Richter (5f 3m MLP); no loc. J Bosq (4f MLP). Corrientes: Corrientes city, July 1934, J Bosq (1m MLP). Entre Ríos: Brazo Largo, May 1940, Castillo 1f MLP); Gualeguaychú, JA Haedo Rossi (1f FIML); Villa Paranacito, July 1939, Castillo (1m MLP). Misiones: NW Apóstoles, Rt 10, 27 52.451 S, 55 39.514 W, 3 March 2010, 146 m, G del Río (1f MLP); Campo Viera, 1952 (1f BMNH); Colonia Taranco, Dto. Candelaria, 20 November 1948, Viana (6f MLP); Iguazú, November 1944, M Birabén (2f MLP); Iguazú, 13 March 1945, Hayward-Willink (2f 2m FIML); Loreto, September 1955, J Bosq (1f MLP); Parque Provincial Las Araucarias, 26° 37 ʹ 52 ʹʹ S, 54° 06 ʹ 13 ʹʹ W, 546 m, 6 March 2010, G del Río (1f 2m MLP-IBOL); Parque Provincial Moconá, 27° 07 ʹ 37.4 ʹʹ S, 53° 56 ʹ 55.5 ʹʹ W, 1 April 2012, 365 m, P Dellapé & S Montemayor (3f 1m MLP- IBOL); Puerto Bemberg, 30 March 1945, Hayward Willink & Golbach (2m FIML); Santa Ana, August 1952, 160m (1f 1m BMNH); San Ignacio, Villa Lutecia, ER Wagner, 1910 (1f 3m MNHN); Santa María, December 1947 (1f MLP); no loc. C Berg (1m MLP). Santa Fe: Rosario, September 1961, A. Martínez col (1f 4m MZSP). BOLIVIA. Cochabamba: Yungas del Palmar, A. Martínez (1f MZSP). BRAZIL. Paraná: Bocaiuva , December 1963 (1f 1m MZSP); Curitiba, December 1937 (4f MZSP); idem, December 1939, Guerin (1f MNRJ); idem, March 1938 (5f USNM); idem, 7 October 1936 (1f BMNH); idem, 13 January 1969, CW & L O ’ Brien, Araucaria forest (5f CWOB); near Curitiba, 9 January 1963, Blake & Cochran (7f USNM); 3km Foz do Iguazú, 9 October 1968, CW & L O ’ Brien (2f CWOB). Rio Grande do Sul: Canela, 2 March 1976, Lise (1f USNM); Farroupilha, November 1960 (1f USNM); São Francisco de Paula, January 1955, Buckup (1f USNM); Tainhas, April 1959, F Plaumann (1m MZSP); Tainmezinho, PN dos Aparados da Serra, February 1960 (2f USNM); Torres, 21 November 1976, Lise (1f USNM). Santa Catarina : Bom Jesus, 28° 40 ’ S, 50° 12 ʹ W, 1200m, December 1962, F Plaumann (1f 1m MZSP); Florianópolis, Lagoa Pequena, Pós Duna, Ocotea punchella (Lauraceae) , 26 May 2007, F F Albertoni (1f 1m MZSP); Joinville, September 1942, B Pohl (3f 1m MZSP); Nova Teutonia, April 1966, F Plaumann (1m MZSP); Nova Teutonia, 27° 11 ʹ S 52° 23 ʹ W, 300 – 500 m, April 1954, F Plaumann (1m MZSP); São Bento do Sul, 26° S 50° W, 800m, September 1923, A Maller (1f BMNH). PARAGUAY. Itapúa: Hohenau (1f MLP, 5f 3m MNHN). URUGUAY. Colonia: Barra Arroyo El Chileno, 11 January 1970, GJ Wibmer (4f 1m MLP); camino a Salto (2f USNM); Colonia, 7 December 1942 (1f USNM); Playa Arenisca, 6 January 1979, GJ Wibmer (1f MLP, 1f CWOB). Maldonado: Cerro Negro, 27° 51 ʹ S, 50° 48 ʹ W, 900 m, December 1962, F Plaumann (1f MZSP). Montevideo: Montevideo, Palermo, 18 March 1922, Marshall (1f USNM); Sayago, 8 January 1969, MS Moratorio & GJ Wibmer (1f MLP, 2f CWOB). Paysandú: Paysandú, Puerto Pepe Ají, 21 – 25 January 1970, MA Monné, G Wibmer & CE Cassini (1f CWOB).
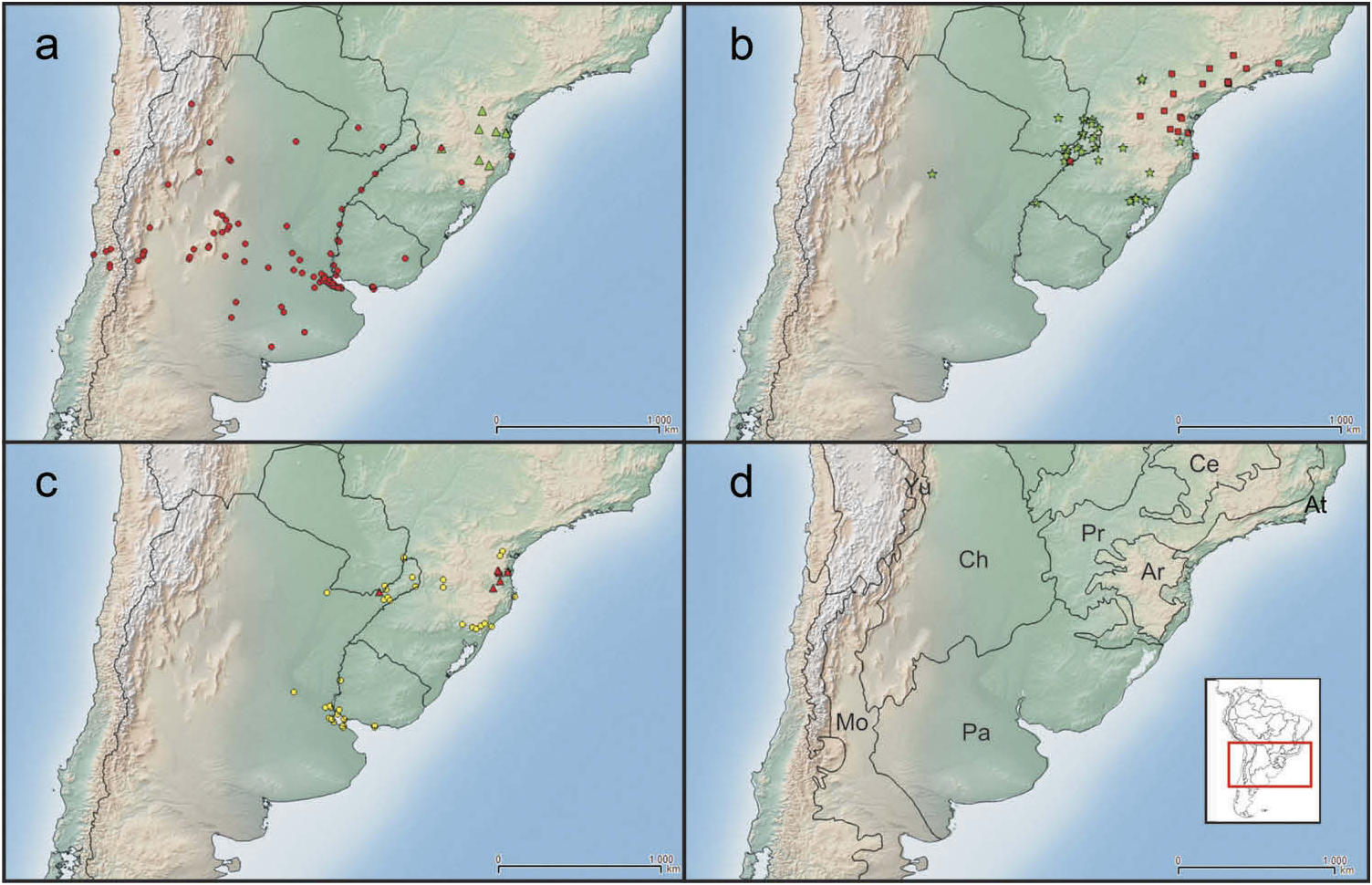
Geographic distribution ( Figure 5c, d View Figure 5 )
Naupactus dissimulator occurs mainly in the southern Paraná forest and extends along the gallery forests of Paraná River , down to the banks of La Plata River. It has been recorded for Argentina (Buenos Aires, Corrientes, Misiones and Santa Fe), Bolivia (Cochabamba), southern Brazil (Paraná, Santa Catarina and Río Grande do Sul), Paraguay (Itapúa) and Uruguay (Colonia, Maldonado, Montevideo and Paysandú). Bolivia is a new country record. This finding indicates that N. dissimulator also inhabits the Yungas forest.
Host plants
One of the native hosts of N. dissimulator is Ocotea punchella (Nees & Mart.) Mez (Lauraceae) , a semideciduous tree distributed in Argentina, Paraguay, central-eastern and southern Brazil, throughout the Atlantic and the Paraná forests. This weevil causes damage on Citrus sp. (Rutaceae) , Ilex paraguariensis Saint Hill (Aquifoliaceae) and Prunus persica (L.) Batsch ( Rosaceae ), in north-eastern Argentina ( Bosq 1943; Lanteri et al. 2002a). Eggs and oviposition habits of this species have been studied by Marvaldi (1999).
Remarks
Boheman (1840) did not specify the number of specimens of the type series of N. dissimulator , consequently we designate the only specimen at the NHRS labelled as typus as the lectotype. In the case of N. fallax , the author described male and female. In the collection of the NHRS we found only the female (one of the syntypes), that was erroneously labelled as allotype. In this paper we designate this female as lectotype.
The populations from Argentina and Uruguay are equal to the typical N . dissimulator (type material from Buenos Aires, Argentina) and slightly different from those occurring in southern Brazil, Bolivia and Paraguay . The latter are usually smaller, the pronotum and elytra are slightly narrower, and the disc of the elytra are less flat and not rubbed . In the type female of N . fallax (from Brazil), the apical tubercles are more reduced than in the females of N. dissimulator from Argentina and Uruguay.
A previous cladistic analysis of a set of Naupactus and Pantomorus Schoenherr from South America ( Scataglini et al. 2005) suggested that N. dissimulator is the sister species of N. cervinus ; however, based on the new material studied herein it might be possible that the closest species is N. marvaldiae . Naupactus cervinus is a parthenogenetic species also harmful for citrus and other fruit trees ( Lanteri et al. 2002b; Rodriguero et al. 2016), easily distinguished from N. dissimulator because it is smaller, without tubercles at the apex of the elytra, the eyes are more convex, the pronotum is smooth and the front femora are slightly wider than the hind femora. The ovipositor and spermatheca are similar in both species, but in N. cervinus the spermathecal duct is longer and very wide.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Naupactus dissimulator Boheman
| Lanteri, Analia A. & del Río, María G. 2017 |
Naupactus dissimulator:
| Blackwelder RE 1947: 794 |
| Hustache A 1947: 39 |
| von Dalla Torre KW & van Emden M & van Emden FI 1936: 19 |
Naupactus dissimulator
| Boheman CJ 1840: 8 |
Naupactus fallax
| Boheman CJ 1840: 9 |