
Neonesidea longisetosa ( Brady, 1902 )
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4903.4.1 |
|
publication LSID |
lsid:zoobank.org:pub:D8AA9035-EB27-4F50-9246-B5450D71F3E2 |
|
DOI |
https://doi.org/10.5281/zenodo.4434581 |
|
persistent identifier |
https://treatment.plazi.org/id/03F88789-847E-FFFB-FF0C-2F3AAE639AE1 |
|
treatment provided by |
Plazi |
|
scientific name |
Neonesidea longisetosa ( Brady, 1902 ) |
| status |
|
Neonesidea longisetosa ( Brady, 1902) View in CoL
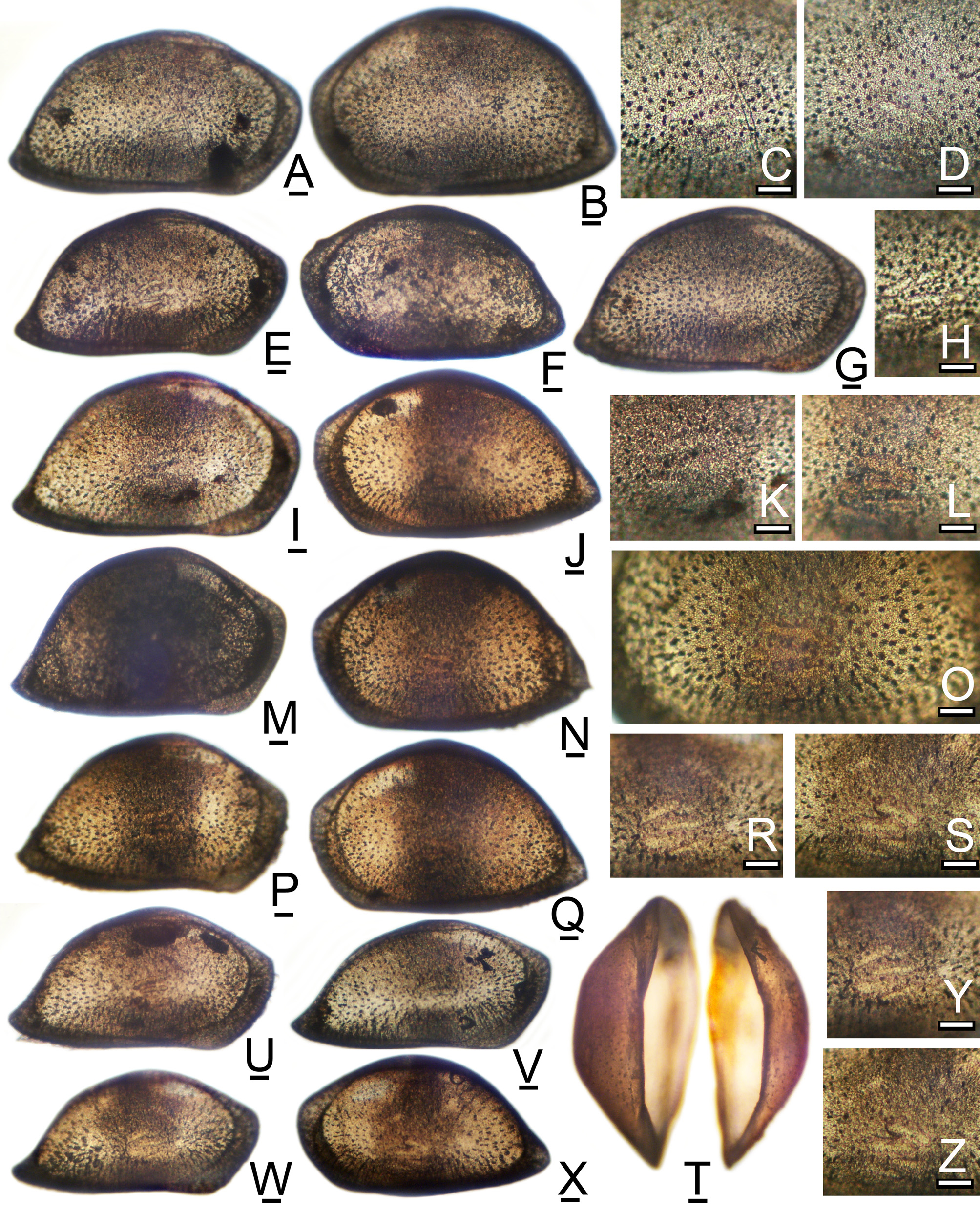
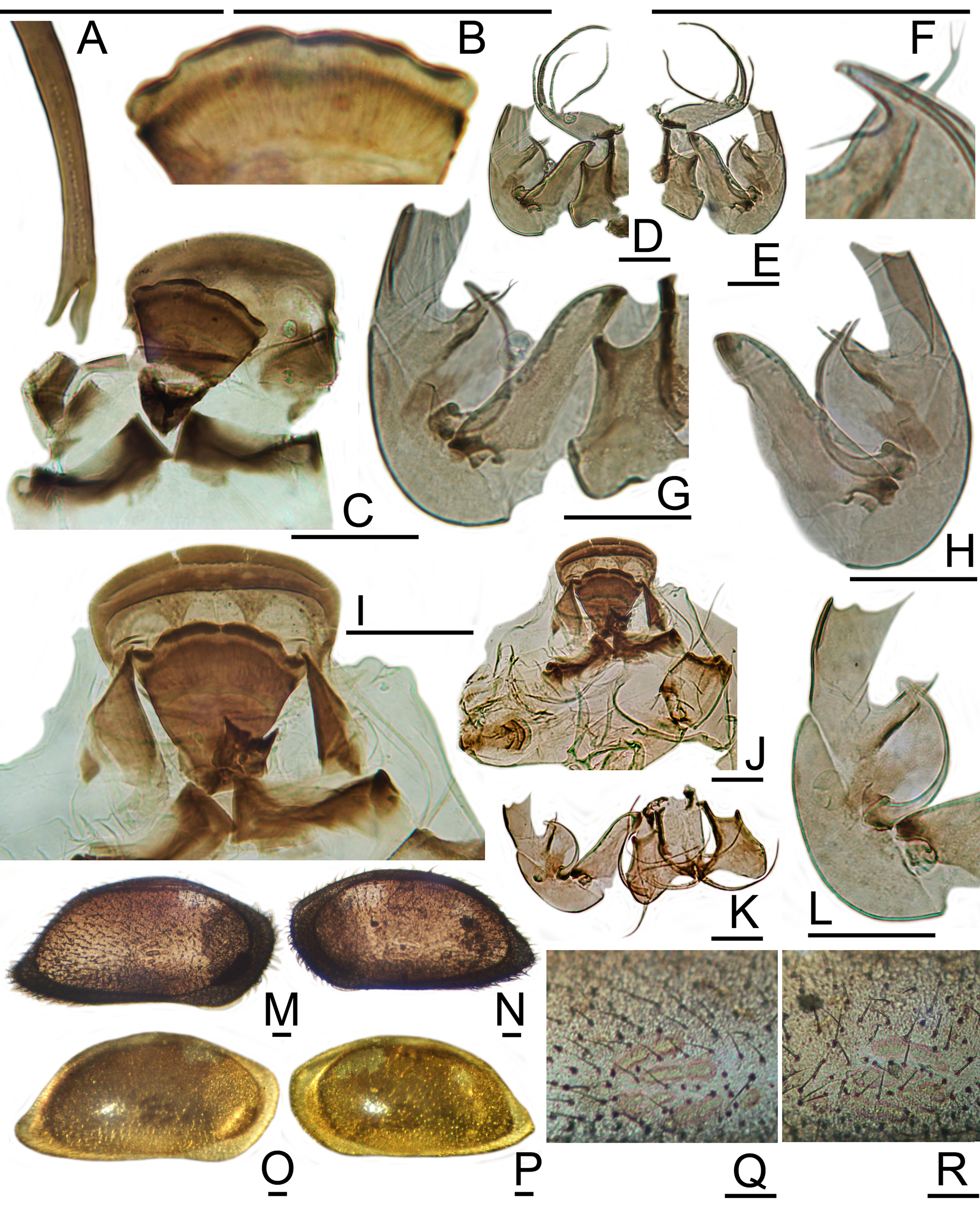
( Figure 11 View FIGURE 11 -I–P, 12A–S; Graph 4)
1902 Bairdia longisetosa sp. nov.: Brady: 197, Pl. 25, figs. 8–9.
1966 Bairdia longisetosa Brady—Bold , p. 45, Pl. 2, figs. 7a–b.
1968 Bairdia longisetosa Brady—Bold , p. 48, Pl. 2, figs. 3a–b.
partim 1969 Neonesidea gerda (Benson & Coleman), 1964 —Maddocks, p. 24, Figs. 7 View FIGURE 7 A–K.
1975 Bairdia longisetosa Brady—Bold pp. 585, 586, 589, 590, 593.
1975 Neonesidea longisetosa (Brady), 1902 —Teeter, p. 417, Figs. 3a View FIGURE 3 , 4a View FIGURE 4 .
1978 Bairdia longisetosa Brady—Bold , Table 9.
1982 Neonesidea longisetosa (Brady) —Breman, Pl. 3, fig. E.
non 1982 Neonesidea longisetosa (Brady) —Krutak, Pl. 5, figs. 1–5 [= N. gerda ].
? 1983 Neonesidea longisetosa (Brady) —Palacios-Fest et al., Pl. 1, fig. 16 (only, not fig. 17).
1988 Bairdia longisetosa Brady—Bold , p. 22.
non 1990 Bairdia cf. B. longisetosa Brady—Machain-Castillo Table 2, p. 122 [= N. gerda ].
? 1992 Bairdia longisetosa Brady—Machain-Castillo & Gío-Argáez (1992), Appendix I, p. 22 [part, may include N. gerda ]
partim 1993 Neonesidea longisetosa (Brady) —Machain Castillo & Gío-Argáez, p. 258 [part, also includes N. gerda ].
1994 Neonesidea longisetosa ( Brady, 1902) —Krutak & Gío-Argáez, p. 200, Pl. 2, figs. 1–6.
1999 Neonesidea longisetosa ( Brady, 1902) View in CoL —Coimbra et al., Pl. 3, fig. 4.
non 2000 Neonesidea longisetosa ( Brady 1902) View in CoL —Keyser & Scĥning, Pl. 2, fig. 19 [= N. omnivaga View in CoL ]
2002 Neonesidea longisetosa ( Brady, 1902) View in CoL —Coimbra & Carreño, p. 193, Pl. 2, figs. 21–22.
partim 2009 Neonesidea longisetosa (Brady) View in CoL —Maddocks et al., p. 888 [part, also includes N. gerda View in CoL ].
non 2014a Neonesidea longisetosa ( Brady, 1902) —Meireles et al., p. 16, Pl. 1, fig. a; Pl. 2, figs. a–l [= a species of the N. tenera species-group].
non 2014b Neonesidea longisetosa ( Brady, 1902) —Meireles et al., Figs. 2–9 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 View FIGURE 8 View FIGURE 9 .
Material: Three dry male carapaces with fragmentary soft parts from Belize; one fragmentary living male from the Bahamas; numerous empty carapaces and valves from shallow-water carbonate sediments of the Bahamas, Belize, Cuba, the Florida Keys, Grand Cayman Island, Jamaica, and the U.S. Virgin Islands.
Dimensions: Specimen 4045M, LVL 1.061 mm, LVH 0.614 mm. Brady reported L 1.1 mm. See also Graph 4.
Supplemental Description: Most of the characteristics described by Brady are typical for most species of the N. schulzi species-group, including the subtly subrhomboidal but streamlined carapace with equant valves, obliquely subtruncate anteroventral margin, caudate posterior angle, and conspicuous carapace sensilla. The lateral outline is elongate-ovate, with smoothly arched dorsal margin, indistinct posterodorsal angle, caudate posterior end, tripartite ventral margin with upswung anteroventral and posteroventral segments, and rounded anterior margin. The acute, caudate posterior angle of the LV is located at approximately 33% of height. Well-conserved specimens have a tiny terminal spine at the caudal angle of each valve, conspicuous anteromarginal and posteromarginal denticles on the LV, and narrow anteromarginal and posteromarginal frills on the RV. The patch pattern consists of a large central oval spot, small spots in the anterior and posterior angles, and less distinct spots or streaks at the anterodorsal and posterodorsal angles. The dorsal spots are most easily seen in dorsal views of a whole carapace.
The hemipenis of N. longisetosa has a moderately long, arcuate copulatory tube, which arises near the base of the median segment. Distally, the tube is contained within a hollow, obliquely-curved, well-sclerotized, claw-like projection. At its end the copulatory tube splits into two or three threads. The terminal segment of the hemipenis is a hood-like plate, the inner surface of which is densely corrugated with wavy chitin threads, which rise above the surface as setules or tiny thorns. At the base of this segment is a well-sclerotized, obliquely-wrinkled ridge with one sharp tooth, the end of which articulates against the base of the clawlike projection.
The esophageal plate is a rather broadly flaring triangle. The curved posterior edge has a large, sharply triangular to subpyramidal tooth on either side of the medial gap, followed by about four smaller teeth of equal or diminishing sizes, subconical to subpyramidal in shape, and a prominent cluster of multilobate teeth at the corner. The U- to V-shaped ventral bracket has long diagonal horns directed outward and down, triangular lateral walls, and a row of tubercles along the anterior edge.
Remarks: N. longisetosa is significantly larger than N. gerda but has similar carapace proportions. The lateral outline usually has a more up-curved posterior angle and more conspicuous marginal denticles on the LV. In dorsal view, as compared to N. gerda , it has more curved contours with more rounded anterior and posterior ends. In fresh specimens the central opaque spot of the patch pattern is larger than that of N. gerda and approximately oval rather than oblong. The single posterodorsal opaque spot easily distinguishes N. longisetosa from N. gerda , which has no spot at this location. This mark is also convenient for quickly separating adult and juvenile valves of N. longisetosa from Bairdoppilata cushmani ( Tressler, 1949) , which consistently has two opaque spots at this posterodorsal location, and which occurs abundantly in the same subfossil assemblages.
The setulose, quarter-spherical hood and asymmetrical serrate ridge of the distal segment of the hemipenis of N. longisetosa are easily distinguished from the smooth, half-cylindrical lamellae and aesthetasc of N. gerda .
The three specimens illustrated as N. gerda by Maddocks (1969, Figs. 7 View FIGURE 7 A–K) from Tortugas (Florida Keys) and Bimini and Andros (Bahama Islands) all belong to N. longisetosa . Her drawing of the hemipenis ( Fig. 7D View FIGURE 7 ) shows the setulose distal texture characteristic of that species. Her drawings of the valve exteriors ( Figs. 7 View FIGURE 7 H–K) show the distinct posterodorsal opaque spot, which is characteristic of N. longisetosa . The measurements cited by Maddocks (1969, p. 24) for these three males are among the smallest known for the species.
The specimens illustrated as N. longisetosa by Krutak (1982) from reefs off Vera Cruz are smaller, more angular in outline, and more triangular in shape. It is likely that they are N. gerda .
The specimen illustrated as N. longisetosa from Bermuda by Keyser & Scĥning (2000) is N. omnivaga . N. longisetosa has not been seen in sediment samples from the Bermuda Platform.
N. longisetosa resembles N. caraionae n. sp. in carapace form and outline, but it is smaller. The hemipenis has a much shorter copulatory tube and a hood-like, textured terminal segment, easily distinguished from the extraordinarily long tube and discoidal terminal segment of N. caraionae .
N. longisetosa has sometimes been confused with Neonesidea crosskeiana ( Brady, 1866) , which lives in the eastern Mediterranean; see Titterton et al. (2001) for redescription and clarification of that species.
The population from the Azores, which was misidentified as N. longisetosa by Meireles et al. (2014a, 2014b), represents a species of the N. tenera species-group (informal). Their illustrations of the ovate lateral outline show a nearly symmetrical, high-arched dorsum and a much less caudate posterior end. It shows similarities to Mediterranean species, such as N. decipiens or N. mediterranea , well as to the West African species, “ Paranesidea ” multiforma Witte, 1993. Meireles et al. (2014a, p. 16) attempted to synonymize N. gerda and N. omnivaga with N. longisetosa , but their justification was vague (“The collected specimens are identical to N. longisetosa , considering the carapace morphology. The morphology of the soft parts is comparable to the figures that Maddocks (1969) gave for Neonesidea gerda ….”). Although they recovered hundreds of living and subfossil specimens, Meireles et al. did not provide carapace dimensions, did not describe or illustrate soft parts, and did not say whether they examined actual specimens from Bermuda and Caribbean localities. The species living in the Azores is certainly not conspecific with the material examined here for N. caraionae , N. gerda , N. longisetosa , and N. omnivaga . Indeed, the wave-dominated rocky slopes of the Azores, washed by the cold Canaries Current (14 oC), are unlikely to share species in common with the carbonate platform assemblages of the tropical Caribbean and Bermuda.
Distribution: N. longisetosa was originally described by Brady from St. Thomas, U.S. Virgin Islands. In the samples examined for this study, it is abundant in shallow-water carbonate sands of Belize, Jamaica and Grand Cayman Island; and it is less common or rare in samples from the Bahamas, the Florida Keys, and Hopetown Harbour, U.S. Virgin Islands. It is not represented in collections from Bermuda.
N. longisetosa has been reported from Alacran Reef on the northwestern Yucatan platform, northeastern Yucatan, and the Caribbean coast of Mexico; Belize; Buccoo Reef, Tobago; Nicaragua; Colon Harbour, Panama; the Gulf of Venezuela; the Brazilian equatorial shelf; and the Neogene of Jamaica and the Dominican Republic ( Breman 1982; Bold 1966, 1968, 1989; Teeter 1975; Palacios-Fest et al. 1983, Krutak & Gío-Argáez 1994, Coimbra & Carreño 2002). On Alacran Reef it was said to be consistently restricted to a particular facies: “Shallow, agitated environment, e.g., windward reef flat, shallow patchreefs, leeward sandflats” ( Bold 1989, p. 145, Table 1). In northeastern Yucatan it was described as abundant in modern carbonate grainstones, but less common in lagoonal sediments ( Krutak & Gío-Argáez 1994). Reported occurrences on the western platform of Yucatan and at localities in the central and southern Caribbean are plausible but cannot be verified without illustrations and dimensions ( Machain-Castillo & Gío-Argáez 1992 Appendix I; Bold 1977, Table 3; 1988, p. 22). The identifications under this name from the Veracruz-Anton Lizardo reefs by Krutak (1982) apply to N. gerda . Reported occurrences in the Bay of Campeche ( Machain-Castillo 1990) probably apply to N. gerda . In the checklist by Maddocks et al. (2009, p. 888), the distribution of this species should be restricted to exclude occurrences in the northwestern and southwestern Gulf of Mexico, which apply to N. gerda . Fossil specimens have been reported from the Upper Miocene to Recent of Venezuela, Panama, the Dominican Republic, Jamaica, and other Caribbean localities ( Bold, 1966, 1968, 1975, 1978, 1988).







No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.