
Oioceros rothii ( Wagner, 1857 )
|
publication ID |
https://doi.org/ 10.5281/zenodo.4650685 |
|
persistent identifier |
https://treatment.plazi.org/id/8938AB31-FFFE-FFC7-FF2A-3C8DF0CD3387 |
|
treatment provided by |
Felipe |
|
scientific name |
Oioceros rothii ( Wagner, 1857 ) |
| status |
|
Oioceros rothii ( Wagner, 1857)
Synonymy: see Pilgrim & Hopwood (1928: 24).
HOLOTYPE. — Frontlet ( Wagner 1857: pl. 8, fig. 20), Palaeontological Museum, Munich.
MATERIAL EXAMINED. — AMPG Pikermi collection: almost complete skull with the left and right horncores (partly preserved), the left P3-M3, and the right
P2-P3 and M1-M3 ( PG 95/1502a); right maxilla fragment with the P2-M2 (PA 3509/91); right mandibular ramus with the p3-m3 (PA 3519/91); right mandibular ramus with the p4-m3 ( PG 95/1502b); left mandibular ramus with the p4-m3 ( PG 95/1502c); frontlets with the left and right horn-cores ( PG 95/1509, PG 95/1510, PG 95/1514, PA 1412/91); right horn-cores (PA 909/91, PA 3553/91, PG 88/1583); left horn-cores (PA 2990/91, PG 88/1540). The mandibles PG 95/1502b & c were found connected to the skull PG 95/1502a by matrix. The frontlet PA 1412/91 brings also the code PA 3031/91.
MNHN Pikermi collection: frontlets ( PIK. 2244, PIK. 2246, PIK. 2247, PIK. 2249). PIK. 2249 has been figured by Gaudry (1865: pl. 52, figs 2, 3).
NHML Pikermi collection: frontlet with the left and right horn-cores ( M 11461 View Materials ). This specimen is mount- ed on a plaster base and according to Pilgrim & Hopwood (1928: 25) it is not certain that the two horn-cores were derived from the same individual.
LOCALITY. — Pikermi, Greece.
AGE. — Late Miocene, middle Turolian (MN 12).
DIAGNOSIS. — Medium sized Oioceros ; lyrate horncores with elliptical cross section throughout their length; postero-lateral keel that runs almost the whole horn-core; horn-core torsion of one gyre; interfrontal and parietofrontal sutures open and complicated; interfrontal suture only slightly elevated between the horn-core bases; ethmoid fissure present; occipital face in obtuse angle to the braincase roof; basioccipital slightly wider posteriorly than anteriorly, without median groove; premolar series long relatively to the molars (LP2-P4/LM1-M3) × 100 = c. 78.
DIFFERENTIAL DIAGNOSIS. — O. rothii differs from O. atropatenes by the larger general size; the larger dimensions of the horn-cores, their more important torsion (less than one gyre in O. atropatenes ), and their more elliptical cross section at their bases; the larger dentition, the more advanced degree of molarization on the lower premolars, and the larger premolar series relative to the molars.
DESCRIPTION
Skull
The available skull (PG 95/1502a) is almost completely preserved ( Fig. 1 View FIG ). The face is short (M3 at the level of the anterior third of the orbit) and slightly bent relative to the braincase. Because of the preservation status it is not clear whether there is a lachrymal fossa, but there is an ethmoid fissure. The premaxillary bones are in contact with the nasals. The width of the
A
premaxilla is 8 mm and its contact with the nasal measures about 20 mm. The posterior end of the premaxillary bone is above the P3. The nasals widen posteriorly, their maximum width occurs at about 20 mm before their posterior end which is above the anterior margin of the orbits. Most probably there is no contact between the nasal and the lachrymal. The infraorbital foramen
Oioceros rothii ( Mammalia, Artiodactyla , Bovidae ) from Pikermi ( Greece)
opens above the anterior half of P3. The maxillolachrymal suture is not clearly visible. The most anterior point of the maxillozygomatic suture is above the posterior part of the M1, and there is a facial crest that extends anteriorly to the M1. The frontals are depressed between the nasals and the horn-core bases.
The interfrontal and parietofrontal sutures are open and complicated. The former is slightly elevated between the horn-cores. The parietofrontal suture has a slight forward central indentation, forming a large obtuse angle in dorsal view. The frontals between the horn-cores are almost at the same level as the supraorbital margins, and their thickness behind the horn-cores is small (about 5 mm). The supraorbital foramina are small, and open above the centre of the orbits almost level with the supraorbital margins, very close to the bases of the horn-cores. The maximum distance between the supraorbital foramina ranges from 32.3 mm to 40.2 mm (AMPG and MNHN, Pikermi, n = 7). The orbital margins are not completely preserved but the orbits look large in relation to skull size. The maximum width over the horn-core bases ranges from 58.4 to 68.8 mm (AMPG and MNHN, Pikermi, n = 8). The supraorbital margins project laterally. In PA 1412/91 for example, the greatest width at the supraobital margins is estimated to at least 84 mm, while the width at the bases of the horncores and laterally is only 65.4 mm. The anterior margin of the orbits lies above the M3/M2 contact. The maxillopalatine suture cannot be traced clearly, its likely anterior limit, however, is level with the M1/M2 contact.
The braincase is relatively long, almost as long as the preorbital region ( Fig. 1 View FIG ), its greatest width is at its mid-length, and there are no temporal lines. The width of the skull behind the horn-cores (at the parietofrontal suture) ranges from 53.1 to 58.1 mm (AMPG, Pikermi, n = 5). The top of the braincase, close to the centre of the parietal bone, is slightly swollen. The supraoccipital bone has the form of a transversely elongated trapezium. The occipital face forms an obtuse angle with the roof of the braincase ( Fig. 1 View FIG ). The nuchal crest is not prominent. The external occipital crest is slightly developed and the two surfaces laterally of this crest form a wide obtuse angle. The foramen magnum is slightly higher (16.8 mm) than broad (14.6 mm). The mastoid bone is not in contact with the parietal. The angle formed by the level of the nasals and the line that connects the nuchal crest with the meeting point of the interfrontal and parietofrontal sutures is about 127°. The angle formed between the previously mentioned meeting point, the nuchal crest and the dorsal margin of the foramen magnum is about 118° (parameters measured on a sagittal profile of the skull).
The auditory bulla is laterally compressed, strongly connected to the jugular process and the basioccipital, and its occipital face lies level with the posterior limit of the posterior tuberosities of
A
B
the basioccipital ( Fig. 2 View FIG ). The external auditory meatus directs postero-dorsally ( Fig. 1 View FIG ). The jugular processes are broken ventrally and the degree of their downward extension cannot be estimated. The post-tympanic process directs ventrally and anteriorly, passing under the external auditory meatus ( Fig. 1 View FIG ). The basioccipital is rectangular, its transverse section at its central area is convex and the posterior tuberosities are slightly wider than that the anterior ones (Annexe: Table 1). The latter are anteroposteriorly elongated and extend anteriorly beyond the level of the oval foramina, which are small (larger diameter about 5 mm).
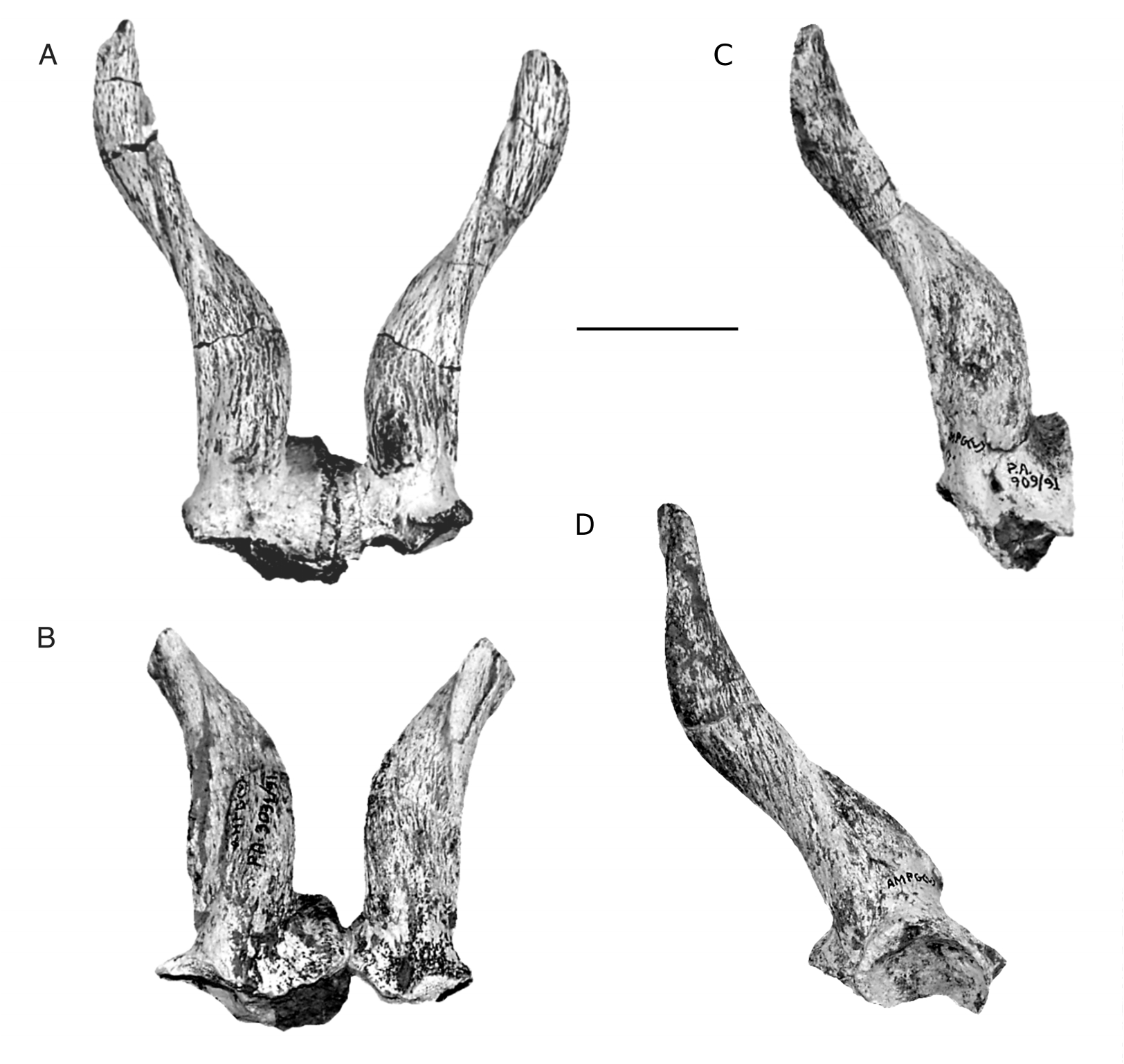
Horn-cores
None of the available horn-cores is completely preserved. The most completely preserved horncores are the right one of PG 95/1514, and PA 909/91. The left horn-core of PG 95/1502a shows a pathological possibly thickening at the base of its postero-lateral keel. The horn-cores of O. rothii , characteristic for their inverse torsion (counter-clockwise in the right horn-core from the base upwards when observed in dorsal view), are inserted above the orbits, so that, in lateral view and with the tooth row horizontal, the anterior part of their bases is above the centre of the orbits. The posterior part of their bases is slightly behind the posterior margin of the orbits ( Fig. 1 View FIG ). In frontal view the bases of the horn-cores are almost parallel to each other, but they diverse upwards and finally their tips are sub-parallel to each other ( Fig. 3A, B View FIG ). In lateral view the horncore axis does not curve but remains almost straight ( Fig. 3D View FIG ). There is a main keel that starts postero-laterally and runs almost the whole horncore. In some of the less massive horn-cores (PG 95/1514, PG 95/1510, PG 95/1509), this keel is less acute and less distinct than in the more massive horn-cores (PG 95/1502a, PA 909/91, PA 1412/91). Anterior to this keel there is a wide groove, variable in development, but always clearly distinct at the base of the horn-core. In some specimens there is another narrow groove, posterior to the keel. The horn-cores taper gradually distally and their section is ovoid throughout their length ( Fig. 4 View FIG ). The pedicels are short and they almost disappear posteriorly. The horncores (especially the more massive ones) protrude anteriorly over the pedicels, forming an anterior V-shaped extension over them. There is a deep postcornual fossa, independent in development from the horn-core robustness. Two almost complete horn-cores (the right horn-core of PG 95/1514, and PA 909/91) trace one complete gyre, and their total length (in straight-line) can be estimated to 120-130 mm. Two horn-cores (PG 88/1540, PG 88/1583) are broken at their bases and show that the pedicels are not hollowed.
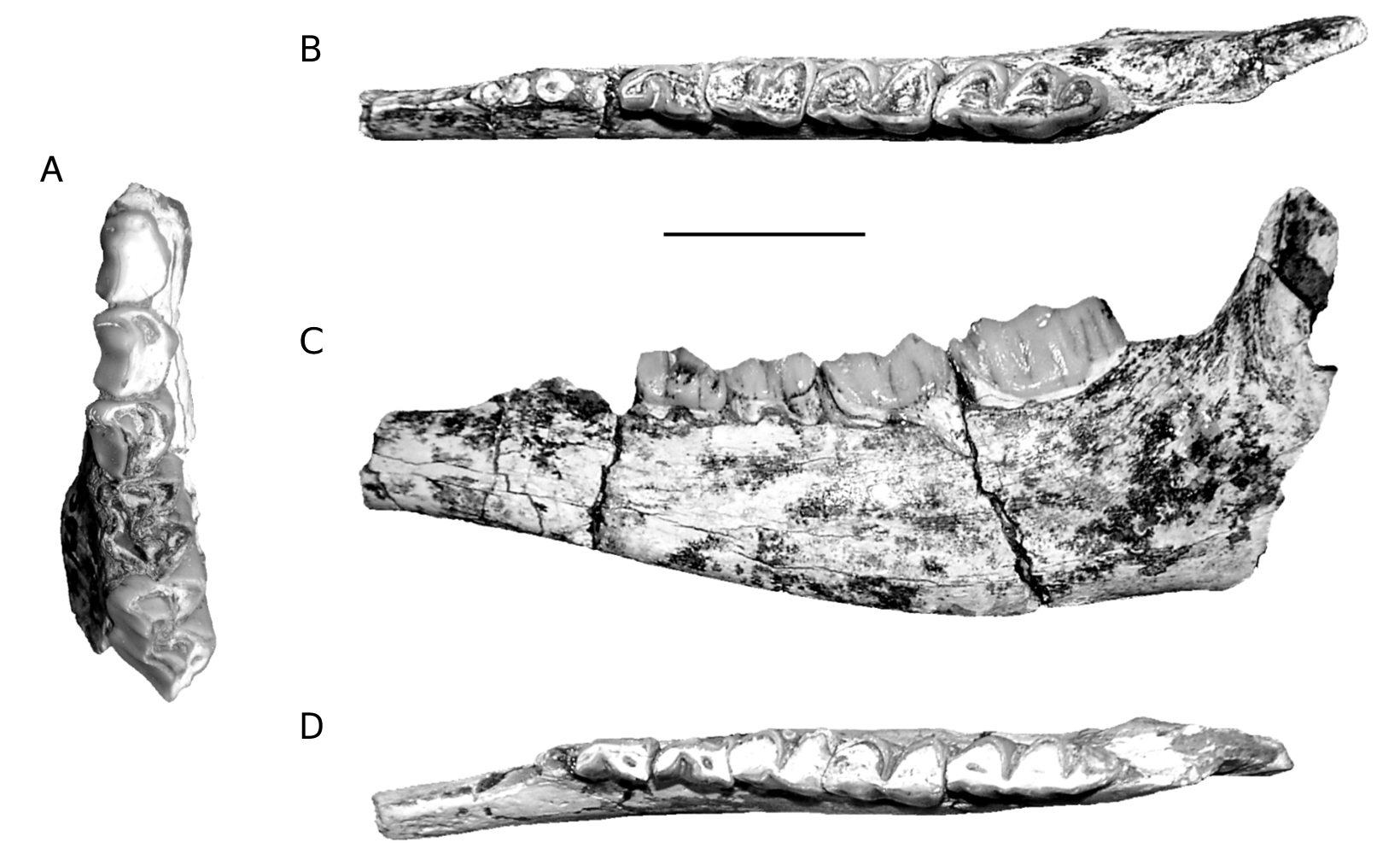
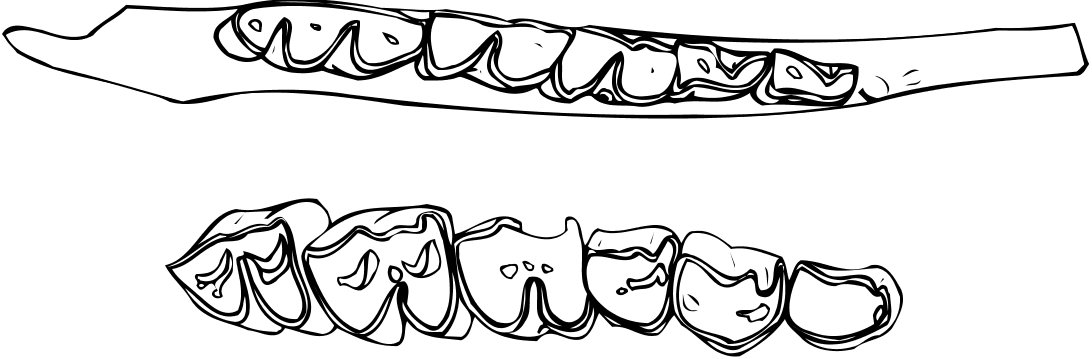
Upper dentition
The upper tooth row is characterised by the long premolar series relative to the molars (Annexe: Table 4; Figs 5A View FIG ; 6C View FIG ). P2 is moderately elongat- ed, its parastyle is very small and does not project labially, and its paracone rib is thick, situated just behind the parastyle. The protocone is less developed than the hypocone and less projected lingually. The protocone and the hypocone are separated lingually by a slight groove, situated almost opposite the rib of the paracone. On the occlusal surface, the anterior fossa is very reduced and situated at the anterior part of the protocone, very close to the enamel. P3 is similar to P2 but less elongated (Annexe: Table 4), while its para-
A B C anterior
style is more developed. The paracone rib is very thick. As in P2, a slight groove separates lingually the protocone from the more developed hypocone. The position and the development of its anterior fossa resemble those of P2. A trace of the posterior fossa can be observed on the left P3 of PG 95/1502a. The P4 has a thick parastyle while its paracone rib is only slightly developed. A very clear groove separates lingually the protocone from the hypocone. The protocone extends lingually more than the hypocone. Its anterior fossa is like that of P2 and P3, but a small posterior fossa is also visible. The metastyle is thinner than the parastyle. The molars have a thick parastyle, the rib of the labial wall of the paracone is moderately developed, and the labial wall of the metacone is only slightly convex. The M1 and M2 of PG 95/1502a have faint entostyles (absent in PA 3509/91). The metastyles and the mesostyles are thinner than the parastyles. The M1 and M2 have a central fossa. A fold at the posterior fossa, or a small additional fossa at the posterior part of the hypocone can be seen on the occlusal surface of some molars.
Lower dentition
Besides the two mandibles (PG 95/1502b & c) found in connection with the skull, we refer to O. rothii another mandible (PA 3519/91). Despite their slightly smaller dimensions (Annexe: Table 5), the teeth of this mandible show many morphological similarities to PG 95/1502b & c, even in details. Because of the lack of a complete mandible the length of the tooth row and that of the premolar row cannot be calculated with accuracy. These parameters are given approximately (Annexe: Table 5). As in the upper tooth row, the lower premolar row is long relative to the molars (Annexe: Table 5; Figs 5 View FIG B-D; 6A, B). The mandibular corpus is not especially deep below m3, and its depth decreases gradually anteriorly. On the labial surface of the mandibular corpus and slightly behind m3 there is a well developed angular tuberosity (rugose area) for the masseter muscle. The p3 is slightly smaller than the p4 (Annexe: Table 5). The paraconid is not distinct from the parastylid, and the metaconid is obliquely direct- ed and connected to the posterior part of the tooth. The valley between the paraconid and the metaconid remains wide and open down to the base of the crown. There is a groove in front of the hypoconid. The p4 is similar to p3, but the metaconid has anteroposterior direction and the groove in front of the hypoconid is more distinct. The lower molars have thick parastylid, but the metastylid and the entostylid are weakly developed. The lingual wall of the metaconid and the entoconid are simple and slightly convex without strong ribs. There are no goat folds. There is a vertical groove on the labial wall of the posterior lobe of m1. The ectostylids are more developed on the anterior molars. There is a second ectostylid in front of the third lobe of m 3 in PG 95/1502c, but not in PG 95/1502b. In PA 3519/91, the m3 is slightly broken in this area, but a trace of an ectostylid in front of the third lobe is still present. The third lobe of m3 is relatively large and has a central fossa.



| MNHN |
Museum National d'Histoire Naturelle |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.