
Orobothriurus ramirezi, Ochoa & Ojanguren Affilastro & Mattoni & Prendini, 2011
|
publication ID |
https://doi.org/ 10.1206/359.1 |
|
publication LSID |
lsid:zoobank.org:pub:870DFDFC-45BF-49CF-8C3F-2527DB611CFC |
|
DOI |
https://doi.org/10.5281/zenodo.4618608 |
|
persistent identifier |
https://treatment.plazi.org/id/08318541-1F41-FF99-18F6-FA21FB954425 |
|
treatment provided by |
Felipe |
|
scientific name |
Orobothriurus ramirezi |
| status |
sp. nov. |
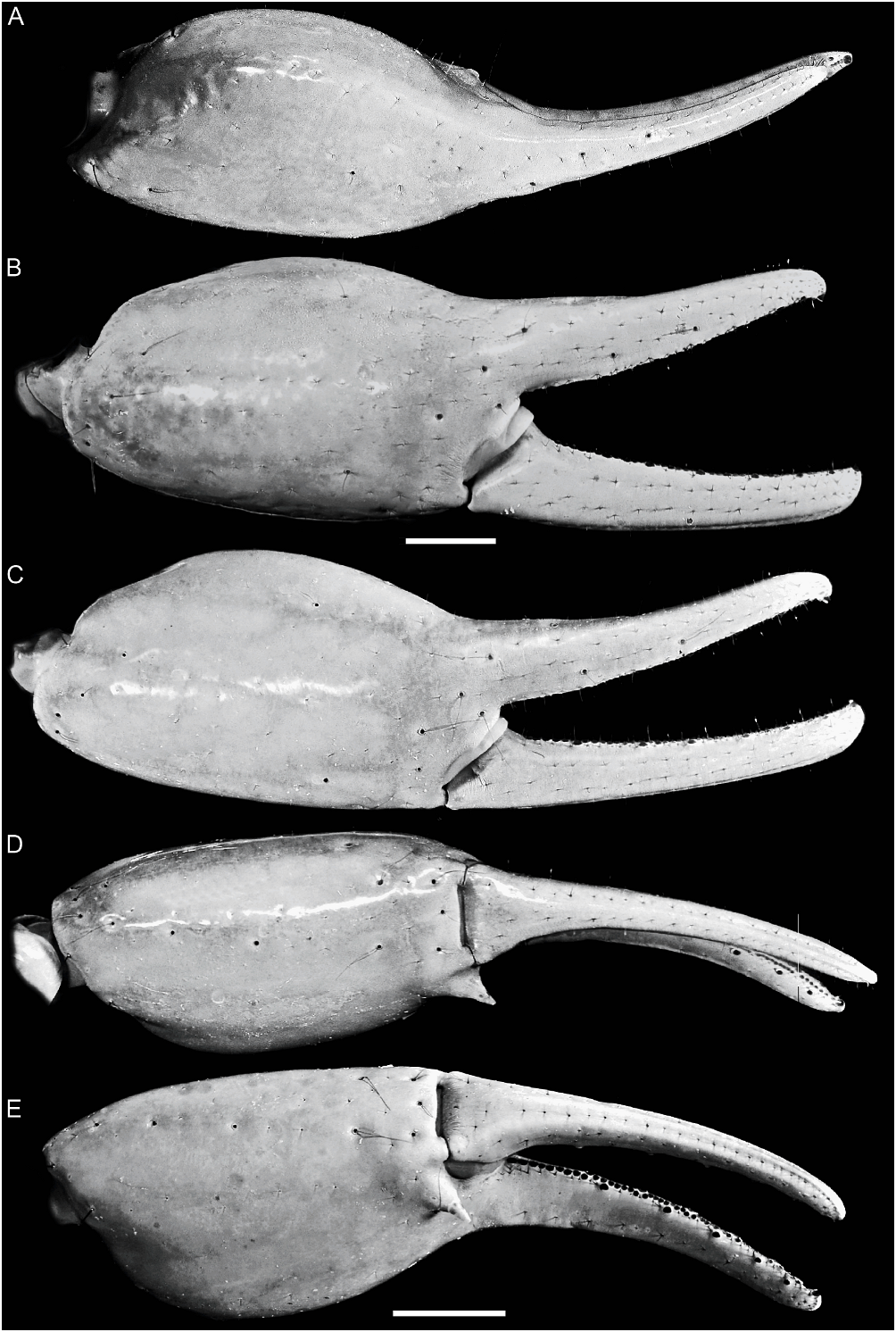
Orobothriurus ramirezi View in CoL , n. sp. Figures 16A, B View Fig , 18C View Fig , 20C, F View Fig , 21G View Fig , 24E View Fig , 44–46 View Fig View Fig View Fig , 47A, B View Fig , 54 View Fig ; table 3
TYPE MATERIAL: CHILE: Region IV (Coquimbo): Elqui Province: Holotype ♂ (MACN-Ar), Paso del Agua Negra , between Juntas and international border, 30 ° 12911.50S 69 ° 54957.50W, 3734 m, 27.i.2005, C.I. Mattoni and A. Ojanguren Affilastro, UV detection, specimens walking actively near a ‘‘vega’’ (small stream), extremely wet. Paratypes: same data, 4 ♂, 1 ♀ ( AMNH), 2 ♂, 1 ♀ ( LBRE), 1 ♂, 1 ♀ (MACN-Ar), 1 ♂ ( MHNC); same data, except 30 ° 16914.50S 69 ° 58927.90W, 3295 m, 1 ♂, 1 ♀, 5 juv. ( AMNH), 1 ♀, 2 subad. ♂ ( AMNH [ LP 4305 ]), 1 ♂, 1 ♀, 3 juv. ( LBRE), 1 ♂, 1 ♀, 2 juv. (MACN-Ar), 1 ♀, 2 juv. ( MHNC), 1 ♂, 2 juv. ( MZUC) .
ADDITIONAL MATERIAL: CHILE: Region IV (Coquimbo): Elqui Province: Mine El Indio, Cancha Sky base camp [29 ° 519380S 70 ° 029120W, 3225 m], v.1996, H. Vásquez C., pitfall trap, 1 ♀, 1 juv. ( ULS) ; Mine El Indio, Mangueras, ca. 4000 m, 1 juv. ( AMNH), 1 juv. ( LBRE) ; Mine El Indio, Pastos Largos [29 ° 499S 70 ° 039W, 3600 m], i.1995, H. Vásquez C., pitfall trap, 1 ♀, 1 juv. ( ULS), 6.xi.2003, L. Prendini, C.I. Mattoni and J.A. Ochoa, UV detection, very cold and windy, full moon, alpine vegetation on scree slope near small stream, specimen sitting in open ground, 1 subad. ( AMNH [ LP 2402 ]) ; Mine El Indio, Quebrada El Negro [29 ° 479060S 70 ° 009000W, 3808 m], 6.xi.2003, C.I. Mattoni, L. Prendini, and J.A. Ochoa, UV detection and diurnal rock rolling, puna vegetation, low bushes and grass tufts, hard, rocky ground with bare patches, stones in places, at night, very cold and windy, full moon, specimens on rocky slope (juv. under stone), 1 ♀, 1 juv. ( AMNH [ LP 2401 ]) ; Mine El Indio, Tambo [29 ° 489S 69 ° 589W, 4030 m], ii.1997, H. Vásquez C., pitfall trap, 1 ♂, 1 ♀, 1 juv. ( ULS) ; Paso del Agua Negra, between Juntas and international border, 30 ° 12911.50S 69 ° 54957.50W, 3350 m, 6.iii.2006, A. Ojanguren Affilastro, L. Compagnucci, and C. Cuezzo, UV detection, 1 ♂, 1 ♀, 1 subad. ♀, 1 juv. (MACN-Ar) .
ETYMOLOGY: This species is named in honor of the Argentine arachnologist Martín J. Ramírez (MACN) for his contributions on the biodiversity and systematics of South American arachnids.
DIAGNOSIS: Orobothriurus ramirezi is closely related to O. alticola (fig. 5), based on similar pigmentation pattern, metasomal carination, and hemispermatophore morphology. The two species may be separated by the distal crest of the hemispermatophore, which is comparatively more developed, occupying almost the entire apex of the lamina, in O. ramirezi , than in O. alticola , in which it occupies only half the apex (figs. 36I, J, 47A, B). The telson vesicle of O. ramirezi is more rounded, especially in females (telson length/width ratio: ♂, 2.65– 2.94, mean 5 2.78; ♀, 2.13–2.37, mean 5 2.27; figs. 23G, 24E) than that of O. alticola (telson length/width ratio: ♂, 2.87–3.21, mean 5 3.07; ♀, 2.44–2.55, mean 5 2.49; fig. 23A). The dorsal surface of the femur and ventral surface of the metasoma are more pigmented, with broader stripes, in O. ramirezi than O. alticola . The metasomal carinae, especially the VL and VSM carinae of segments I and V, and the DL carinae of segment V, are more developed in males of O. ramirezi than O. alticola (fig. 20A, C).
DESCRIPTION: Based on holotype ♂ and paratypes. Measurements of holotype ♂ and paratype ♀ recorded in table 3.
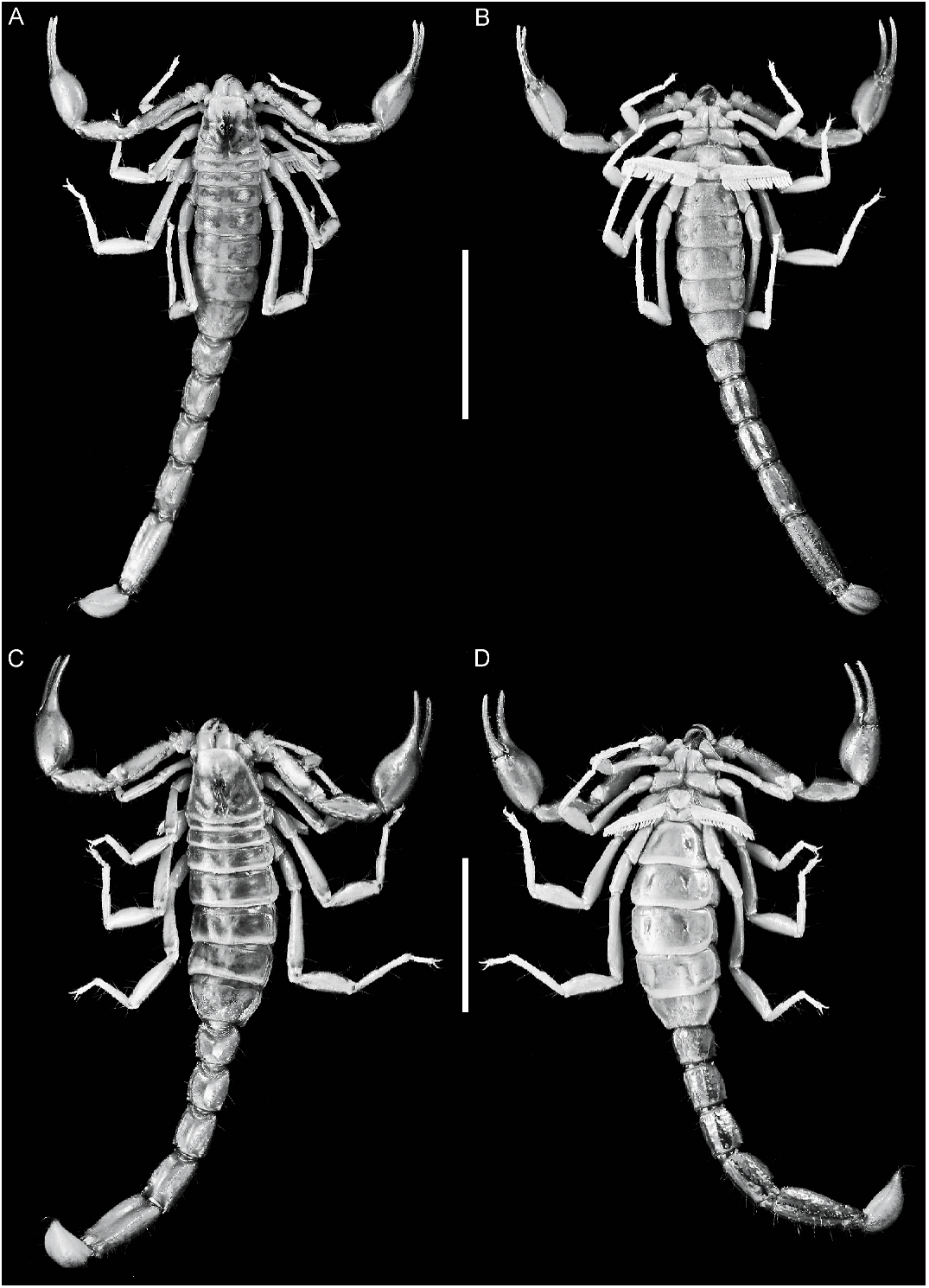
Total length: ♂, 27.3–36.0 mm (n 5 13, mean 5 31.6 mm); ♀, up to 37.8 mm.
Color: General color yellowish with dark brown spots. Carapace, anterior margin unpigmented (fig. 44A, C); lateral margins with two spots laterally and two smaller spots posterolaterally, intermediate area with faint reticulate pigmentation; median ocular tubercle and lateral ocelli dark brown or black; posterior half of anteromedian longitudinal sulcus, median ocular tubercle, and posteromedian longitudinal sulcus pigmented. Chelicerae unpigmented. Pedipalp coxa unpigmented; trochanter, dorsal and lateral surfaces faintly pigmented; femur, ventral surface unpigmented, dorsal surface pigmented, except for oval unpigmented area, occupying up to twothirds of segment distally; internal and external surfaces unpigmented, except for faint stripes along VL and VI carinae; chela manus with six complete longitudinal stripes along carinae, joining at base of movable finger; fingers unpigmented except basally. Legs, coxa and trochanter unpigmented; femur, dorsal and prolateral surfaces sparsely spotted, spots becoming larger near articulation with patella; patella, prolateral, dorsal and ventral surfaces sparsely pigmented; basitarsi and telotarsi, dorsal and prolateral surfaces faintly spotted. Tergites I–VI each with two faint spots sublaterally, reaching pretergites on III–VI and delimiting unpigmented median stripe; VII with two small, faint, spots of pigmentation submedially (fig. 44A, C). Sternum, genital opercula, pectines, and sternites III–VI unpigmented; sternite VII with two, often faint, spots sublaterally. Metasomal segment I, dorsal surfaces each with faint subtriangular spot medially; lateral surfaces pigmented between ML and LIM carinae; ventral surface with two narrow VL stripes becoming slightly broader in posterior half (fig. 44). Segments II–IV, dorsal surfaces each with small subtriangular spot medially; lateral surfaces densely pigmented below ML carinae; ventral surfaces each with three separate, narrow stripes (one VM and two VL), becoming broader in posterior half. Segment V, dorsal surface unpigmented; lateral surfaces with weak reticulate pigmentation; ventral surface with three separate stripes (one VM and two VL), becoming broader posteriorly. Telson vesicle, dorsal surface unpigmented (♀) or with faint longitudinal stripe (♂); ventral surface with three broad dark stripes (two VL and one VM), separated by two narrow unpigmented stripes; aculeus sclerotized, dark reddish brown.
Chelicerae: Movable finger with two subdistal teeth.
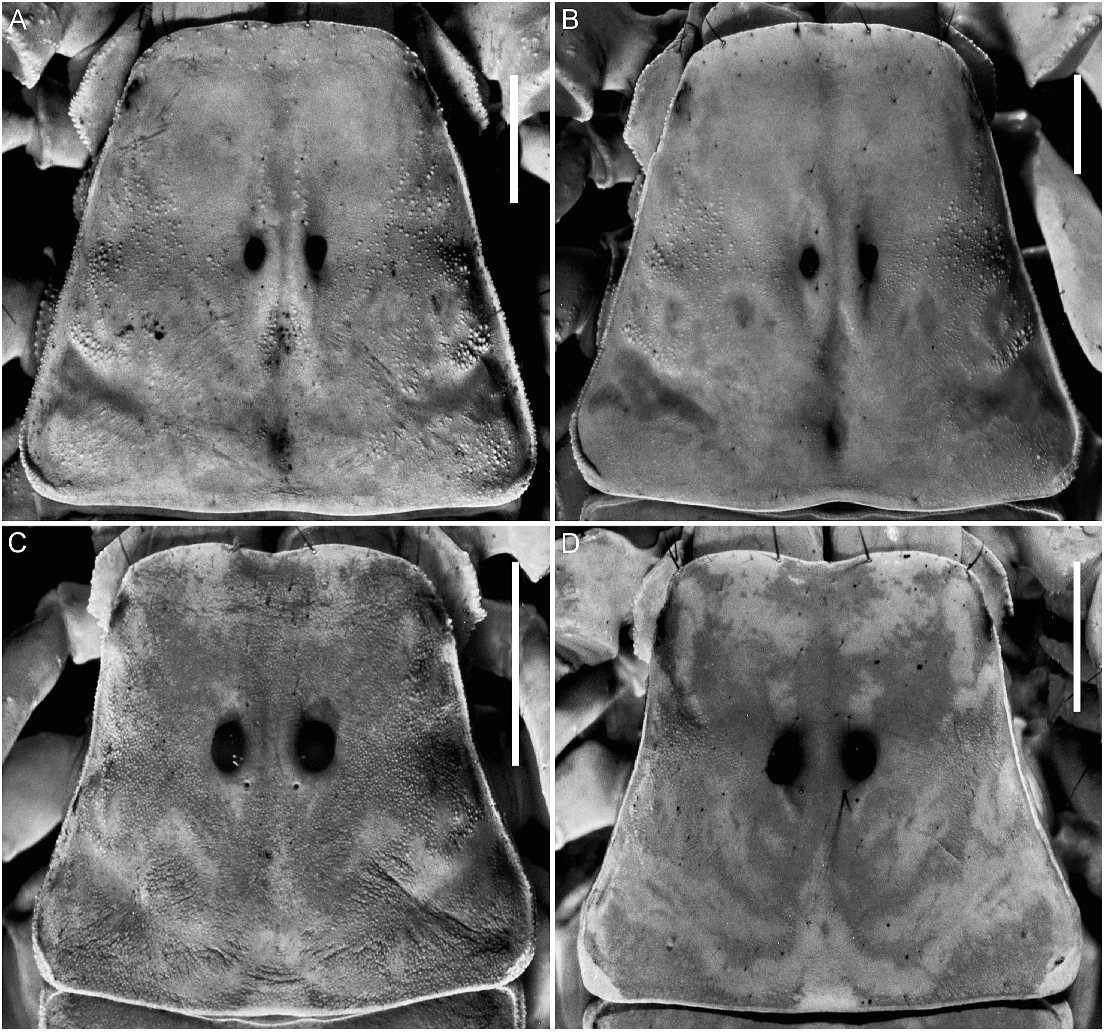
Carapace: Median surfaces smooth; lateral margins finely granular, less so in ♀. Anterior margin linear, without median notch (fig. 16A, B). Anteromedian longitudinal sulcus obsolete; median ocular, posteromedian longitudinal and posterolateral sulci well developed. Median ocular tubercle raised, situated anteromedially; median ocelli two ocular diameters apart.
Pedipalps: Femur, DI and VI carinae complete, finely granular; DE carina obsolete, smooth (fig. 38A); internal surface finely and sparsely granular medially; other intercarinal surfaces smooth. Patella, DI and VI carinae obsolete, finely granular (♂) or smooth (♀) (fig. 38B–D); internal surface with two prominent granules distally, the larger adjacent to trichobothrium i; other intercarinal surfaces smooth.
Pedipalps: Femur, DI and VI carinae complete, granular (fig. 45A); DE carina complete, granular in distal third to distal half of segment, other carinae smooth; ventral surface sparsely granular, more densely granular in proximal half. Patella, DI and VI carinae complete, granular, less so in ♀ (fig. 45B–D); DPP carina comprising small granules proximally; VPP carina absent; internal surface coarsely granular with prominent granule adjacent to trichobothrium i near DI carinae; other surfaces smooth. Chela manus slender, fingers relatively elongated (fig. 46); length/width ratio: ♂, 3.58–4.14 (n 5 12; mean 5 3.84), ♀, 3.31– 3.9 (n 5 6, mean 5 3.62); length/height ratio: ♂, 2.94–3.75 (n 5 12; mean 5 3.3), ♀, 3.06– 3.4 (n 5 6, mean 5 3.29); DMA, DI, and VM carinae obsolete, other carinae absent; intercarinal surfaces smooth; internal surface with acuminate apophysis (♂) or low bulge (♀) near articulation of movable finger (fig. 46A, D); fingers, dentate margins each with median denticle row and 4–5 pairs of internal and external accessory denticles.
Trichobothria: Femur with 3 trichobothria, patella with 19, chela with 27. Chela trichobothrium Et 3 situated proximal to Est (fig. 46).
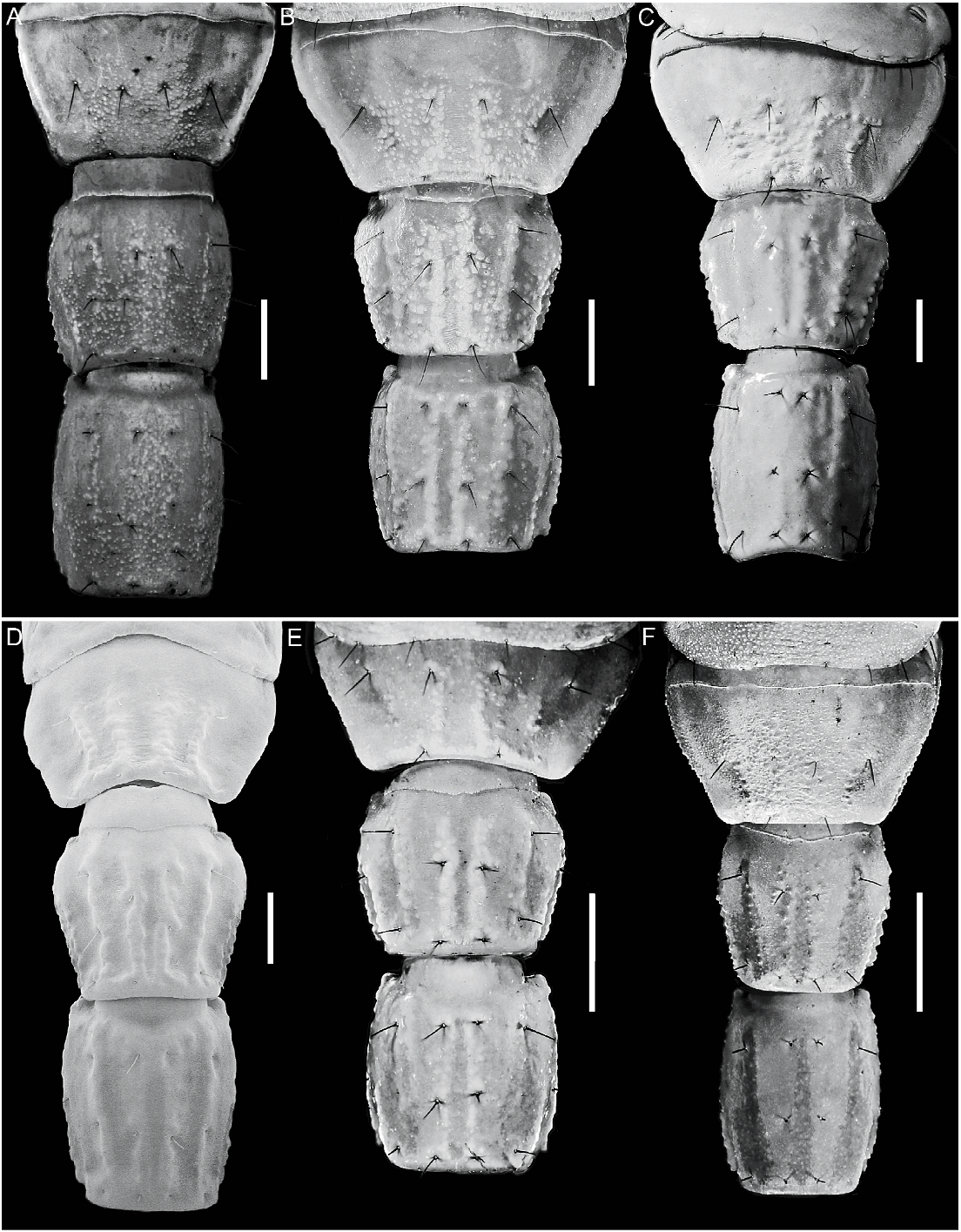
Tergites: Tergites I–VI, surfaces finely granular (♂) or smooth (♀). Tergite VII tetracarinate, paired DL carinae restricted to posterior two-thirds of segment, paired DSM carinae to posterior half; intercarinal surfaces coarsely granular, other surfaces finely granular.
Legs: Femur and patella, prolateral surfaces finely granular, retroventral surfaces smooth. Femur, ventral carinae weakly developed; other carinae absent. Patella acarinate. Telotarsi, pro- and retroventral rows of spiniform macrosetae with following counts on leg I, 1/1; II, 2/2; III and IV, 3/3.
Pectines: Pectinal tooth count: ♂, 21–26 (n 5 33, median 5 23); ♀, 15–20 (n 5 34, median 5 18).
Sternites: Sternites III–VI, surfaces smooth (♀) or finely corrugated (♂), especially in posterior half; spiracles small, narrow. Sternite VII, surface smooth; VL and VSM carinae obsolete (fig. 18C).
Metasoma: Segment I, DL carinae complete, moderately granular; ML carinae complete, moderately granular in posterior two-thirds; one pair of ML macrosetae; LIM carinae complete, moderately granular in posterior half; VL and VSM carinae well developed (especially in ♀, fig. 19C), complete, granular; two pairs of VSM and VL macrosetae. Segment II, DL carinae complete, granular; ML carinae complete, granular; one pair of ML macrosetae; LIM carinae restricted to the posterior third; VSM and VL carinae well developed, complete (♀, fig. 18C) or obsolete (♂); usually two pairs of VL and three pairs of VSM macrosetae. Segment III, DL and ML carinae complete, moderately granular; one pair of ML macrosetae; LIM carinae reduced to two or three granules in posterior quarter; VL and VSM carinae obsolete; two or three pairs of VL and three pairs of VSM macrosetae. Segment IV, DL carinae complete, granular; one pair of DL macrosetae; ML carina reduced to few distal granules (♀) or obsolete (♂); one or two pairs of ML macrosetae; LIM, VL and VSM carinae absent; three pairs of VL and three or four pairs of VSM macrosetae. Segment V, length/width ratio, ♂, 2.29–2.69 (n 5 12; mean 5 2.44), ♀, 1.9–2.2 (n 5 6; mean 5 2.02); DL carinae complete, more evident in anterior half; lateral margin smooth; ML carinae absent; four pairs of ML macrosetae; VL carinae complete (♂) or restricted to posterior three-quarters (♀); VL and VSM carinae situated close together, fused in posterior third; VM carina complete, obscured by surface granulation in posterior half (fig. 20C); four pairs of VL and VSM macrosetae; two pairs of macrosetae along posterior margin.
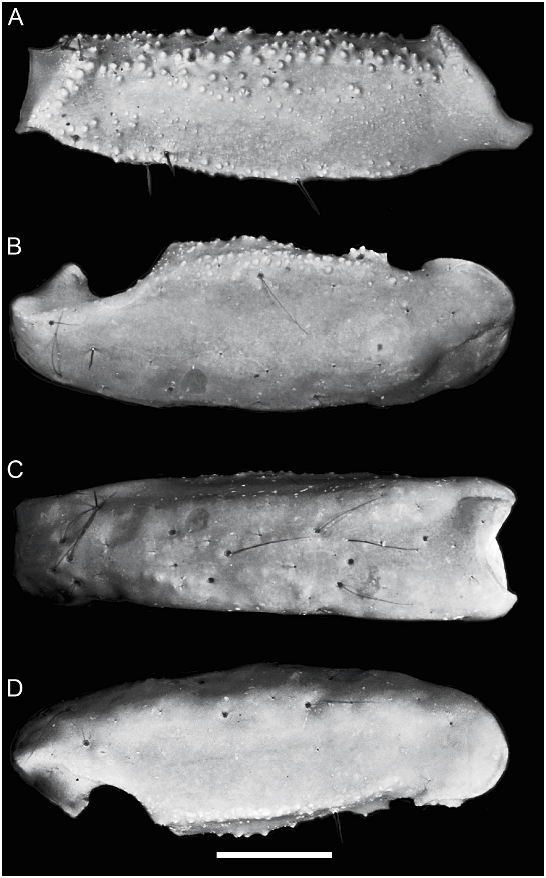
Telson: Length/width ratio: ♂, 2.65–2.94 (n 5 13, mean 5 2.78); ♀, 2.13–2.37 (n 5 10, mean 5 2.27). Length/height ratio: ♂, 3.33– 3.89 (n 5 13, mean 5 3.53); ♀, 2.63–3 (n 5 10, mean 5 2.86). Vesicle elongated (♂, fig. 23G, 24G) or globose (♀, fig. 24E); dorsal surface smooth, slightly concave, gland not apparent (♂); ventral surface smooth (♂) or granular (♀). Aculeus short and curved, more so in ♀.
Hemispermatophore: Apex very well developed; distal crest very well developed, occupying almost three quarters of apex, curved like ventral margin. Frontal crest weakly developed, less than half length of lamina; basal part oblique; distal part short, parallel to ventral margin of lamina, slightly undulated. Basal lobe, terminal process extending almost to constriction of frontal crest (fig. 47A, B).
DISTRIBUTION: Orobothriurus ramirezi is known only from localities above 3200 m in the Cordillera de Doña Ana of the Andes, in the Elqui Province of Region IV (Coquimbo), Chile (fig. 54).
ECOLOGY: Most specimens were collected near wetland ecosystems termed ‘‘vega’’ or ‘‘humedal’’ ( Squeo et al., 1994; CepedaPizarro, 2004) in the high Andes. These habitats are characterized by an accumulation of water resulting in permanently flooded soil. The temperature at 3750 m varies from ‾ 17 ° C in winter, to more than 24 ° C in summer, but temperatures below zero are possible throughout the year ( Cepeda-Pizarro, 2004). Precipitation in the form of snow, which may accumulate up to 8 m in some areas, occurs mainly during winter. The vegetation corresponds to a steppe with diverse altitudinal levels: shrub steppe (or sub-Andean floor) at 2700–3500 m, subshrub steppe at 3500–4250 m, and highAndean steppe at 4250–4450 m, above which there is no vegetation ( Squeo et al., 1994; Cepeda-Pizarro, 2004). Orobothriurus ramirezi has not been found in the upper altitudinal level, where the ‘‘vega’’ ecosystem is absent. It is sympatric with another bothriurid, Brachistosternus perettii Ojanguren Affilastro and Mattoni, 2006 , that apparently prefers rocky areas ( Ojanguren Affilastro and Mattoni, 2006). The most closely related species, O. alticola , inhabits similar habitats on the western side of the Andes. Despite the proximity of the nearest locality of O. alticola (only 16 km as the crow flies), the two species are allopatric, separated by the highest peaks of the Andes, the lowest pass being 4700 m, where life is almost absent. Most adult males of O. ramirezi were collected at night with UV light detection, sitting or walking near streams, probably in search of females, suggesting that the period when they where collected (late January, midsummer in the Southern Hemisphere) is the active season for this species.





| AMNH |
American Museum of Natural History |
| MHNC |
Museo de Historia Natural de Concepcion (Chile) |
| MZUC |
Museo de Zoologia, Universidad de Concepcion |
| ULS |
Universidad de La Serena |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.