
Pheretima longiprostata, Aspe, Nonillon M. & James, Samuel W., 2014
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3881.5.1 |
|
publication LSID |
lsid:zoobank.org:pub:FE9048E9-DE3A-4502-A95E-27EE8F706AC3 |
|
DOI |
https://doi.org/10.5281/zenodo.5670425 |
|
persistent identifier |
https://treatment.plazi.org/id/5B458787-FF92-FF87-FF5A-FB2FE48EBF5F |
|
treatment provided by |
Plazi |
|
scientific name |
Pheretima longiprostata |
| status |
sp. nov. |
Pheretima longiprostata n. sp.
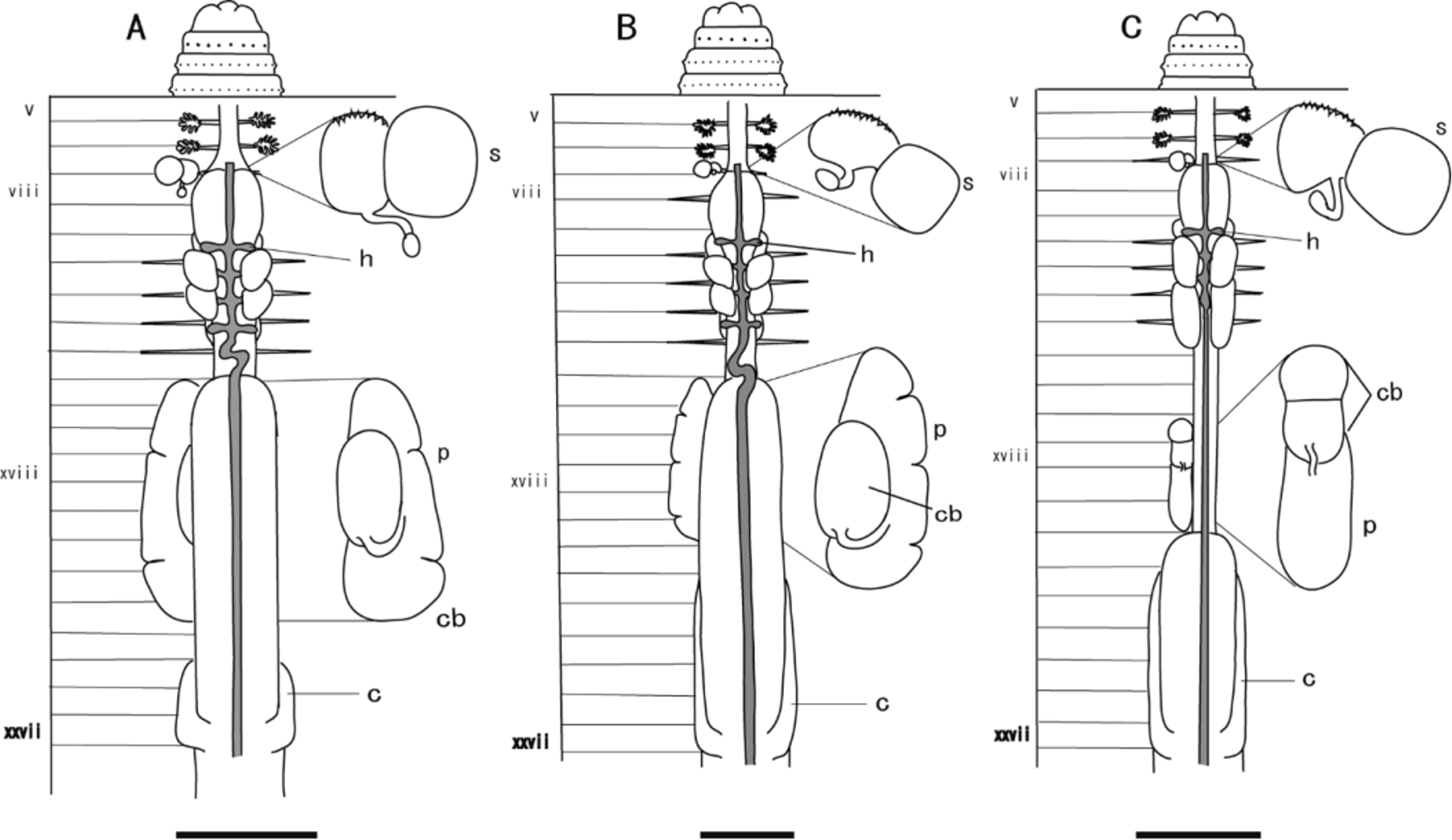
( Fig. 6 View FIGURE 6 A, Table 2)
Material examined. Holotype: adult (NMA 4519) Brgy Small Potongan, municipality of Concepcion, Misamis Occidental Province, Mt. Malindang Range (8º24'04"N, 123º36'47"E), 900 m asl., Mindanao Island, Philippines, coll. Nonillon Aspe, Nolan Aspe, M. Lluch, and J. Adeva, Feb. 18–25, 2004. Paratype: one adult (NMA 4539), same collection data as for holotype.
Etymology. The species name derives from the Latin 'longus' (long) and 'prostata' (prostate gland), referring to the long prostate glands.
Diagnosis. Very small, yellowish-brown worm reaching 37–41 mm in adult length; first dorsal pore at 11/12; one pair of spermathecal pores at 7/8; spermathecae small with oval ampulla and stout duct; intestinal origin in xv; elongate prostate glands extending from xv to xxiii; short caeca extending from xxvii to xxv; 20 intestinal vessels.
Description. In living animals, dorsum light yellowish-brown anteriorly, fading posteriorly; equators nonpigmented; ventral side non-pigmented; clitellum gray. Length 37–41 mm (n=2 individuals); diameter 3.5 mm at x, 3 mm at xx; body cylindrical in cross-section, tail tapering; 82-90 segments. First dorsal pore at 11/12; paired spermathecal pores at 7/8, 0.16 circumference apart ventrally; female pore single on xiv; openings of copulatory bursae paired on xviii, 0.16 circumference apart ventrally, 0–5 setae between openings. Clitellum annular, extending from xiv to xvi. Setae evenly distributed, 35–40 setae on vii, 37–47 setae on xx, dorsal gap present, ventral gap absent.
Septa 5/6/7/8 thin, 8/9/10 absent, 10/11–13/14 thin. Dense tufts of nephridia on anterior faces of 5/6 and 6/7; nephridia of intestinal segments located mainly on body wall at anterior and posterior faces of septa, at septum/ body wall junction. Gizzard large, extending from viii to x; esophagus with lamellae from xi to xiii; intestinal origin in xv; caeca originate in xxvii, extending forward to xxv; typhlosole a simple fold of about 1/4 lumen diameter, originating at 26/27. Intestinal wall with 20 longitudinal blood vessels.
Hearts in x to xiii, esophageal; commissural vessels in vi, vii, and ix, lateral; supra-esophageal vessel extends from x to xiii; extra-esophageal vessels join ventral esophageal wall in x, receive efferent parieto-esophageal vessels in xiv.
Ovaries and funnels free in xiii; paired spermathecae pre-septal in vii, with nephridia on ducts; each spermatheca with transversely placed oval ampulla, short thick muscular duct with slight bulge for diverticulum attachment, single stalked diverticulum attached to posterior face of duct, terminating in ovate receptacle; stalk longer than receptacle. Spermathecal duct walls thick, complexly corrugated. Male sexual system holandric; testes and funnels enclosed in paired sacs in x and xi; seminal vesicles in xi and xii, each with short dorsal lobe; vasa deferentia slender, free from body wall, passing around lateral face of copulatory bursae en route to ental end of prostatic ducts; relatively large and extensive prostates in xv to xxiii, each racemose, bilobed, wrapped around lateral margins of copulatory bursa; muscular duct attached over half of its length to medial face of copulatory bursa, running along surface and entering at apex. Single ductlets from anterior prostatic lobes join vas deferens; these join one or two posterior ductlets at common junction with muscular prostatic duct. Copulatory bursae ovate, extending from xvii to xx, attached to body wall; coelomic surface of copulatory bursa muscular, secretory diverticula lacking. Interiors of bursae lacking pads; large penis from dorsal apex fills most of bursal interior; copulatory bursae lack penial sheaths.
Remarks. Pheretima longiprostata n. sp. belongs to the P. sangirensis group of Sims & Easton (1972). Individuals of this species were by far the smallest among the P. sangirensis group at Malindang, and are among the smallest Pheretima reported from the Philippines. In size and color, P. longiprostata is most similar among the Malindang species to P. vergrandis n. sp. (see description below). However, the two species differ in the location of the first dorsal pore, the number and position of spermathecae and male pores, the length of the caeca, and the size and shape of the prostate glands. Pheretima longiprostata has the most elongate prostate glands relative to body size among the known Philippine Pheretima species, extending 9 segments from xv to xxiii (see Table 2). Pheretima wati n. sp. has prostate glands similar in size to those of P. longiprostata , extending 8 segments from xv to xxii, but the two species differ in size, coloration, location of the first dorsal pore, septal arrangement, length of the caeca, the shape of the spermathecae, and the number of intestinal vessels. Prostate glands function to produce fluids in which sperm cells can be transferred between worms during copulation ( Edwards & Bohlen 1996), and these glands may also help to provide the cocoon with nutrients. However, the physiological significance of long or large prostate glands is not clear. Some species in Pheretima and in Amynthas ( Kinberg, 1867) lack prostate glands; in these cases, the clitellum may be larger so as to supply the necessary nutrients to the cocoon. This remains to be investigated.
Occurrence. Pheretima longiprostata was uncommon, with only a few individuals found in disturbed forest in Brgys Small Potongan and Sibucal. We did not observe it above 1067 m asl. Individuals occurred in the soil and on rotten logs (Table 1).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.