
Phrynomedusa appendiculata ( Lutz, 1925 )
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5087.4.2 |
|
publication LSID |
lsid:zoobank.org:pub:39E2E033-AB5B-44A2-9438-33360B994FE4 |
|
DOI |
https://doi.org/10.5281/zenodo.5832858 |
|
persistent identifier |
https://treatment.plazi.org/id/0386879E-5F79-FF90-FF25-462A4C2D1ACA |
|
treatment provided by |
Plazi |
|
scientific name |
Phrynomedusa appendiculata ( Lutz, 1925 ) |
| status |
|
Phrynomedusa appendiculata ( Lutz, 1925) View in CoL
( Figs. 1–3 View FIGURE 1 View FIGURE 2 View FIGURE 3 )
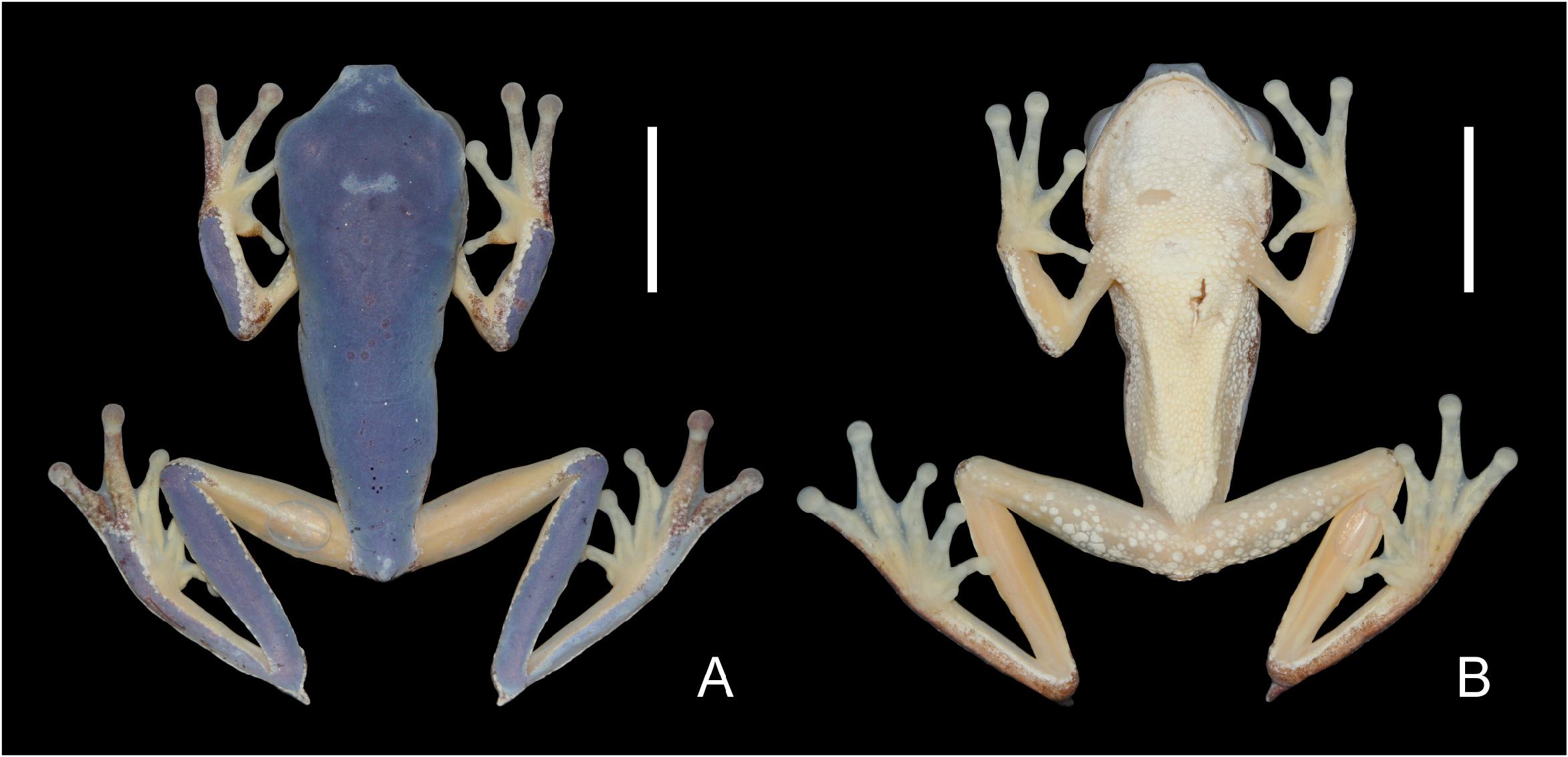
Description of the specimen. Adult male (MZUSP 159234; field number DT 2969, Figs. 1 View FIGURE 1 , 2 View FIGURE 2 ), collected by D. Pavan on 04 December 2011. Slender body; head wider than long (33% of SVL); snout acuminate in dorsal view, oblique in lateral view, internarial distance 55% of interorbital distance; nostrils lateral and slightly protuberant; canthus rostralis curved (concave); loreal region oblique; upper lip dermal fringe absent; interorbital region flat; eye large; palpebral membrane not reticulated; eye diameter 82% of interorbital distance; supratympanic fold well defined, extending posteroventrally to above insertion of arm; tympanum visible; tympanic membrane weakly differentiated from surrounding skin; tympanic annulus round, with its uppermost portion obscured by the supratympanic fold, tympanum diameter 47.5% of eye diameter, separated from eye by 21% of tympanum diameter; tongue cordiform, anteriorly attached; dentigerous process of vomer absent; choanae large and elliptical widely separated from each other; vocal sac single, subgular, not externally evident.
Forearm more robust than upper arm; forearm length 80% of hand length; inner metacarpal tubercle single and elliptical; outer metacarpal tubercle absent; subarticular tubercles rounded; supernumerary tubercles absent; dermal folds smooth on postaxial margin of forearm; webbing formula: I 2 + – 21/2 II 2 – 3 – III 22/3 – 2 IV; finger length I <II <IV <III; adhesive discs rounded, more distinct in fingers III and IV; keratinized nuptial pad present at dorsal and preaxial surface of finger I, not extending ventrally.
Hind limbs robust; tibia length 51% of SVL; foot length 72.5% of tibia length; triangular calcar appendix present, slender and long, parallel with tarsus sagittal axis and occupying approximately half of tibio-tarsal articulation; smooth lateral fringes present on external face of tibia and feet, less developed at the toe V; inner metatarsal tubercle single and elliptical; outer metatarsal tubercle absent; subarticular tubercles single and rounded; supernumerary tubercles absent; webbing formula I 2 + – 2 II 2 – – 2 + III 1 1/2 – 2 IV 2 – – 1 V; toe length I <II <III <V <IV; adhesive discs rounded, more distinct on toes IV and V.
Skin on the dorsal surfaces of head, body and limbs smooth; skin on ventral surfaces smooth to areolate on throat, areolate in chest and belly and almost smooth with few areoles in the posterior region of thigh and arm; cloaca directed posteriorly above dorsal plane of thighs; osteoderms parotoid and dorsolateral glands absent.
Measurements of the specimen. SVL 31.4; HL 10.5; HW 11.6; IN 2.7; IO 4.9; ED 4.0; EN 3.3; TD 1.9; FA 7.9; HA 9.9; TH 15.4; TL 16.0; FL 11.6.
Coloration. The color in life for P. appendiculata has been described based on a sketch ( Cochran, 1955) and recently redescribed with the aid of old photographs from an individual recorded in Santo André, state of São Paulo ( Baêta et al. 2016). Here we complement the description of color in life variation for this species based on the newly recorded individuals ( Figs. 2 View FIGURE 2 , 3 View FIGURE 3 ). Dorsal surface of the head and body, lateral surface of head and loreal region bright green. Upper and lower lips grayish-white, often delimited above by a thin darker stripe. Grey bordered nostrils. Lateral surface of the body bright green up to its medial portion, abruptly changing to orange. A marbled black and white stripe borders this green color. Lateral surface of the body includes a thin pale orange stripe upon an areolate skin, ventrally changing to bright orange upon a smooth skin ( Figs. 2 View FIGURE 2 , 3 View FIGURE 3 ). Throat, chest and abdomen pale yellow ( Fig. 2 View FIGURE 2 ).
In forelimbs, the bright green is present on the dorsal surface of the forearm, bordered by a marble black and white stripe that gradually changes to bright orange in the upper arm and forearm, reaching the finger IV. Remaining dorsal and ventral surfaces of hands pale orange; dorsal surfaces of the adhesive discs in fingers brownish-orange ( Figs. 2 View FIGURE 2 , 3 View FIGURE 3 ). Nuptial pad brown. On hindlimbs, bright green covers most of the dorsal surface of tibia and reaches the toe V. The bright green is bordered in feet by a marbled black and white stripe, which reaches the dorsal surfaces of toes IV and V and the adhesive disc of toe V. This marble stripe is gradually replaced by bright orange, which covers the remaining dorsal and ventral surfaces of thighs (along with sparse pale yellow over the areolate skin), ventral surface of tibia, and dorsal surface of feet. Dorsal surfaces of adhesive discs of toes brownish-orange. Ventral surface of feet, toes and adhesive discs of toes pale orange. The calcar retains part of the bright green from dorsal surfaces of tibia, but is marble in most of its dorsal and ventral surfaces ( Figs. 2 View FIGURE 2 , 3 View FIGURE 3 ).
Transparent eyelid, bordered by a thin grayish-white stripe. A black bordered eye with a black pupil, a bicolored iris (golden-yellow in its upper third and light-grey in the remaining lower portion), dark vermiculations and a diffuse dark stripe in the medial region ( Figs. 2 View FIGURE 2 , 3 View FIGURE 3 ). Newly metamorphosed individuals have a similar coloration pattern, except by an olive green color instead of bright green, and by less pronounced grey borders between the olive green and bright orange areas ( Fig. 3 View FIGURE 3 ).
In preservative (based on the collected specimen after 10 years of its preservation), green colored areas became light blue, orange areas (at the limbs and the lateral surface of the body) became almost transparent cream, pale yellow areas (the venter and other parts with areolate skin) became creamy-white, as well as the marble stripe delimiting the division of green and orange colored areas; and the pale grey color at dorsal surfaces of hands and feet became greyish-purple ( Fig. 1 View FIGURE 1 ).
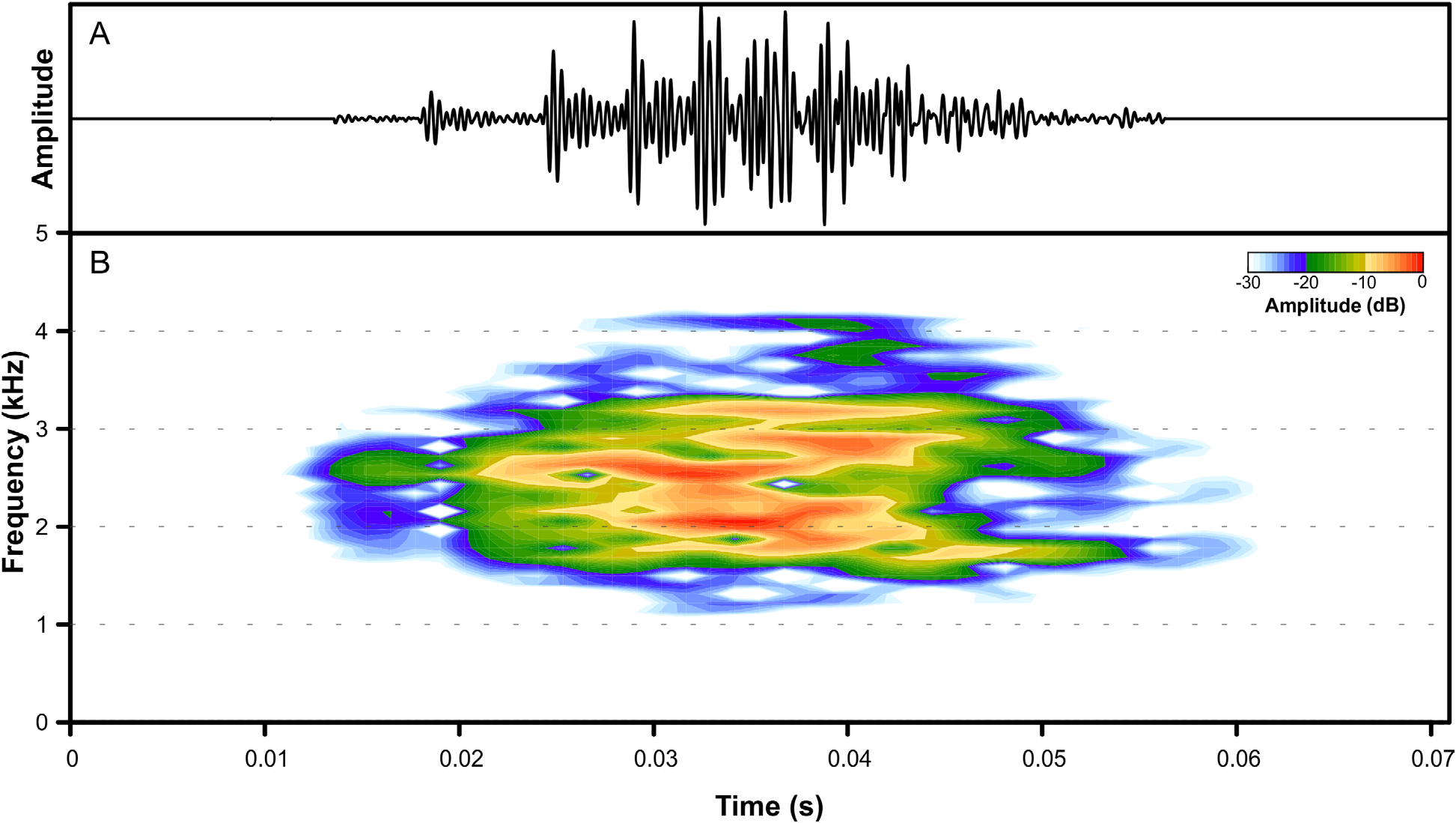
Advertisement call. The call of P. appendiculata in the new locality is composed by a single short note, followed by silence intervals with variable duration ( Fig. 4 View FIGURE 4 ). Call duration is 0.03 ± 0.00 s (n = 15 calls), and is emitted at a variable rate of 2.98 ± 1.66 calls/min (n = 5 males). Notes are composed of highly fused pulses, the first one more evidently segregated from the others ( Fig. 4 View FIGURE 4 ). Minimum frequency ranges 1,236 –1,538 Hz GoogleMaps (1,435 ± 86 Hz; n = 15 calls), peak frequency ranges 1,781 –2,531 Hz GoogleMaps (2,118 ± 301 Hz; n = 15 calls), and maximum frequency ranges 2,824 –3,257 Hz GoogleMaps (3,047 ± 109 Hz; n = 15 calls). The GoogleMaps recorded advertisement call is very similar to the call from Santo André GoogleMaps , state of São Paulo (recorded in 1963) ( Forti et al. 2019). The GoogleMaps only subtle divergences are more evident note pulses, slightly higher maximum frequency, and a lower call rate.
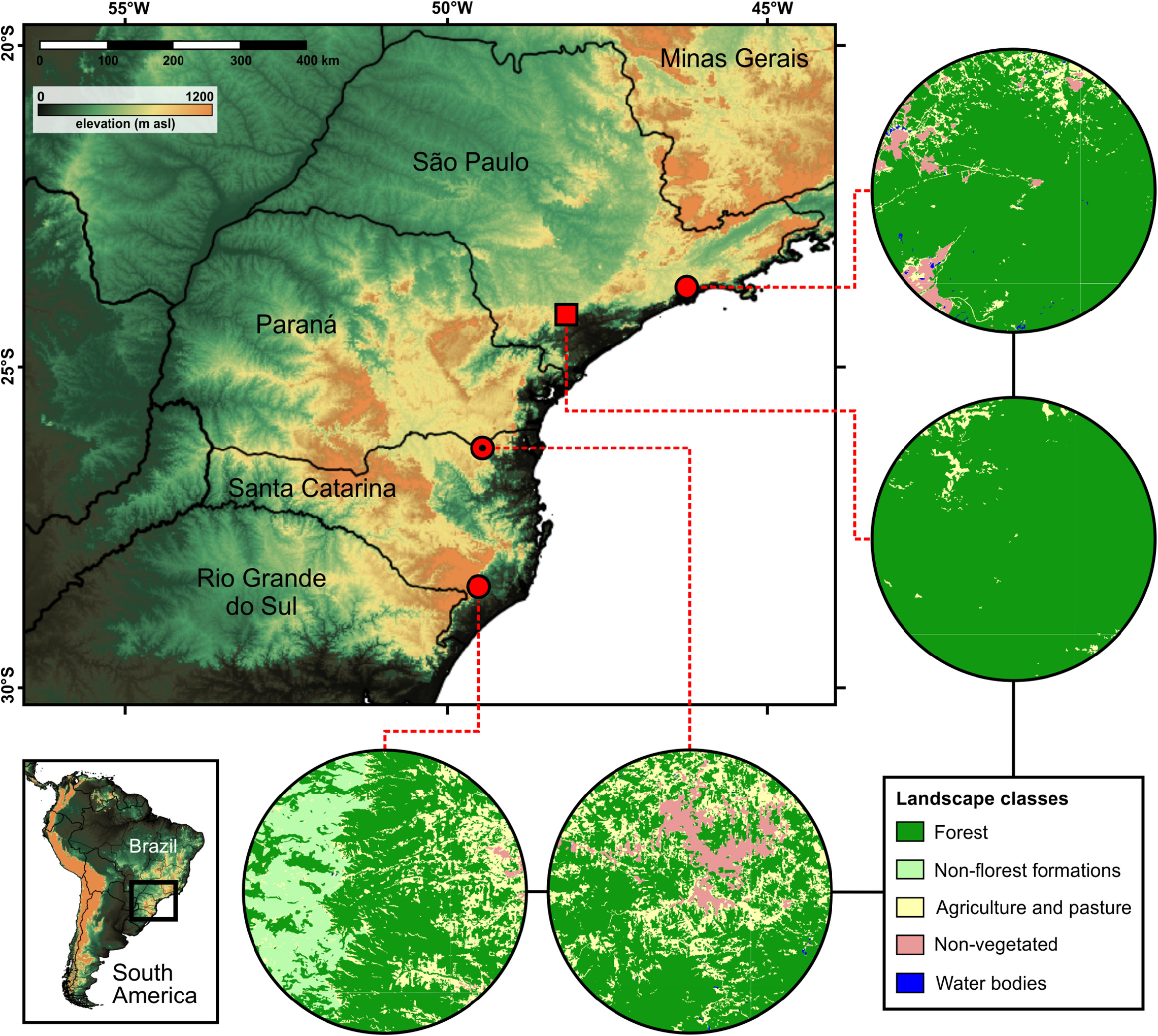
Geographic distribution. Phrynomedusa appendiculata is only known from four localities in montane forests of the Atlantic Plateau at southern Atlantic Forest ( Baêta et al. 2016, Fig. 5 View FIGURE 5 ). These localities are within the limits of the municipalities of São Bento do Sul (type locality) and Lauro Müller in the state of Santa Catarina, and Santo André in the state of São Paulo. Our new record represents the second known locality for the species’ occurrence in the state of São Paulo, also filling a range gap ( Fig. 5 View FIGURE 5 ). This locality is ca. 250 km northwards from the type locality and ca. 200 km southwards from the first record in the state of São Paulo ( Fig. 5 View FIGURE 5 ). It is noteworthy that the species has not been recorded since 1924–1970 in the other three localities, despite being extensively inventoried ( Verdade et al. 2009; Trevine et al. 2014).
Natural history. Despite the diurnal and nocturnal sampling effort in distinct habitats of the regional landscape, we only recorded P. appendiculata during night surveys and in a single collecting site. The species was recorded in this locality ( Figs. 5 View FIGURE 5 , 6 View FIGURE 6 ) during three distinct days, on 04 December 2011 at ca. 22:00 h, on 24 January 2012 at ca. 23:00 h, and on 25 January 2012 at ca. 01:00 h. In late January (rainy season), individuals of P. appendiculata were recorded in breeding activity, based on males emitting advertisement calls and newly metamorphosed individuals ( Fig. 3 View FIGURE 3 ). The species was breeding in two near black-water ponds, part of oxbow lakes of an adjacent river ( Fig. 6B View FIGURE 6 ), corroborating the known habitat condition for breeding events of Phrynomedusa species ( Haddad & Prado 2005; Faivovich et al. 2010, Baêta et al. 2016). About 10 adult individuals of P. appendiculata were assembled at this breeding event based on direct observations and vocalizations. Individuals were spotted perched at leaf petioles, herbaceous vegetation, and thin branches of understory trees, always near or above the water ( Fig. 3A, B View FIGURE 3 , 6C View FIGURE 6 ). No egg-clutch and tadpole were observed. The species was not recorded in lentic rivulets with clean water near these ponds.
Regarding antipredator mechanisms (following the classification system of Ferreira et al. 2019), we observed that P. appendiculata avoid detection through a combination of camouflage (background matching with vegetation), immobility and call interruption. To prevent attack, the species performed a series of mechanisms widely employed by phyllomedusids ( Ferreira et al. 2019): showed hidden orange coloration (supposedly aposematic) in movement; retreated when disturbed, hiding the orange parts of the body (contraction posture); or escaped, by climbing or jumping away (in most cases). Besides the secretory products of cutaneous glands (usual for amphibians; Ferreira et al. 2019), no additional counterattack mechanism were noted. Other syntopic breeding species included Boana bischoffi ( Boulenger, 1887) , Dendropsophus giesleri ( Mertens, 1950) and Physalaemus olfersii ( Lichtenstein & Martens, 1856) .
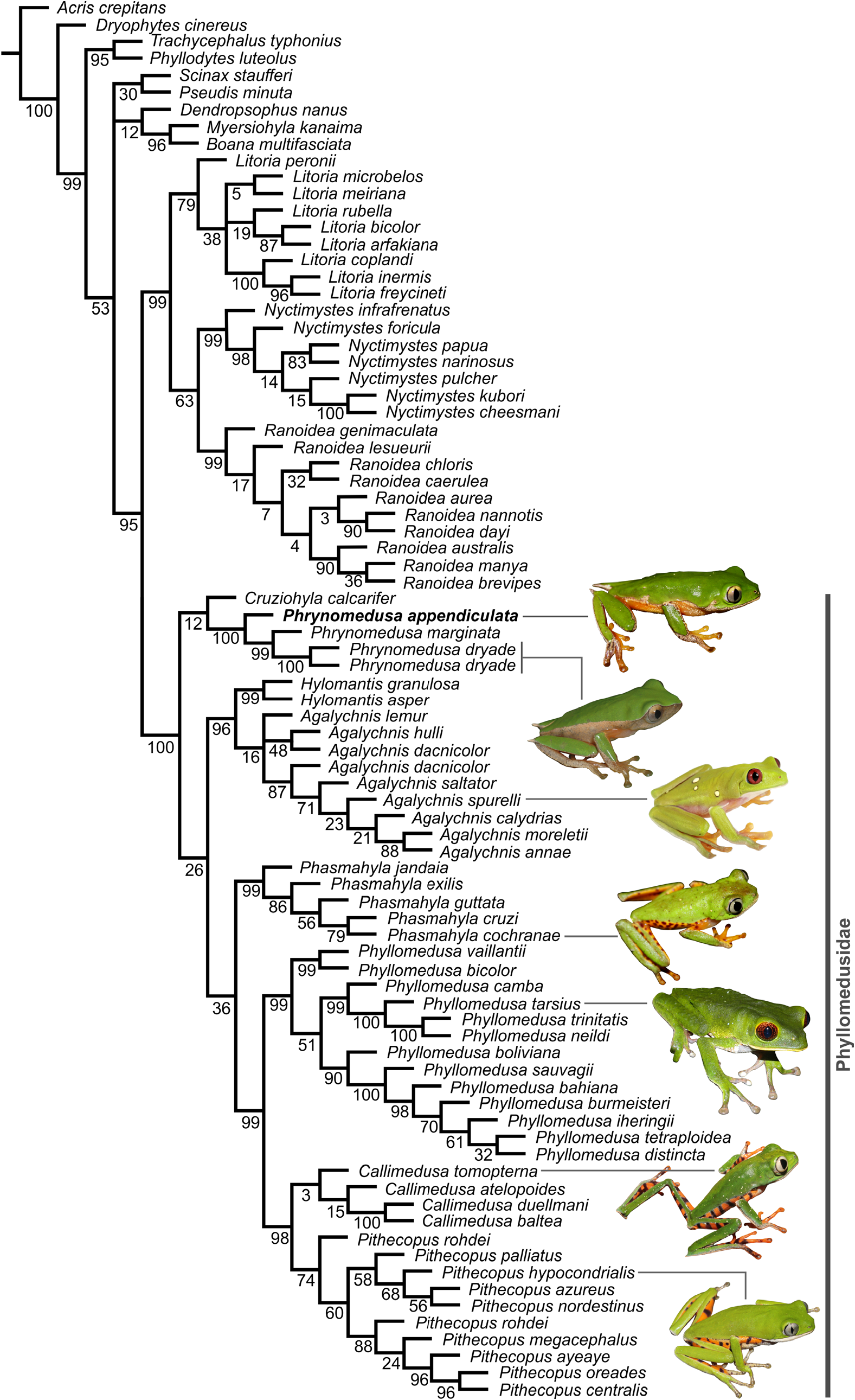
Molecular-based phylogenetic relationships. Our phylogenetic inference mirrored the results of previous phylogenies (e.g. Wiens et al. 2006; Faivovich et al. 2010; Duellman et al. 2016) by evidencing a monophyletic Phrynomedusa as an early divergent lineage within the phyllomedusid radiation ( Fig. 7 View FIGURE 7 ). The generic placement and evolutionary distinctiveness of P. appendiculata was corroborated, as the species is recovered with high support as sister to the clade P. marginata + P. dryade ( Fig. 7 View FIGURE 7 ). Genetic distances between P. appendiculata and P. marginata + P. dryade clade range from 8.9–9.1% (12S), 8.0% (16S), 1.7% (Rag1) and 1.3% (Rhod), while genetic distance between P. marginata and P. dryade reached 3.0% at the single locus sampled for P. marginata (12S).



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.