
Magelona equilamellae Harmelin, 1964
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4767.1.4 |
|
publication LSID |
urn:lsid:zoobank.org:pub:7B5EC00B-44DA-4A09-8B0A-3DCA78CD7C37 |
|
DOI |
https://doi.org/10.5281/zenodo.3799104 |
|
persistent identifier |
https://treatment.plazi.org/id/039487E1-FFDA-1578-FF37-40B51A2B3F64 |
|
treatment provided by |
Carolina |
|
scientific name |
Magelona equilamellae Harmelin, 1964 |
| status |
|
Magelona equilamellae Harmelin, 1964 View in CoL
Figures 1–5 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 , Table 1 View TABLE 1
Material examined. Mediterranean Sea. FRANCE: Rade de Villefranche, opposite the laboratory (see Harmelin 1964): HP6–HP7, ~ 43.696°N, 7.309°E, 13 m, 20 April 1963, degraded Posidonia oceanica meadow covered by a deposit of thin mud, and Golfe de Marseille, off Anse du Prophète, MM 7, ~ 43.272°N, 5.359°E, 18 m, 02 September 1963, “matte morte” of Posidonia oceanica , syntypes ( SMF 4675, 2af, 1f), coll. J.G. Harmelin. SPAIN, CATALO- NIA: Ebro Delta, Alfacs Bay (see Jordana et al. 2015): AT 3–1, 40.605°N, 0.592°E, 5 m, July 2008, mud ( NMW. Z.2019.100.0001, 1af); AT 5–1, 40.629°N, 0.680°E, 4 m, July 2008, muddy sand ( NMW.Z.2019.100.0002, 4af; EJ & SP reference collection, 1af); AT 5–2, 40.629°N, 0.680°E, 4 m, July 2008, muddy sand ( NMW.Z.2019.100.0003, 3af, 2f, 1pf); AT 6–2, 40.633°N, 0.729°E, 1 m, July 2008, medium sand ( SMF No 27727, 2af); AT 7–1, 40.622°N, 0.731°E, 4 m, July 2008, muddy sand ( NMW.Z.2019.100.0004, 1af); AT 7–2, 40.622°N, 0.731°E, 4 m, July 2008, muddy sand ( MUHNAC / MB 29–000344, 2af); AT 9–1, 40.612°N, 0.683°E, 6 m, July 2008, sandy mud ( NMW. Z.2019.100.0005, 2af); AT 9–2, 40.612°N, 0.683°E, 6 m, July 2008, sandy mud ( NMW.Z.2019.100.0006,3af); AT 10–1, 40.602°N, 0.663°E, 2 m, July 2008, fine sand ( NMW.Z.2019.100.0007, 2af);AT 12B–2, 40.592°N, 0.646°E, 2 m, July 2008, fine sand ( NMW.Z.2019.100.0008, 2af); AT 4–1, 40.621°N, 0.631°E, 4 m, July 2011, sandy mud (parcel lost by the postal service, 2af); AT 4–2, 40.621°N, 0.631°E, 4 m, July 2011, sandy mud ( NMW.Z.2019.100.0011, 1af; NMW.Z.2019.100.0012–14, 3af SEM); AT 5–2, 40.631°N, 0.678°E, 2 m, July 2011, muddy sand ( NMW. Z.2019.100.0015, 1af); AT 7–2, 40.622°N, 0.731°E, 4 m, July 2011, muddy sand ( NMW.Z.2019.100.0016, 1af); AT 10–2, 40.602°N, 0.663°E, 2 m, July 2011, medium sand ( NMW.Z.2019.100.0017, 1af); AT 11–2, 40.603°N, 0.626°E, 6 m, July 2011, mud ( NMW.Z.2019.100.0018, 1af). SPAIN, CATALONIA: Ebro Delta, Fangar Bay (see Jordana et al. 2015): AT 15–1, 40.784°N, 0.724°E, 4 m, July 2008, sandy mud ( NMW.Z.2019.100.0009, 1af); AT 15–2, 40.784°N, 0.724°E, 4 m, July 2008, sandy mud ( NMW.Z.2019.100.0010, 1af); AT 14–1, 40.780°N, 0.746°E, 4 m, July 2011, sandy mud ( NMW.Z.2019.100.0019, 2af; EJ & SP reference collection, 1af). SPAIN, CATALONIA: Ebro Delta, Alfacs Bay (see Capaccioni-Azzati 1987; 1989): E–3 AH, ~ 40.595°N, 0.615°E, 2 m, September 1984, sandy mud ( MNCN 16.01/18575, 2af); I–1 AH, ~ 40.632°N, 0.708°E, 3 m, April 1985, muddy sand ( MNCN 16.01/18576, 2af, 1f).
Diagnosis. Stout species, with thoracic pigment band. Prostomium wider than long, subtriangular carrying one pair of ridges, but without prostomial horns. Chaetigers 1–9 with slender, smooth-edged, triangular lamellae, neuropodial lamellae initially prechaetal, becoming postchaetal by posterior thorax. Thoracic chaetigers with capillary chaetae only. Abdominal lateral lamellae long, pointed triangular lacking dorsal or ventral processes. Hooded hooks tridentate, in two groups, vis-à-vis.
Dimensions. Larger syntype; prostomium 0.85 mm long, 1.4 mm wide; thorax (including prostomium) 5.2 mm long, 1.4 mm maximum width (excluding lamellae); abdomen 1.25 mm wide; total length 12.0 mm for 19 chaetigers. Smaller syntype; prostomium 0.55 mm long, 1.1 mm wide; thorax (including prostomium) 5.0 mm long, 1.0 mm maximum width (widest at chaetiger 1, tapering towards abdomen); abdomen 0.85 mm wide; total length 7.0 mm for 14 chaetigers. 4 chaetiger fragment, of similar width to larger syntype present. Other non-type specimens with 9–29 chaetigers for 2.5–14 mm in length.
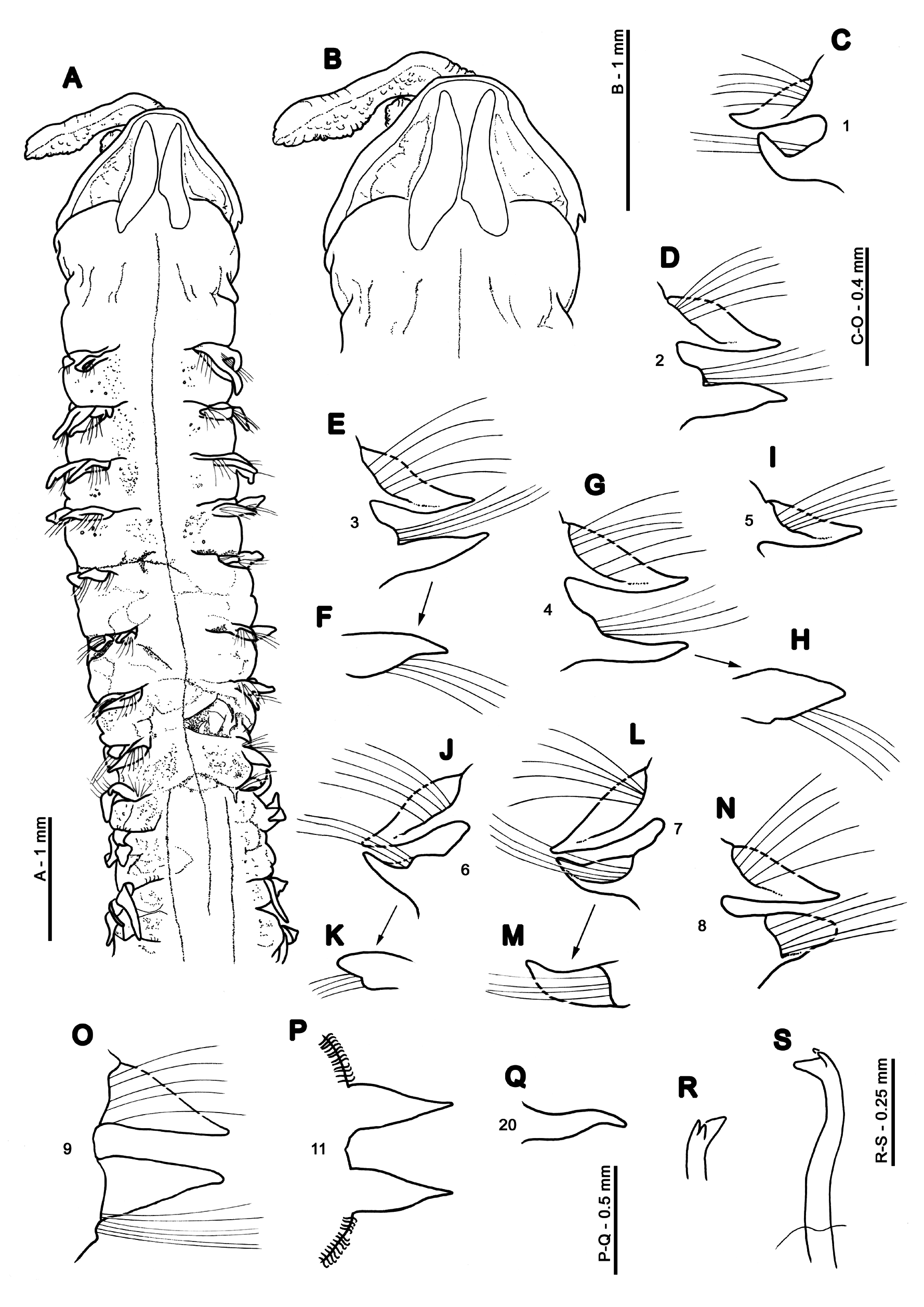
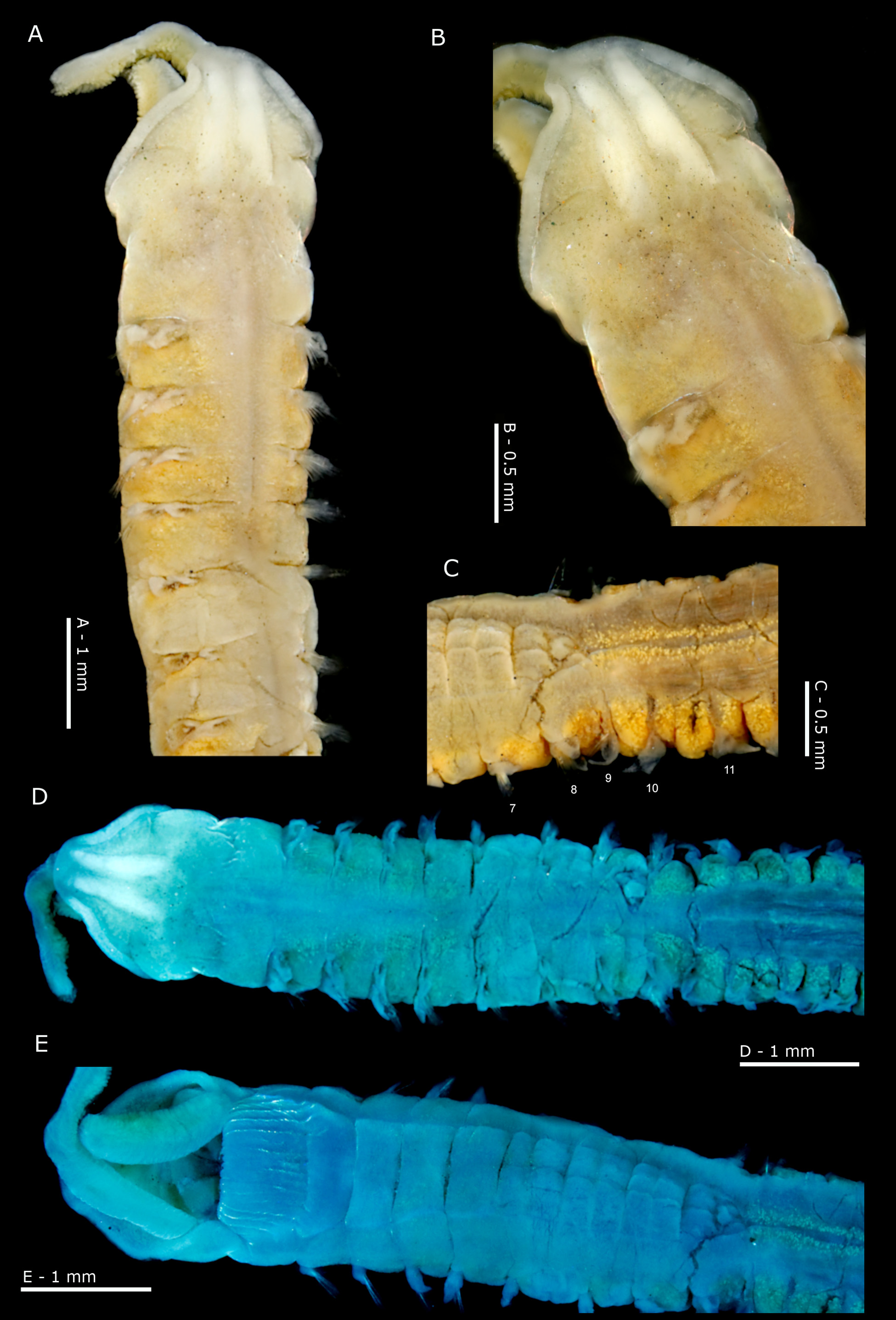
Description. A large, stout species ( Figs 1A View FIGURE 1 ; 2 View FIGURE 2 ; 3 View FIGURE 3 ; 4A View FIGURE 4 ; 5A View FIGURE 5 ); thorax marginally wider (when viewed dorsally), but slightly thinner (when viewed laterally) than abdomen, constriction at chaetiger 9 not distinct. Both syntypes posteriorly incomplete, and lamellae (particularly those of the thoracic region) showing signs of damage (e.g. neuropodia of chaetiger 5 of larger syntype). Prostomium wider than long ( Figs 1B View FIGURE 1 ; 2B View FIGURE 2 ) (L:W ratio 0.5–0.61), subtriangular, anterior margin smooth. Prostomial horns absent, although anterior margin squared. Lateral prostomial margins of several specimens, including larger syntype, relatively thick compared to those on anterior margin ( Figs 1B View FIGURE 1 ; 2B, D View FIGURE 2 ; 3A View FIGURE 3 ), although, of a similar size in smaller syntype. One pair of longitudinal dorsal muscular ridges abutting medially but divergent at either end ( Fig. 1A, B View FIGURE 1 ). Very faint markings either side of prostomial ridges, indistinct ( Fig. 4B View FIGURE 4 ). Bur- rowing organ not everted in type material but everted partially in 11 specimens from Ebro Delta, longitudinally ridged, somewhat fainter on superior surface. Very short palps (broken) on larger syntype, arising ventrolaterally from base of prostomium ( Fig. 2E View FIGURE 2 ), heavily papillated. Palps retained on 12 specimens from Ebro Delta ( Figs 3C, D View FIGURE 3 ; 4 View FIGURE 4 A–C). Palps generally short (reaching up to chaetiger 20 when folded backwards, length between 4.5–10 mm) and thick ( Fig. 4A View FIGURE 4 ). Non-papillated region short, reaching chaetigers 1–2 when folded backwards ( Fig. 4B View FIGURE 4 ). Papillae very long, giving palps a ‘frilly’ appearance ( Fig. 4A View FIGURE 4 ); 6–8 rows of papillae proximally and medially, 2–4 rows distally. Papillae grouped equally either side of inconspicuous mid palp line, devoid of papillae, e.g. 1–2 papillae in each row either side distally, and 3–4 medially and proximally. Buccal region on ventral side of prostomium with three lips, one larger triangular lip above two smaller lateral lips ( Fig. 3D View FIGURE 3 ). Buccal region appearing large and swollen in several specimens.
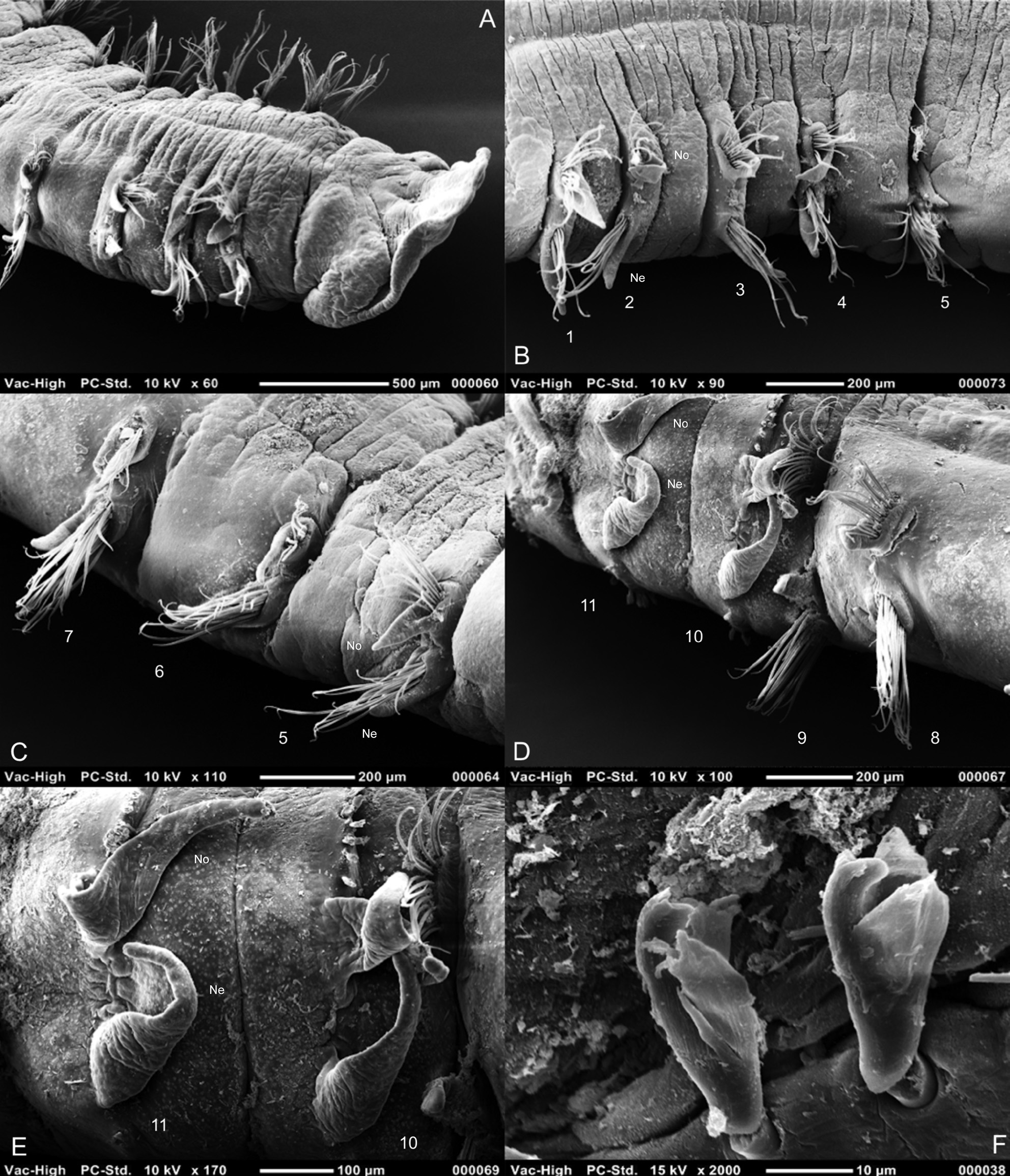
Achaetous region behind prostomium large, approximately twice the size of chaetiger 1 ( Figs 1A View FIGURE 1 ; 2A, B View FIGURE 2 ). Chaetigers 1–8 ( Figs 1 View FIGURE 1 C–N; 5A–D) similar; parapodia biramous, low triangular notopodial prechaetal ridges confluent with slender triangular postchaetal lamellae, with pointed tips. Join between pre- and postchaetal lamellae towards the distal tip of the postchaetal lamellae, the latter of which becomes almost sub-chaetal by posterior thorax. No superior dorsal lobes present on thorax (N.B. where pre- and postchaetal lamellae meet, above chaetal bundle, may be hunched, resembling a process, see Fig. 5B View FIGURE 5 , chaetiger 4). Neuropodial postchaetal lamellae triangular, initially prechaetal in position ( Figs 1 View FIGURE 1 C–E), moving to a ventral position (underneath chaetal bundle, Figs 1G, J View FIGURE 1 ) and becoming entirely postchaetal from chaetiger 7. Neuropodial lamellae of fresher material often slightly longer, with more pointed, delicate tips ( Figs 3A View FIGURE 3 ; 5A, B View FIGURE 5 ). However, within fresher material, lamellae of a few parapodia with broken tips resembling that of syntypes.
Parapodia of chaetiger 9 ( Figs 1O View FIGURE 1 ; 5D View FIGURE 5 ): notopodia similar to preceding chaetigers, however, neuropodial lamellae more triangular, in a distinctly lateral position, with low inferior postchaetal ridges terminating in minute triangular processes (not seen in smaller material and sometimes observed only on one side of a chaetiger). Prechaetal lamellae low ridges in both rami. Parapodia of chaetiger 9 often interposed in furrow between surrounding chaetigers, particularly neuropodial ( Fig. 5D View FIGURE 5 ). All thoracic chaetae bilimbate capillaries, those towards posterior thorax with wider limbations with an irregular blade. Neurochaetae longer than the notochaetae. Chaetae of chaetiger nine splayed apart in both rami. No ventral swellings or markings of the thoracic region observed in any specimens.
Abdominal chaetigers with sharply-pointed triangular lateral lamellae, of about equal size in both rami ( Figs 1P, Q View FIGURE 1 ; 5D, E View FIGURE 5 ), those of the anterior abdomen longer than thoracic lamellae. Lateral lamellae do not extend postchaetally, hooks rise from definite ridge. No dorsal (DML) or ventral (VML) processes observed on abdominal chaetigers. Larger syntype damaged abdominally between chaetigers 14–17.
Abdominal chaetae all tridentate hooded hooks ( Figs 1R, S View FIGURE 1 ; 5F View FIGURE 5 ), of similar size (no small hooks adjacent to lamellae observed). Hooks in two unequal groups (group adjacent to lamellae with fewer hooks, therefore can look unidirectional if these few are broken), main fangs vis-à-vis. Hooks in middle of rami often occurring in two rows side by side. Initially about 14 hooks per ramus (syntype), reducing to 8–10 on median chaetigers, however, many hooks on larger syntype broken. No pouches observed. Syntypes both posteriorly incomplete, however, one pygidial fragment present within Catalonian material (NMW.Z.2019.100.0003, N.B. although unattached, as this was the only magelonid species within the sample it is believed to belong to M. equilamellae ). No anal cirri observed, anus large and terminal but tilted towards ventral surface ( Fig. 4D View FIGURE 4 ).
Tube. Several specimens with distinct paper-like tubes covered in sediment were observed (NMW. Z.2019.100.0001; NMW.Z.2019.100.0005; NMW.Z.2019.100.0006, Fig. 4E View FIGURE 4 ).
Colour. Only preserved material observed; syntypes now dark cream, pigment band faded. Pigment band retained on fresher material ( Fig. 3A View FIGURE 3 ) between chaetigers 4/5–8 as noted for the syntypes. Glandular areas particularly noticeable in abdomen as broad, intensely speckled interparapodial patches (often starting from chaetiger 8, Fig. 2C View FIGURE 2 ). Dorsal speckled areas just behind the parapodia, either side of the mid thoracic line noticeable from chaetigers 1–4 and in the posterior thorax, extending laterally ( Figs 3A, C View FIGURE 3 ). Very sparsely speckled areas dorsally in pigment band region, ventrally on chaetigers 2–3 and along mid ventral line. Methyl green staining rather diffuse ( Figs 2D, E View FIGURE 2 ), no obvious pattern. Slightly darker staining in mid-dorsal (chaetigers 4 to 7) and mid-ventral areas of thorax. Staining increasing visibility of speckled areas next to parapodia, in particular between chaetigers 1–4. Several specimens stained with Rose Bengal ( Fig. 4 View FIGURE 4 ), colouration generally light all over; speckled areas slightly pigmented. Buccal region heavily speckled on inner surfaces of the mouth and down into the gut. The latter staining is visible through the body wall level with chaetigers 1–2.
Habitat. Harmelin (1964) collected M. equilamellae in silty sediments and in amongst “ matte morte ” (leaf frag- ments and root-rhizomes of the Mediterranean seagrass Posidonia oceanica , see Borg et al. 2006 for full description) at relatively shallow depths (13 and 18 m respectively). Further records exist from muddy, silty sediments, amongst detritus and with the following species: Venus ovata, Nucula sulcata, Abra alba , Amphiura chiajei, Scoloplo s armiger, Caulerpa prolifera and Cymodocea nodosa ( Guille 1971; 1972; 1973; Capaccioni-Azzati 1983; 1987; 1989). The species has been recorded to occur in lagoons, estuarine inlets ( Capaccioni-Azzati 1983; 1987; 1989; Capaccioni-Azzati et al. 1987; 1991), semi-enclosed shallow water bays ( Martin et al. 2000) and from one locality “under the influence of a sewage outfall” ( Royo-García et al. 2009). Capaccioni-Azzati (1989) suggested the species is euryhaline, occurring in both hyperhaline and mixo-euhaline waters, something not commonly reported for magelonid species. Present observations indicate that M. equilamellae may be typical of transitional waters. Depths between 0–50 m have been recorded for the species ( Aguirrezabalaga et al. 1988; Dando et al. 1995; Martin et al. 2000; Royo-García et al. 2009; Çinar et al. 2014; Koulouri et al. 2015).
In the particular case of the Ebro Delta, Catalonia, specimens recorded herein were collected in sediments with a high silt-clay (at least 10% up to 79%) and organic matter (up to 11%) content, corresponding to muds, sandy muds and muddy sands. However, they were also present in predominantly sand sediments, without a silt-clay fraction (0%) and with an organic matter content below 1%. All observations were from shallow waters (between 1 and 6 m) and in many cases with the presence of Caulerpa prolifera and Cymodocea nodosa . The bays of the Ebro Delta, where the highest number of animals were recorded, are semi-enclosed, transitional waters ( Jordana et al. 2015; 2016) highly influenced by continental waters with little water renewal. Thus, supporting the view of Capaccioni- Azzati (1989) that M. equilamellae is a euryhaline species.
The results of the present study (2008 and 2011) have revealed several locations in Els Alfacs Bay where M. equilamellae was not recorded during 1984–1985 surveys (Capaccioni-Azzati 1987; 1989). These sites are located: in the entrance of the bay (AT 3, muddy sediments inhabited by a Amphiura chiajei community), the innermost area of the bay (AT 7, corresponding to muddy sand sediments) and the southern cove area (AT 10 and AT 12B, consisting of clean sands with less than 1% of organic matter and no mud, see Jordana et al. 2015). It should be noted this is the first record of this species occurring in fine or medium sands sediments of non-modified waters ( Jordana et al. 2016).
Within Alfacs Bay and Fangar Bay M. equilamellae is found to co-occur with the morphologically similar M. alleni at several stations (Ebro Delta: AT 3–1, AT 9–1, AT 9–2 of July 2008 and AT 9–1, AT 11–2, AT 14–1 of July 2011).
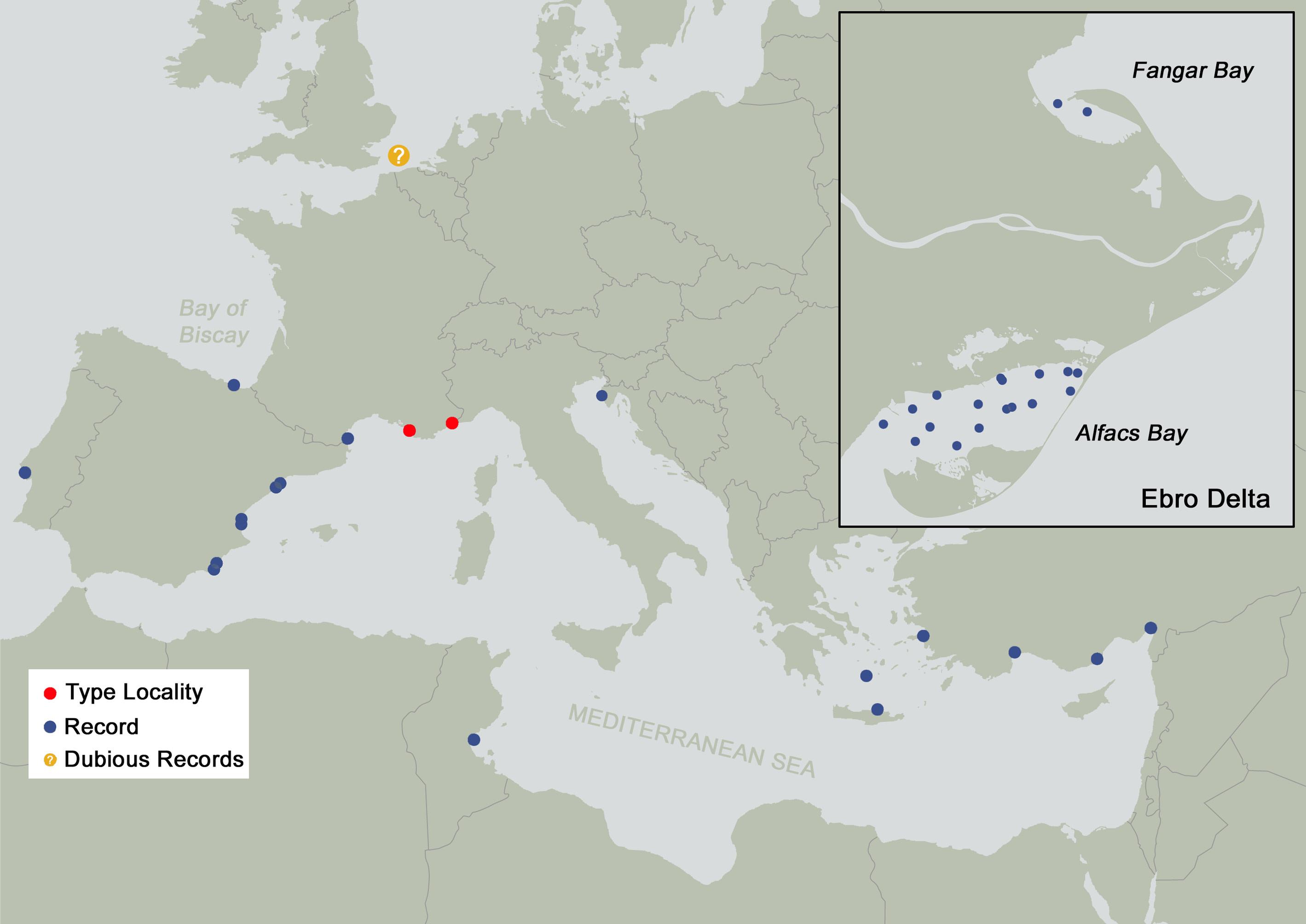
Distribution. Magelona equilamellae is recorded from several localities around the Mediterranean ( Fig. 6 View FIGURE 6 ): Southern France ( Harmelin 1964; Guille 1971; 1972; 1973); East Mediterranean Coast of Spain ( Capaccioni-Azzati 1983; 1987; 1989; Capaccioni-Azzati et al. 1987; 1991; Tena et al. 1993; Martin et al. 2000; Royo-García et al. 2009; Martínez-Garcia 2019); Adriatic Sea ( Amoureux 1976; Cantone 2003; Mikac 2015); Greece ( Dando et al. 1995; Arvanitidis 2000; Faulwetter 2010; Koulouri et al. 2015); Turkey ( Çinar et al. 2014, plus records from the Museum of Ege University Faculty of Fisheries: ESFM-POL/2005–238, 1326, 1426, 1436); and lastly Tunisia ( De Gaillande 1970; Ayari et al. 2009). Fiege et al. (2000) considered the species to be limited to the Mediterranean. However, it has been additionally recorded outside of this region in the Bay of Biscay ( Aguirrezabalaga et al. 1988) and Tagus Estuary, Portugal ( Sousa 2016). Mills & Mortimer (2019b) concluded that all northern European records (North Sea, off Wales and Sweden: Hobbs et al. 1997; Hansson 1998; English et al. 2008; Cooper & Barry 2017; Flanders Marine Institute 2019) of the species are unlikely/incorrect and that records in Southern and Western European waters outside the Mediterranean warrant verification given previous misidentifications with M. alleni (see remarks below) and M. filiformis . For the latter species, misidentifications associated with the key in Fiege et al. (2000) have occurred due to misunderstandings about the presence/absence of frontal horns (the squared anterior margin of M. filiformis being often termed “rudimentary horns”). As both species possess equal abdominal lamellae, inevitably a M. filiformis specimen will be keyed out as M. equilamellae if an identifier regards the species as being without frontal horns.
Diet. One specimen (NMW.Z.2019.100.0003) with balls of sediment clearly visible within body cavity as seen previously for M. alleni ( Mills & Mortimer 2019a) .
Remarks. The original description of M. equilamellae by Harmelin contained drawings of the lamellae of only one thoracic and one abdominal chaetiger. However, since its description, the importance of describing and drawing the morphology of every thoracic chaetiger has been highlighted (e.g. Brasil 2003), including the shape and size of lamellae and their position in comparison to chaetal bundles. Several features not originally described, particularly in relation to the thoracic region (e.g. neuropodial lamellae being distinctly prechaetal in the anterior thorax but postchaetal by the posterior thorax), are noted herein.
The material from Ebro Delta, Catalonia, agrees well with the syntype material in all respects. Nevertheless, in some specimens there were slight differences noted in terms of the length and pointedness of lamellar tips between the type and fresher material. However, it was observed in some of the fresher material, where odd lamellar tips had been broken, that their shape matched that of the type material. As noted above the lamellae of the syntype material are not well preserved and those of the fresher material are particularly delicate, thus suggesting that the syntypes are likely to have had slightly longer and more pointed lamellar tips originally. Certainly, it should be noted that a couple of thoracic lamellae on the smaller syntype were more pointed, giving credence to this idea.
Harmelin (1964) stated that based on the presence of a thoracic pigment band and tridentate abdominal hooks, but in lacking specialised thoracic chaetae, prostomial horns, and abdominal medial lobes, M. equilamellae shares similarities with two other magelonid species: M. alleni and Magelona cincta Ehlers, 1908 . Whilst he stated that it differed from the latter species in the shape of the prostomium and the development of the lateral lamellae, it further differs in lacking scoop shaped neuropodial lamellae in the anterior thorax, and in the lamellae of chaetiger 9, which are triangular (those of M. cincta are digitiform and in distinctly lateral positions both in the noto- and neuropodia, see Mortimer & Mackie 2009: fig. 5K). Although, M. cincta is a South African species, both M. equilamellae and M. alleni occur in European waters, co-occurring within the same habitats in some regions (see above). They differ in the nature of the abdominal lamellae; being sub-equal in the latter species and somewhat equal in the former species. Despite this difference, the two species share many morphological similarities. In fact, this has led to much confusion between the two species, as was previously noted by Fiege et al. (2000). However, there are several additional features that allow their separation, even in the absence of abdominal chaetigers. The prostomium of M. alleni is narrower at the distal end, whilst in M. equilamellae the lateral margins are much thicker than the anterior margin, and in comparison, to the lateral prostomial margins of M. alleni . The change from a prechaetal to postchaetal position of the thoracic neuropodial lamellae along the thorax of M. equilamellae is in contrast to those of M. alleni , which are ventral, underneath the chaetal bundle.
There are two additional species known to carry distinct thoracic pigment bands: Magelona japonica Okuda, 1937 and Magelona variolamellata Bolívar & Lana, 1986 . However, M. equilamellae differs from the former species in lacking prostomial horns (present in M. japonica ). Whilst M. equilamellae shares many similarities with M. variolamellata in terms of prostomial and lamellar shape, it differs in the nature of the lamellae of chaetiger 9, in lacking dorsal or ventral processes on abdominal chaetigers, and lacking thoracic dorsal furrows (both present in M. variolamellata ).


| MM |
University of Montpellier |
| SMF |
Forschungsinstitut und Natur-Museum Senckenberg |
| NMW |
Naturhistorisches Museum, Wien |
| EJ |
Ein Yabrud collection catalogue entries at The Hebrew University |
| SP |
Instituto de Botânica |
| MUHNAC |
Museu Nacional de Historia Natural e de CIência |
| MB |
Universidade de Lisboa, Museu Bocage |
| MNCN |
Museo Nacional de Ciencias Naturales |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.