
Apostenus californicus, Ubick, Darrell & Vetter, Richard S., 2005
|
publication ID |
https://doi.org/ 10.5281/zenodo.801450 |
|
persistent identifier |
https://treatment.plazi.org/id/0D2EDC50-6D44-FFEC-27A9-F97FFC7966F1 |
|
treatment provided by |
Jeremy |
|
scientific name |
Apostenus californicus |
| status |
sp. nov. |
Apostenus californicus View in CoL new species
Type material.- Male holotype GoogleMaps and female allotype from moist Quercus kelloggi duff at intersection of Cedar Springs and Pacific Crest Trails off Morris Ranch Road , 33º40'00"N, 116º34'31"W, 2090 m, San Jacinto Mountains, Riverside County, California, U.S.A., 7 January 2001, R. Vetter ( CAS) GoogleMaps .
Paratypes: U.S.A.: California: Kern County: 1 ♂, 1juvenile, Los Padres National Forest, 100 m S of snow gate on Cuddy Valley Road toward Mount Pinos, (at mile marker 6.01), 1 km S of intersection with Cerro Mil Potrero Hwy , 34º49'51"N, 119°05'03"W, 1895 m, in moist Quercus shrub duff, 12 April 2003, R. Vetter ( UCR) GoogleMaps ; 3 juveniles, same road as above but at mile marker 8.95, 34"49'17"N, 119º05'01"W, 2105 m, in moist Quercus kelloggi duff, 12 April 2003, R. Vetter ( UCR) ; Riverside County: 1 juvenile, San Jacinto Mountains: same locality as holotype, 29 March 2001 GoogleMaps ; 4 juveniles (3 ♀ reared to maturity), 28 April 2001 GoogleMaps ; 1 juvenile, along Cedar Springs Trail (Trail 4E17) , 1950 m, in dry Quercus wislizenii duff, 7 January 2001 GoogleMaps ; 6 ♂, 6 ♀, 9 juveniles (1 ♀ reared to maturity), on Cedar Springs Trail off Morris Ranch Rd. , 33º39'42"N, 116º34'41"W, 1790 m, in moist Quercus chrysolepis duff, 30 September 2001 GoogleMaps ; 4 females, 4juveniles, near Cedar Springs trailhead, 33º39'26"N, 116º35'01"W, 1720 m, in dry Quercus chrysolepis oak duff near streambed, 7 January 2001 GoogleMaps ; 1juvenile, in moist Quercus chrysolepis oak duff under snow, 18 March 2001; all above collected by R. Vetter GoogleMaps (6 ♂ and 4 ♀ at CAS, remainder at UCR); 2 ♂, 2 ♀, 1 juvenile, James Reserve, Lake Fulmor , 33º48'31"N, 116º46'36"W, 1640 m, in dry Quercus kelloggi oak-pine duff next to wet stream, 8 October 200 1, R. Vetter and T. Prentice ( UCR) GoogleMaps ; 1 ♀, 4.2 km N Lake Fulmor on Hwy 243, trailhead of trail 2E35 , 33º49'39"N, 116º47'44"W, 1575 m, in extremely dry Quercus leaf duff, 26 September 2003, R. Vetter ( UCR) GoogleMaps ; 1 ♀, 1juvenile, Spillway Canyon, S of Lake Hemet , 33º39'07"N, 116º41'32"W, 1365 m, probably from oak litter, 29 May 2001, T. Prentice & D. Popko ( UCR) GoogleMaps ; San Bernandino County: San Bernardino Mountains: 1 ♀, 4.8 km W Angelus Oaks general store on Hwy 38 , 34º10'N, 116º52'W, 1820 m, in Quercus kelloggi duff, 6 June 2003, R. Vetter ( UCR) GoogleMaps ; 1 ♀, Forest Falls, Momyer-Alger Trail , 34º05'05"N, 116°55'07"W, 1660 m, 1April 2001, TR. Prentice ( TRP) GoogleMaps ; 6 juveniles (1 ♂, 2♀ reared to maturity), in oak duff, 28 May 2001, T. Prentice ( UCR) GoogleMaps ; 1juvenile (♀ reared to maturity), 17 April 2002, T Prentice ( TRP) GoogleMaps ; 1♀, 2 juveniles, Forest Falls, near Vivian Creek trailhead (Trail 1E08) , 34º04'58"N, 116°53'35"W, 1850 m, in dry scrub oak duff, 25 March 2001, R. Vetter ( UCR) GoogleMaps ; 2 ♀, 3 juveniles, 1 April 2001, T. Prentice ( TRP) GoogleMaps ; 1 juvenile, Forsee Creek and Hwy 38, 0.4 mi E of Camp Cedar Falls turnoff , 34º09'29"N, 116º55'54"W, 1850 m, in Quercus sp. duff, 15June 2003, R. Vetter ( UCR) GoogleMaps ; 1 ♀, 1 km W Jenks Lake Loop Road East turnoff, 34º10'14"N, 116º50'29"W, 2093 m, in scrub oak duff, 6 May 2001, R. Vetter ( UCR) GoogleMaps ; 1 penultimate male, Ponderosa Pines trail (1E19)near w entrance to Jenks Lake Loop Road on Hwy 38 , 34º09'56"N, 116°54'46"W, 1950 m, in Quercus sp. duff, 15 June 2003, R. Vetter ( UCR) GoogleMaps ; 3♀, 6 juveniles, Mill Creek Canyon, 1.3 km E of Hwy 38 on Valley of the Falls Dr ., 34º05'42"N, 116°56'44"W, 1450 m, 2 March 2002, R. Vetter ( UCR) GoogleMaps ; 1juvenile, near Seven Oaks, 1.6 km N of Hwy 38 on Glass Rd , 34º10'29"N, 116"54'00"W, 1820 m, in mixed Quercus kelloggi and pine duff, 6 May 2001, R. Vetter ( UCR) ; 1 ♀, 1 juvenile, in Quercus kelloggi and Q. chrysolepis duff, 6 June 2003, R. Vetter ( UCR) ; 1 ♀, 3 juveniles, Skinner Ridge between Skinner Creek and Mountain Home Creek , 34º06'48"N, 116º58'53"W, 1500 m, in oak duff, 29 November 1983 - 26 January 1984, M. Narog ( UCR) GoogleMaps ; 1 ♂, 23 January 1986, M. Narog ( UCR) GoogleMaps ; San Diego County: 1♂, 5♀, 1 penultimate ♂, 3 juveniles, Cleveland National Forest, Julian, 4839 Pine Ridge Ave ., 33º02'34"N, 116º37'49"W, 1300 m, in mixed Quercus kelloggi and Quercus sp. leaf duff, 31 March 2002, R. Vetter ( UCR) GoogleMaps ; 2 penultimate ♂, Cleveland National Forest, ca. 1.6 km N Cibbets Flat , 32º46'38"N, 116º26'56"W, 1250 m, 12 July 2003, J. Berrian ( SDM) GoogleMaps ; 1 ♂, 1♀, Descanso Junction , 32º50'N, 116º36'W, 1040 m, ex willow duff, 31 March 1961, E. Lindquist ( CDU) GoogleMaps ; 4 juveniles, Laguna Mountain across from fire station, 1/ 8 mi N Camp Ole Station , 32º53'N, 116º25'W, 1755 m, in duff of black oak, Quercus kelloggi, 20 February 2003, L. Merrill and R. Vetter ( UCR) GoogleMaps ; 2♀, Palomar Mountain State Park, Doane Pond trail, 20 m from parking lot , 33°20'29"N, 116°54’05”W ,, 14 15 m, in mixed Quercus oak duff on creek bank next to road, 20 January 2003, R. Vetter ( UCR) GoogleMaps ; 1 ♀, 0.3 mi W of Ranchita on Hwy S22 , 33º12'37"N, 116º32'30"W, 1193 m, in oak leaf duff, 16 March 2003, R. Vetter ( UCR). GoogleMaps
Non-para types (identification tentative): U. S.A.: California: 2 juveniles, Inyo County: Independence: Oak Creek Campground, just beyond Mt. Whitney Fish Hatchery , 36º50'31"N, 118º15'37"W, 1455 m, in black oak duff, 7 May 2003, E.E Drake ( UCR) GoogleMaps .
Etymology.-The species name refers to its known distribution.
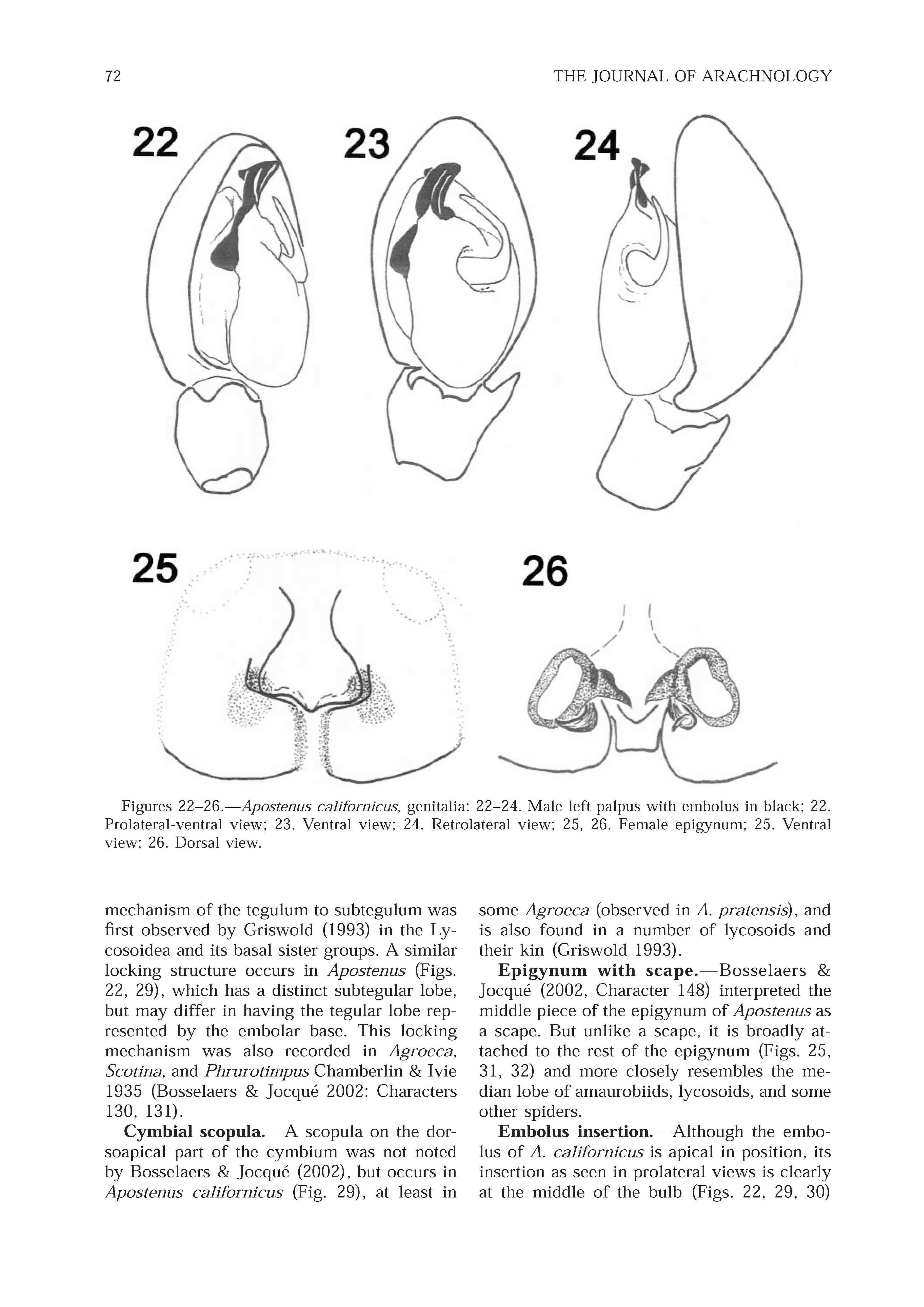
Diagnosis.-This is the only Apostenus known from the Nearctic region. The male is similar to A. annulipedes Wunderlich from which it differs in having the median apophysis longer and originating more basad on the bulb. The female is close to A. grancanariensis Wunderlich (male unknown) but has the spermathecae more widely separated. (compare Figs. 22-26 View Figures 22 - 26 with figs. 750e-h and 75 1- 751a in Wunderlich 1992).
Description.- Male (holotype, range o f other males in parentheses; n = 8): Total length 2.42 (2.24-2.95). Carapace length 1.10 (0.98-1.15), width 0.88 (0.79-0.94), height 0.34. Clypeus 0.08 (at AME), 0.05 (at ALE). Fovea length 0.18. Abdomen length 1.32, width 0.74. Eye sizes and interdistances: AME 0.04, ALE 0.08, PME 0.06, PLE 0.06, AME-AME 0.03, AME-ALE 0.02, PME-PME 0.06, PME-PLE 0.04, ALE-PLE 0.04, AER 0.25, PER 0.30. Palpus and leg lengths: Palpus: 1.10 (0.38 + 0.34 + 0 + 0.38); Leg I: 3.38 (0.90 + 1.22 + 0.64 + 0.62); Leg 11: 2.96 (0.84 + 1.02 + 0.58 + 0.52); Leg 111: 2.84 (0.76 + 0.94 + 0.64 + 0.50); Leg I V 4.06 (1.06 + 1.34 + 0.98 + 0.68). Leg formula 4123.
Color: Carapace brown, black in eye region and along margin, light brown at fovea. Abdomen dorsum dark brown to black with two short longitudinal pale marks anteriorly, followed by two pairs of transverse marks, and 2-3 transverse bands posteriorly; venter light brown with dark median maculation. Legs light brown with dark annulations, anterior femora and tibia dark brown, coxae light brown. Sternum brown.
Vestiture: Carapace largely glabrous, eye region and clypeus with strong setae and recumbent white scales in longitudinal band. Sternum with setae mostly at margins and at posterior projection. Abdomen dorsum densely setose, anteriorly with recumbent white setae, venter with modified short setae ( Figs. 15, 16 View Figures 15 - 17 ); appendages densely clothed with long setae, spines, plumose hairs and trichobothria.
Carapace piriform in dorsal view, somewhat flattened, highest at fovea. AME smallest, about half the diameter of ALE, PE subequal slightly smaller than ALE, AER straight, PER slightly recurved in dorsal view. Chelicerae not geniculate, lacking boss, anterior face with several setae, no spines, retromargin with 2 teeth, promargin with 3 teeth. Sternum rounded, anteriorly truncate, with posterior extension between coxae IV, with marginal setae, especially at posterior extension. Precoxal triangles absent. Labium round- ed, wider than long, one half length of endites; endites quadrate, with serrula on anterior margin.
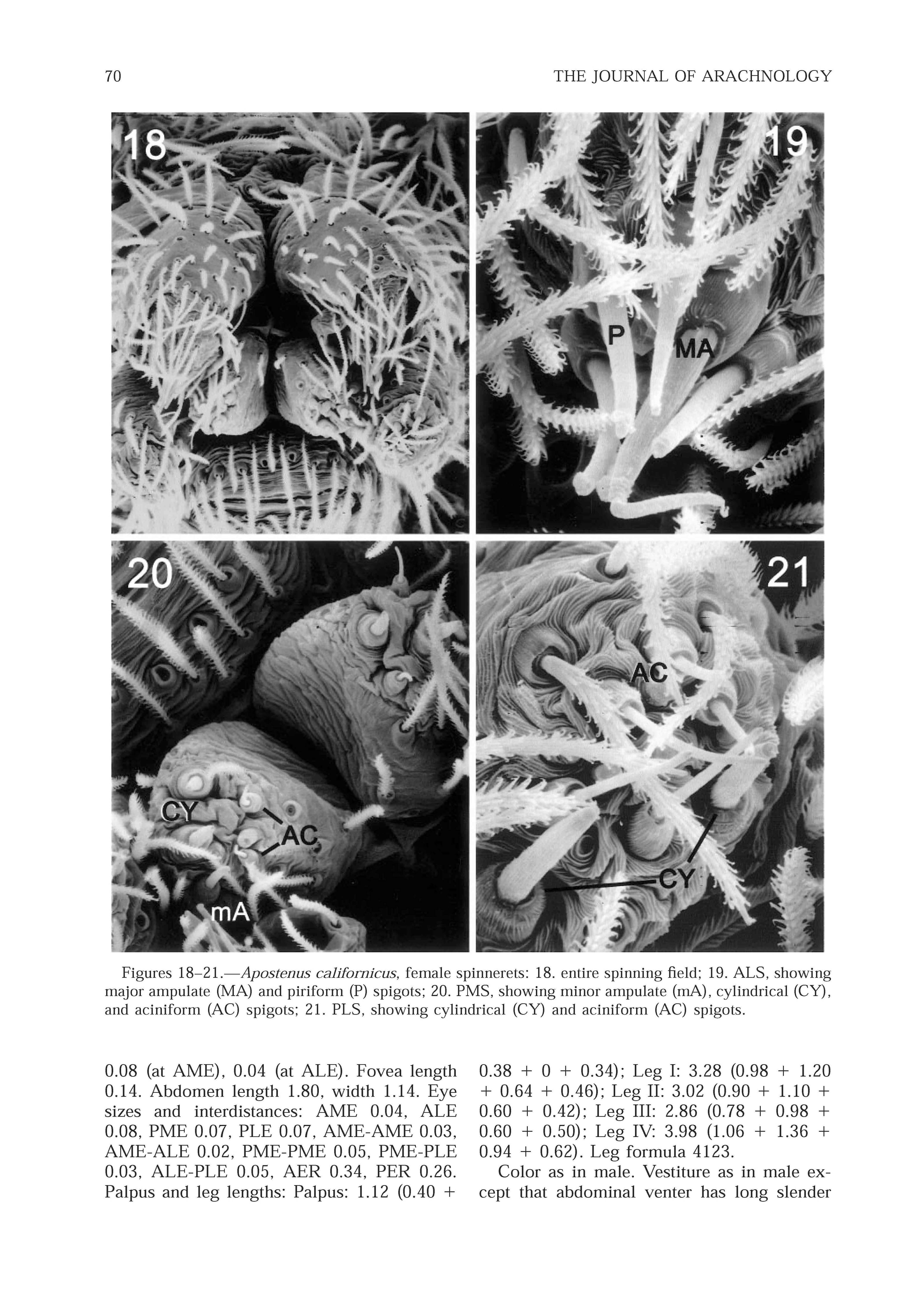
Abdomen: lacking dorsal scute; epigastric furrow lacking epiandrous spigots. Spinnerets with colulus represented by two setae; ALS conical, 2-segmented, contiguous, twice the width of the PLS; ALS with 3 spigots (piriform) and 3 nubbins; PMS with 3 spigots (aciniform) and 2 nubbins; PLS with 3 spigots (aciniform) and 3 nubbins.
Tarsi, metatarsi, and tibiae with dorsal trichobothria in two rows. Tarsi subsegmented; anterior with lateroventral rows of spatulate bristles ( Fig. 3 View Figures 3 - 6 ); posterior longer than anterior, bent in apical third ( Fig. 5 View Figures 3 - 6 ); with two pectinate lateral claws and two broad tenent hairs originating laterad of claws ( Figs. 8-10 View Figures 7 - 10 ). Leg spines: I: metatarsus v2-2-2, tibia v2-2-2-2- 2, femur dl-1-0, p0-0-1, v0-0-0-8 (bristles); 11: metatarsus v2-2-2, tibia v2-2-2-2, femur dl-1-0, v0-0-0-8 (bristles); 111: tibia d2-1, vl-1-0; IV metatarsus d2-2-0, v2-2-0, tibia dl-2-2, v2-2-0.
Palpus: Cymbium with dorsoapical scopula, lacking trichobothria. Bulb with median apophysis sickle-shaped, conductor absent, embolus broad with apical groove and angular base which forms a lock with the subtegulum. RTA short, curved, thorn-like prong. Femur lacking ventral process. ( Figs. 22-24 View Figures 22 - 26 , 27-30 View Figures 27 - 30 )
Variation: Penultimate males lack the modified setae on the venter of the abdomen and the recumbent scales on the carapace and abdomen.
Female (allotype, range o f other females i n parentheses; n = 8): Total length 3.14 (2.30- 3.60). Carapace length 1.14 (1.05-1.32), width 0.94 (0.85-1.05), height 0.47. Clypeus 0.08 (at AME), 0.04 (at ALE). Fovea length 0.14. Abdomen length 1.80, width 1.14. Eye sizes and interdistances: AME 0.04, ALE 0.08, PME 0.07, PLE 0.07, AME-AME 0.03, AME-ALE 0.02, PME-PME 0.05, PME-PLE 0.03, ALE-PLE 0.05, AER 0.34, PER 0.26. Palpus and leg lengths: Palpus: 1.12 (0.40 + 0.38 + 0 + 0.34); Leg I: 3.28 (0.98 + 1.20 + 0.64 + 0.46); Leg 11: 3.02 (0.90 + 1.10 + 0.60 + 0.42); Leg 111: 2.86 (0.78 + 0.98 + 0.60 + 0.50); Leg IV 3.98 (1.06 + 1.36 + 0.94 + 0.62). Leg formula 4123.
Color as in male. Vestiture as in male except that abdominal venter has long slender setae and the carapace and abdomen lack the conspicuous recumbent scales. Form essentially as male except that tarsi are shorter and tarsi IV straighter than in male.
Epigynum with rounded lateral lobes and triangular median lobe; copulatory openings in median grooves. Vulva with 2 rounded spermathecae, short copulatory ducts, and curved fertilization ducts. Spinnerets as in male; PMS conical, not compressed; ALS with 6 long spigots (4 piriforms and 2 larger major ampulates); PMS with 2 large cylindrical spigots, 1 smaller minor ampulate, and 3 small aciniforms; PLS with 2 cylindrical and 5-6 aciniform spigots ( Figs. 18-2 1 View Figures 18 - 21 ).
Sexual dimorphism: Adult males have a vestiture of short setae on the abdominal venter and recumbent white scales on the carapace and abdominal dorsum. Males have tarsi longer, and posterior tarsi more strongly bent, than females.
Biology.-This species is widespread in the mountains of southern California and has been collected from several contiguous localities each in San Diego County and the San Jacinto and San Bernardino Mountains. It is also known from two isolated localities to the west in Kern County, and a tentative record, based on juveniles, to the north in Inyo County. The spider occurs in leaf litter (which varies from moist to slightly dry) of various oak species (with two records from oak and pine duff and one from willow) at elevations from 1040- 2100 m. Males have been taken from September-April, females from September-June. In the lab, juvenile Apostenus were successfully reared to maturity on a diet of Collembola, Psocoptera, and Lepidoptera larvae.
Distribution.-Known only from southern California ( Fig. 2 View Figure 2 ).
DISCUSSION
Our examination of A. californicus has turned up some observations that have not been adequately, if at all, described in the literature. To date, the most complete description of Apostenus is in the recent analysis of the clubionoid genera by Bosselaers & Jocque (2002), to which we can add the following:
Dimorphic abdominal setae.-The presence of short setae ventrally on the male abdomen ( Figs. 15, 16 View Figures 15 - 17 ) was not scored by Bosselaers & Jocque (2002), but occurs in A. californicus and A. fuscus and appears to be an autapomorphy for Apostenus . Although Wunderlich (1999) refers to the presence of modified setae in some species of Agroeca Westring 1861 , of the species we examined, the males have normal setae (A. minuta Banks 1895 , A. pratensis Emerton 1890 , and A. trivittata (Keyserling 1887)) , slightly shorter setae ( A. ornata Banks 1892 ) or short ones interspersed with normal setae (A. brunnea (Blackwall 1833) , J. Bosselaers pers. comm.).
Claw tufts.-The tip of the tarsus bears two spatulate tenent hairs ( Figs. 8-10 View Figures 7 - 10 ) which also appear to be an autapomorphy for the genus. Although this was interpreted as a claw tuft by Bosselaers & Jocque (2002: Character 63), it is clearly not homologous to a true tuft, which is generally understood to arise from the transformed median claw ( Forster 1970). In Apostenus californicus , the modified hairs originate laterad of the paired claws and the region of the median claw is represented by a vestige consisting of a central protuberance and a series of lateral ridges ( Figs. 9, 10 View Figures 7 - 10 ). In a true claw tuft, the modified setae originate from the lateral ridge portion of the median claw vestige, as for example in Drassinella ( Fig. 12 View Figures 11 - 14 ). Such tufts are of a different origin, as are the tufts in 3-clawed spiders. Forster (1970) recorded various forms of claw tuft analogs in several 3-clawed desid genera from New Zealand, and similar analog tufts are also found in the 3- clawed Titiotus Simon 1897 and related tengellids from the Nearctic region ( Figs. 13, 14 View Figures 11 - 14 ). Finally, claws lacking tufts of any sort are found in several liocranid genera, for example, in Liocranum ( Fig. 11 View Figures 11 - 14 ). Detailed observations of these structures will be necessary to determine homology.
Bent posterior tarsi.-All tarsi are subsegmented in both sexes of Apostenus , but tarsi I11 & IV are much more markedly bent in the male ( Figs. 3-6 View Figures 3 - 6 ). Subsegmented and bent posterior tarsi occur in several Holarctic liocranid genera ( Agroeca , Agraecina Simon 1932 , Cybaeodes Simon 1878 , Neoanagraphis Gertsch & Mulaik 1936 , and Scotina Menge 1873 ). This character was first noted by Wunderlich (1999) and interpreted as a synapomorphy by Bosselaers & Jocque (2002: Character 9) for this group of genera. Interestingly, in Apostenus the subsegmented tarsi occur in both sexes, but only in males of Agroeca and Neoanagraphis .
Tegulum/subtegulum lock.- The locking mechanism of the tegulum to subtegulum was first observed by Griswold (1993) in the Lycosoidea and its basal sister groups. A similar locking structure occurs in Apostenus ( Figs. 22 View Figures 22 - 26 , 29 View Figures 27 - 30 ), which has a distinct subtegular lobe, but may differ in having the tegular lobe represented by the embolar base. This locking mechanism was also recorded in Agroeca , Scotina , and Phrurotimpus Chamberlin & Ivie 1935 ( Bosselaers & Jocquk 2002: Characters 130, 131).
Cymbial scopula.-A scopula on the dorsoapical part of the cymbium was not noted by Bosselaers & Jocquk (2002), but occurs in Apostenus californicus ( Fig. 29 View Figures 27 - 30 ), at least in some Agroeca (observed in A. pratensis ), and is also found in a number of lycosoids and their kin ( Griswold 1993).
Epigynum with scape.- Bosselaers & Jocquk (2002, Character 148) interpreted the middle piece of the epigynum of Apostenus as a scape. But unlike a scape, it is broadly attached to the rest of the epigynum ( Figs. 25 View Figures 22 - 26 , 31, 32 View Figures 31 - 32 ) and more closely resembles the median lobe of amaurobiids, lycosoids, and some other spiders.
Embolus insertion.-Although the embolus of A. californicus is apical in position, its insertion as seen in prolateral views is clearly at the middle of the bulb ( Figs. 22 View Figures 22 - 26 , 29, 30 View Figures 27 - 30 ) and not apical as recorded by Bosselaers & Jocquk (2002: Character 140)
Abdominal scute.- Bosselaers & Jocquk (2002) indicate the presence of a male abdominal scute in Apostenus (Character 102); but this was not observed in A. californicus .
Plumose hairs.- Bosselaers & Jocque (2002) state that Apostenus lacks plumose hairs (Character 57); in A. californicus they are present on the legs ( Fig. 7 View Figures 7 - 10 ).
Leg bristles.-These bristles have been interpreted as diminutive spines ( Ubick & Platnick 1991) and occur in a wide number of clubionoids. Although they were not recorded for Apostenus by Bosselaers & Jocquk (2002, Characters 4 & 5), they are present in Apostenus californicus ( Figs. 3, 4 View Figures 3 - 6 ), and also occur, at least on anterior tarsi, in other species of Apostenus and in Agroeca and Liocranoeca .
The family placement of Apostenus is presently in a state of flux. Although traditionally associated with the Liocranidae , the most recent analysis of the clubionoid genera ( Bosselaers & Jocque 2002), argues that the genus belongs neither to the Liocranidae , sensu stricto, nor to the Phrurolithinae (which they transferred to the Corinnidae ) but to an intermediate clade which was not assigned to family. As mentioned above, the genus appears to cluster with the several genera having subsegmented tarsi. These genera also lack claw tufts and precoxal triangles and show some affinities to the lycosoid complex, suggested by the bulbal locking mechanism and cymbial scopula, which may be worth exploring further.







| CAS |
USA, California, San Francisco, California Academy of Sciences |
| UCR |
USA, California, Riverside, University of California |
| TRP |
TRP |
| SDM |
SDM |
| CDU |
CDU |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |