
Mimobdella japonica Blanchard, 1897
|
publication ID |
https://doi.org/ 10.12782/sd.21.1.049 |
|
DOI |
https://doi.org/10.5281/zenodo.4734108 |
|
persistent identifier |
https://treatment.plazi.org/id/F247911D-A129-FFB7-FC46-FD6EC4B5FBE0 |
|
treatment provided by |
Felipe |
|
scientific name |
Mimobdella japonica Blanchard, 1897 |
| status |
|
Mimobdella japonica Blanchard, 1897
( Figs 1–3 View Fig View Fig View Fig )
Mimobdella japonica Blanchard, 1897: 94 , 95, pl. 6, figs 16, 17; Nakano 2011: 3–7, figs 1–4; Nakano 2013: 100–103 View Cited Treatment , figs 2–4.
Material examined. Three specimens collected from Nakanogo , Hachijo, Hachijojima island, Tokyo, Japan (33.071886°N, 139.811194°E, elevation 130 m), by Victor Benno Meyer-Rochow: KUZ Z1653, dissected, collected from inside a rotten wooden log, on 19 April 2015; KUZ Z1657, in soil under a rotten wooden log, on 10 July 2015; and KUZ Z1658, dissected, in soil under a rotten wooden log, on 11 July 2015 GoogleMaps .
Description. BL 68.2–91.1 mm, BW 6.2–8.8 mm ( Fig. View Fig 1). Caudal sucker ventral, elliptic, CL 3.8–4.2 mm, CW 4.3– 5.0 mm ( Figs 1B View Fig , 2D View Fig ).
Annulation of somites I–VII unclear, comprising 16–17 annuli altogether; according to annuli formation, annulation of somites I–VII tentatively interpreted as follows: somite I merged with prostomium; somite II uniannulate; somite III uni- or biannulate; somite IV bi- or triannulate; somite V triannulate; somite VI 4-annulate; somite VII basically 4-annulate; possibly somite V a3 to somite VI a2 forming posterior margin of oral sucker ( Fig. 2A, B View Fig ). Somite VIII 5-, or 6-annulate. Somite IX 6-, or 7-annulate. Somites X and XI 7-annulate ( Fig. 2E View Fig ). Somites XII and XIII 7-, or 9-annulate, a2 with slight furrow ( Fig. 2E View Fig ). Somites XIV–XXIII 9-annulate; a2 of each somite in somites XVII–XXIII with slight secondary furrow. Somite XXIV 6-annulate ( Fig. 2C, D View Fig ). Annulation of somites XXV–XXVII, comprising 5–8 annuli altogether, hardly discernable; 158th or 159th annulus last complete annulus on venter; according to annuli formation, annulation of somites XXV–XXVII tentatively interpreted as follows: somite XXV tri-, or 4-annulate; somite XXVI biannulate; somite XXVII uni- or triannulate. Anus between 159th [XXVI (a1 + a2)] and 160th (XXVI a3) annuli with 2 or 3 post-anal annuli ( Fig. 2C, D View Fig ).
X b5 (c9) and XIII a2, respectively, first and last annuli of clitellum ( Fig. 2E View Fig ).
Male gonopore in XI/XII. Female gonopore in XII/XIII ( Fig. 2E View Fig ). Gonopores separated by one full somite.
Eyes undetectable. Papillae numerous, minute, hardly visible, one row on every annulus, and 2 or 3 rows on annuli with secondary furrow(s). Nephridiopores, in 17 pairs, in somites VIII–XXIV ( Fig. 2B, D, E View Fig ).
Pharynx reaching to XIV/XV (KUZ Z1653) or XV c1 (KUZ Z1658), with 3 myognaths separated by triangular paragnaths, each myognath bearing two conical stylets arranged in tandem. Crop reaching to XXI a2/c9; terminal end of crop forming sphincter between crop and intestine, in XXI c1 (KUZ Z1653) or c1/c2 (KUZ Z1658) to XXI a2/ c9; pair of post-crop caeca, right post-crop caecum in XXI c1–d22 (KUZ Z1653) or XX d22–XI d22 (KUZ Z1658), lefl post-crop caecum in XXI c1–d22 (KUZ Z1653) or XX c12– XXI d21 (KUZ Z1658) ( Fig. 2F View Fig ). Intestine reaching to XXIV c2/b2 (KUZ Z1653) or a2/b5 (c9) (KUZ Z1658). Rectum tubular, thin-walled, reaching straight to anus.
Testisacs multiple ( Fig. 2G View Fig ), ca. 220–260 on each side (KUZ Z1653) or uncountable (KUZ Z1658); on right side, in XVI c2 to 155th (KUZ Z1653) or 156th (KUZ Z1658) annulus (XXV b1), in total ca. 220 testisacs, ca. 20 in XVI, 30 in XVII, 27 in XVIII, 27 in XIX, 26 in XX, 26 in XXI, 23 in XXII, 27 in XXIII, 15 in XXIV, 5 in XXV; on lefl side, in XVI b2 to 156th (KUZ Z1653) (XXV b2) or 155th (KUZ Z1658) annulus (XXIV b6), in total ca. 260 testisacs, ca. 16 in XVI, 30 in XVII, 32 in XVIII, 26 in XIX, 31 in XX, 35 in XXI, 33 in XXII, 19 in XXIII, 32 in XXIV, 5 in XXV. Sperm duct paired; right sperm duct in XI c12 to XVI c2 (KUZ Z1653) or b2/a2 (KUZ Z1658); lefl sperm duct in XI c12 to XVI b2 (KUZ Z1653) or a2/c9 (KUZ Z1658). Paired atrial cornua in XI c11 (d21) to XII c1–c2. Male atrium in XI c11 (d21) to XII c1–c2.
One pair of ovisacs; right ovisac descending directly to XXII c9 (KUZ Z1653) or descending to XIX c10/d21, turning anteriorly to reach XIX c1, then either turning posteriorly to XIX c2 (KUZ Z1658) ( Fig. 2G View Fig ); lefl ovisac descending directly to XXII a2 (KUZ Z1653) or descending to XXI b2/a2, turning anteriorly to reach XX c10, then either turning posteriorly to XX d21 (KUZ Z1658) ( Fig. 2G View Fig ); both ovisacs converging in XIII c1; then each ovisac descending directly to female gonopore.
Colouration. In life, dorsal surface ochre; ventral surface pinkish gray ( Fig. 3 View Fig ). Colour faded in preserved specimens; clitellum obvious, slightly deeper than surrounding body parts ( Fig. 1 View Fig ).
Distribution. This species has been recorded previously from Amamioshima and Okinawajima islands in the Ryukyu Islands, Japan ( Nakano 2013). Herein, we report its first record from Hachijojima island, Izu Islands, Japan ( Fig. 4 View Fig ).
Natural history. Mimobdella japonica has been reported to be semi-aquatic ( Nakano 2013), but all three individuals from Hachijojima were collected from terrestrial habitats. All had an obvious clitellum. Based on their dates of collection, we deduce that the reproductive season of this species on Hachijojima begins in April and continues until at least early July. It seems to have established a breeding population on this island despite the island’s being ~ 500– 700 km further north and having a colder climate than its other known localities in the Ryukyu Islands.
Genetic distances. Three specimens from Hachijojima, two from Amamioshima, and four from Okinawajima had completely identical sequences of COI (1,267 bp), tRNACys– 16S (1,004 bp), and tRNALeu (34 bp). The pairwise ND1 uncorrected p -distance among these nine specimens was 0.0– 0.2% ( Table 2 View Table 2 ). The 570th bp position (counted from the first position, thus in the third position of the codon) of the ND1 sequences of the two specimens from Okinawajima (KUZ Z721 and Z722) was T (it was C in the other seven sequences; Fig. 4 View Fig ).
Remarks. The specimens collected from Hachijojima were clearly identified as Mimobdella japonica based on the diagnostic characteristics defined by Nakano (2013): midbody somites 9-annulate; anus with 2 or 3 post-anal annuli; male gonopore in XI/XII, female gonopore in XII/XIII, gonopores separated by one full somite; post-crop caeca in pairs in XX and XXI; sperm duct reaching to level anterior of middle part of XVI; and ovisacs long, reaching to XIX– XXII. In addition, their COI, tRNACys– 16S, and tRNALeu– ND1 sequences were identical to those of specimens collected from the Ryukyu Islands. These genetic data confirm the implications of the morphological data that these specimens are indeed M. japonica .
Considering that all of the examined specimens including individuals from Hachijojima have essentially identical sequences of mitochondrial genes, it is conceivable that M. japonica has arrived in its known localities via recent dispersal events, or that it has been introduced via human activities. Hirano et al. (2014) considered the low genetic diversity of the bradybaenid land snail Bradybaena phaeogramma (Ancey, 1888) , which is distributed in the Izu Islands as well as the Ryukyu Islands, and concluded that its distribution is the result of long-distance dispersal via an oceanic current. Despite this, some other terrestrial invertebrates are considered to have been introduced into Hachijojima via human activities. Karasawa et al. (2015) and Umezawa (1973) reported the whip scorpion Typopeltis stimpsonii (Wood, 1862) , originally from Amamioshima and adjacent islands, in Hachijojima. Umezawa (1973) suggested that this whip scorpion was introduced to the island attaching with cycads in 1968. Fujiyama et al. (2012) and Meyer-Rochow (2015) reported mass outbreaks on Hachijojima of the polydesmid diplopod Chamberlinius hualienensis Wang, 1956 , originally known from Hualien County, Taiwan ( Wang 1956). Fujiyama et al. (2012) stated that this Taiwanese diplopod was first found on Hachijojima in 2002.
The distributional history of this species remains unveiled because the number of examined specimens of M. japonica is still severely limited, and the distribution of this leech species is not fully revealed. Although two of the four specimens from Okinawajima island had ND1 sequence that were identical to specimens from the other two islands, the other two specimens from Okinawajima had unique ND1 sequences ( Fig. 4 View Fig ). However, it is possible that this unique ND1 sequence will be detected from leeches in other localities in the future, which could further elucidate the distributional history of this species. Further faunal, distributional, and genetic studies are thus essential to determine the distributional history of M. japonica .
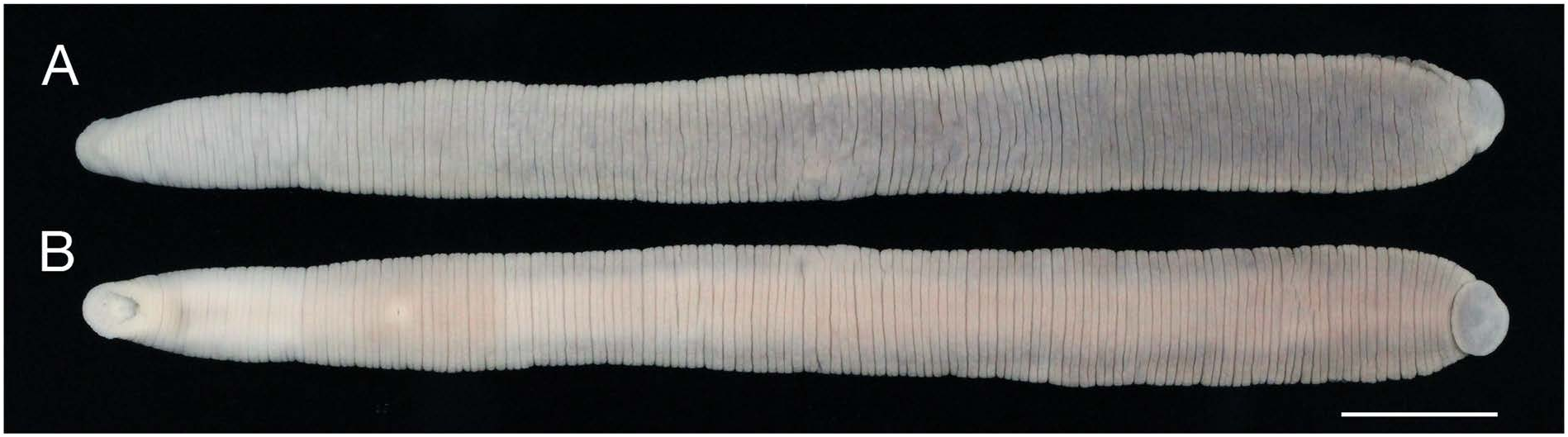
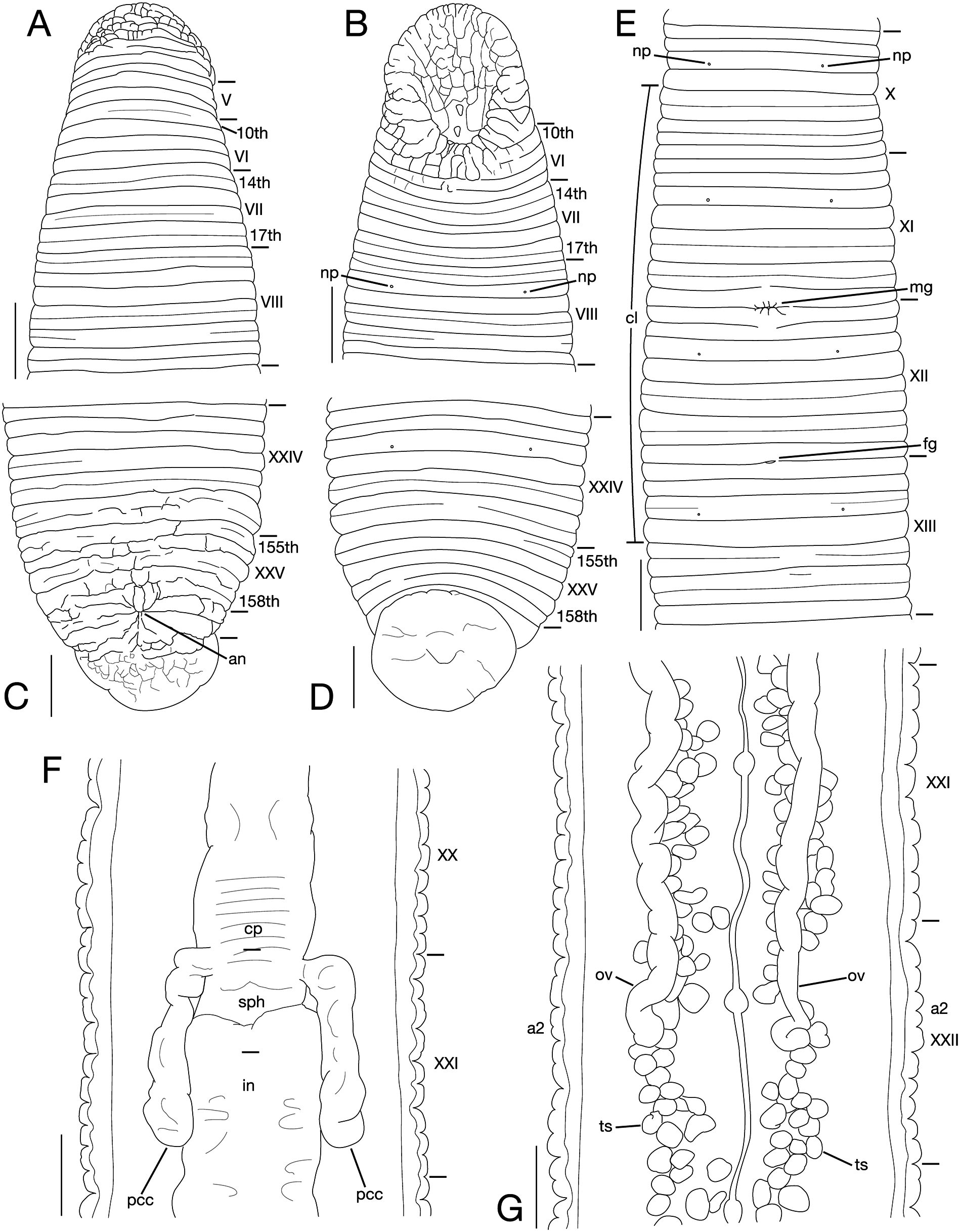

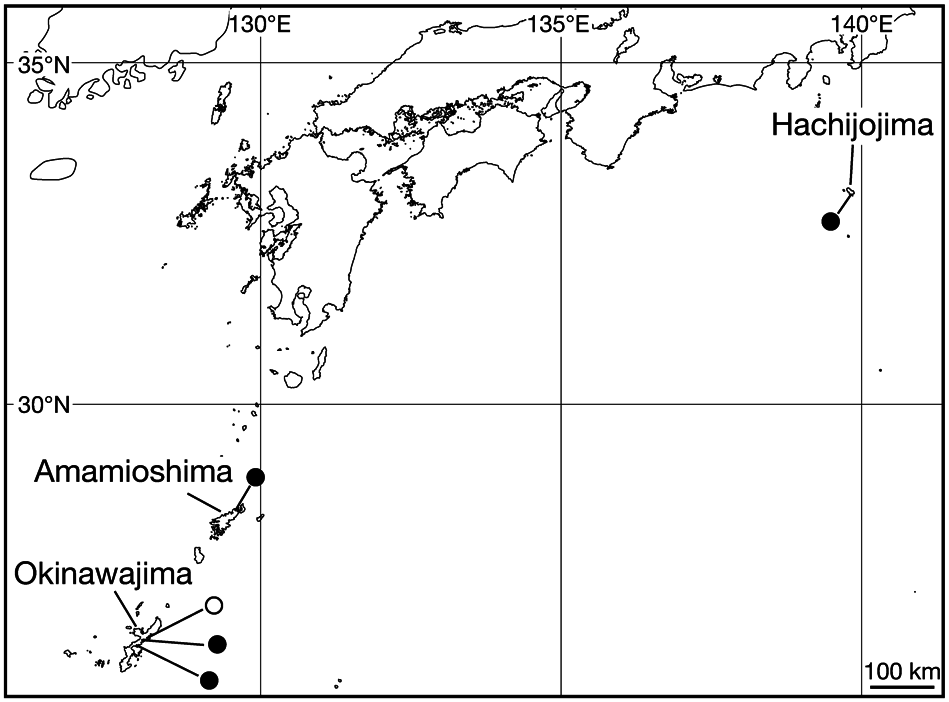
Table 2. Uncorrected p-distances for 594bp of the available NADH dehydrogenase subunit 1 sequences of Mimobdella japonica Blanchard, 1897. Acronym for voucher registration numbers: KUZ, Zoological Collection of Kyoto University.
| Voucher Reg. No. | (1) | (2) | (3) | (4) | (5) | (6) | (7) | (8) | (9) |
|---|---|---|---|---|---|---|---|---|---|
| (1) KUZ Z179 | |||||||||
| (2) KUZ Z228 | 0.000 | ||||||||
| (3) KUZ Z227 | 0.000 | 0.000 | |||||||
| (4) KUZ Z229 | 0.000 | 0.000 | 0.000 | ||||||
| (5) KUZ Z721 | 0.002 | 0.002 | 0.002 | 0.002 | |||||
| (6) KUZ Z722 | 0.002 | 0.002 | 0.002 | 0.002 | 0.000 | ||||
| (7) KUZ Z1653 | 0.000 | 0.000 | 0.000 | 0.000 | 0.002 | 0.002 | |||
| (8) KUZ Z1657 | 0.000 | 0.000 | 0.000 | 0.000 | 0.002 | 0.002 | 0.000 | ||
| (9) KUZ Z1658 | 0.000 | 0.000 | 0.000 | 0.000 | 0.002 | 0.002 | 0.000 | 0.000 | |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Mimobdella japonica Blanchard, 1897
| Nakano, Takafumi & Meyer-Rochow, Victor Benno 2016 |