
Mokopirirakau galaxias, Knox & Hitchmough & Nielsen & Jewell & Bell, 2021
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4964.1.7 |
|
publication LSID |
lsid:zoobank.org:pub:E704BAD8-917E-43B0-9FD9-B32EA1A36BE9 |
|
DOI |
https://doi.org/10.5281/zenodo.4741134 |
|
persistent identifier |
https://treatment.plazi.org/id/40D87BBC-EFD2-47F6-9CDD-5E4FEBCB3849 |
|
taxon LSID |
lsid:zoobank.org:act:40D87BBC-EFD2-47F6-9CDD-5E4FEBCB3849 |
|
treatment provided by |
Plazi |
|
scientific name |
Mokopirirakau galaxias |
| status |
sp. nov. |
Mokopirirakau galaxias sp. nov.
( Figs 2–5 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 )
Holotype. Adult male NZMZ RE.008613, from an undisclosed location within Oteake Conservation Park, North Otago, New Zealand, collected by C. Knox and T. Jewell on 14 January 2020. The collection site (and all other known populations of Mokopirirakau galaxias sp. nov.) are between latitudes of 44º 35’S–44º 58’S, longitudes of 169º 43’E–170º 19’E, and altitudes of 1,300-1,600 m ASL. GoogleMaps
Paratypes. None (but see Variation section below).
Etymology. Latin (from Ancient Greek) galaxias for the patterning of small white spots on the dorsal and lateral surfaces of the body reminiscent of the Milky Way. The epithet is formed as a noun in apposition.
Suggested Common Name. ‘Hura te ao gecko.’ From te reo Mâori, the indigenous language of New Zealand’s first people, for ‘breaking dawn.’ ‘Hura’ commonly means ‘reveal,’ but can also be applied colloquially to signify ‘the break of dawn.’ This concept ties in with aspects of this species’ colouration (i.e., the orange-yellow around the mouth, evocative of sunlight at daybreak), as well as the idea that this lizard has for the first time revealed itself to us.
Diagnosis. Both the phylogenetic analysis and the combination of straight rather than strongly curved or vshaped lamellae, presence of brightly coloured mouth pigment, absence of apical plates from some toes, lamellae forming a pad which does not extend to the end of the toe, and ventral scales on the tail being roughly twice the size of dorsal scales, identify the species as a member of the genus Mokopirirakau .
The species can readily be distinguished from all Mokopirirakau species , except M. kahutarae , on the basis of eye colour (very dark brown versus grey, light brown or occasionally light blue with dark ‘filigree” markings), body colour pattern (olive-grey to olive-green with small white spots versus grey with transverse bands), and head colour pattern (contrasting v-shaped dark marking behind eyes and pale stripe from eye to ear absent in this species and M. kahutarae ; present in all others). Distinguished from M. kahutarae , to which it has some obvious similarities (see Whitaker 1984), on the basis of a number of characters: on average smaller body size (76–84 mm SVL versus up to 91 mm); snout slightly more elongate (longer than versus equal to eye-ear distance); tail length shorter than SVL (versus equal or longer); distal attenuation of digits ~25% (versus 31%), with webbing extending to approximately half of the distal phalange (versus approximately one quarter); 7–9 subdigital lamellae (versus 12–14); apical plates slightly enlarged on the first 3 digits of each autopodium (versus absent); single scale row between middle of orbit and labial scales (versus 2 rows); 2–3 supraciliary scale rows (versus 4–5), forming an ‘eyebrow,’ with supraciliary scales scarcely enlarged (versus greatly enlarged), with the effect of upper margin of eye fully visible from the side (versus hidden by an ‘eyebrow’); 8–9 infralabial scales (versus 11–12); ~18 ventral scale rows from axilla to groin (versus ~24); two (versus three) cloacal spurs; iris very dark brown (versus totally black); pupil margin very dark brown (versus black); labial scales and above surrounding scales, as well as the palmar surfaces of the manus, pes, and digits coloured vivid orange (versus muted olive-grey); mouth and throat colour orange-pink and grey-black, respectively (versus pink to pale red); base colouration of dorsal skin more consistently olive (versus olive to slate- grey); and dorsal markings composed of numerous pale spots with narrow, asymmetrical paler transverse dorsal markings (sometimes absent or indistinct) (versus 6–7 wide pale grey bands).
Table 1 View TABLE 1 compares characters of M. galaxias sp. nov. with all other described Mokopirirakau species. Diagnostics are not given for any other non-described Mokopirirakau species on the basis that formal taxonomic descriptions for these are in preparation (R. Hitchmough, S.V. Nielsen and A. Bauer, unpub. data); however, none of these resemble M. galaxias and all are allopatrically distributed.
Description of holotype. Adult male, 68 mm SVL, 65 mm TL. Dorsal scales fine, regular. Ventral scales regular, larger than dorsal. Tail cylindrical, tapering. Eye large. Snout blunt. Rostral broader than long, with deep cleft, c. 50% of scale. Nasals 3. Internasals 1, Supralabials/upper labials 11. Infralabials/lower labials 9. Mental 1, broader than wide. Postmentals 1. Precloacal pores. 20 wide, 4 deep. Cloacal spurs 2. Digits moderate, narrow. Lamellae 9, straight. Measurements (in mm): HL 23.5; HW 15.5; AG 31; SF 29.5; S-E 18; ED 3.8; E-E 10.2; EF 12.5. Ratios: HL/ED 6.18. SE/EE 1.76; TL/SVL 0.95.
Colour. Base body colour olive-grey to olive-green on the dorsum with a light grey venter. Dorsal markings indistinct, consisting of thin blotches and/or streaks extending outward from the midline or slightly alternating and overlapping the midline, markings coalesce to form a broader patch on top of head, and continue down tail. Small, whitish spots on dorsal and particularly numerous along the flanks, sometimes aligning to form secondary, transverse bars between main dorsal markings; spots vary in intensity between individuals, but often dominate the overall colour pattern. Head markings subtle, indiscernible or absent. Eye particularly large, lined with orange, almost touching labial scales, with enlarged supraciliary fringe creating slightly ‘hooded’ eyebrow. Iris dark blackish brown; black pupil with paler edges just discernible. Labials and adjacent scales distinctively pale yellow-olive to orange. Tongue light pink in middle, darker stippled pink at tip, with bright orange base. Roof of mouth bright orange on and around gums; coloured from pink anteriorly to mid-grey to black at back and in throat. Floor of mouth pink, with bright orange at base of tongue and thin lining along inner margin of gums. Ventral very finely peppered with lighter and darker specks. Cloacal opening bright orange. Soles and undersurface of digits pale to bold creamyorange.
Variation. Twenty one live individuals (eight adult females, five adult males, four subadult females, three subadult males, and one juvenile) were captured, measured, photographed, and released across the four known sites between January 2018 and December 2020. The following data is from the first six specimens captured: adult males, adult females; 76–84 mm SVL (x=80, SD 3.5). Dorsal and ventral scales as for holotype. Tail cylindrical, tapering. Eye large. Snout blunt. Rostral, broader than long, with deep cleft c. 50% of scale. Nasals 3 or 4. Internasals 0–2. Supralabials/upper labials 9–11, Infralabials/lower labials 8–9, usually 9. Mental 1. Postmentals 1. Precloacal pores 21–24 wide, 4–5 deep. Cloacal spurs 2–4 in males only. Digits moderate, narrow. Lamellae 7–9. straight. Ratios: HL/ED 4.2–4.5 (0.15), SE/EE1.10–1.28 (0.07), TL/SVL 0.83–0.89 (0.02). Twenty specimens had intact original tails and these were consistently much shorter than the snout-to-vent length. The exception was an adult male for which almost the whole tail was regenerated, yet was still the same proportional length as for those with their original tails.
Distribution. Mokopirirakau galaxias sp. nov. appears to be an alpine specialist and is known only from Oteake Conservation Park. This species is currently known from c.8 rock tors and bluffs within the 1.4-ha type locality, plus three additional sites in deep boulderfield habitat. It could possibly occur elsewhere in Otago and Canterbury, despite different rock types and other Mokopirirakau species being present in some areas, if sufficient niche separation is possible. Both M. kahutarae and M. galaxias are thought to be solely alpine in nature; however, distributional limits and pre-human altitudinal limits of both are unknown. The ~400-km distributional gap between them may at least partly be an artefact of poor sampling, particularly given the difficult access to many of their habitats. Suitably creviced rock exposures, deep boulderfields and talus slopes occur widely in the mountain ranges between their known distributions but are patchy in distribution.
Environment. The climate in the 64.03 Hawkdun and 64.04 Saint Bathans Ecological Districts is semi-arid with warm, dry summers and cold winters ( McEwen 1987). LENZ Level IV data indicate the land environment at the type locality is particularly harsh, with an annual average temperature of 4.2 °C, a mean minimum temperature of -4.1 °C during the coldest month of the austral winter (July), average annual solar radiation of 13.2 MJ/m2/day and winter solar radiation of 3.8 MJ/m2 /day ( Leathwick et al. 2002).
Habitat. Fractured and deeply creviced greywacke outcrops and deep boulderfields and talus slopes. Snow tussock ( Chionochloa rigida ) grassland and Dracophyllum rosmarinifolium shrubland are the predominant vegetation types in the areas where the species has been found, interrupted by greywacke rock outcrops or boulderfields and talus. At the type locality, M. galaxias have been found on six discrete, but proximate rock outcrops and bluffs comprised of a protruding but deeply fissured and fractured central tor several metres tall, surrounded by varying amounts of rock talus, fellfield and/or scree, along a ~270-m section of a ridgeline. The first gecko discovered was located within a crevice inside a c. 50 cm fractured rock, in an expanse of stony ground c. 10 m from a major outcrop system. The other eighteen geckos encountered at the type locality have all been inside vertical rock fissures, or out on rock ledges. At the other three sites (across two mountain ranges) the five geckos observed were emergent in deep boulderfield or talus, but three of these were well concealed by surrounding rocks and were in positions close to retreats or escape routes. Some of the rocks at all known sites, particularly on tors, are heavily encrusted with colourful lichens of various species, including large amounts of the tall, bristly Neuropogon spp. , the latter which may limit the extent of usable rock surface for the geckos ( Fig. 6 View FIGURE 6 ).
Natural history. Mokopirirakau galaxias is apparently an alpine specialist gecko, saxicolous in greywacke rock outcrops, boulderfields, and talus. The geckos are strongly nocturnal and thigmothermic. They have been detected on exposed rock surfaces during surveys at night in air temperatures of 9-16 °C. Nothing is known of the species’ life history, diet or reproduction. The sex ratio appears to be close to 1:1 based on the limited survey results to date (C. Knox, unpub. data). A small number of juvenile and subadult geckos have been located, but not neonates. It is likely to be mostly insectivorous, but may consume berries of alpine shrubs where available. It is likely to be viviparous, as all New Zealand geckos are. However, due to the species’ alpine niche, it is most likely slow-growing, late to mature (at least 5 years) and long lived (to decadal timeframes), with a very low reproductive output ( Cree 1994, Cree & Hare 2016; Knox et al. 2019). One of two adult females captured in January 2018 and a female captured in November 2020 were pregnant; however, no inference can be made on the annual reproductive output per female based on this very small sample size.
At all known sites M. galaxias is sympatric with the Southern Alps gecko ( Woodworthia “Southern Alps”), which is sometimes numerous and occupies all available rocky habitat from outcrops to talus and scree. The two gecko species can be syntopic, sharing the same rock crevice or foraging on the same rock ledge. Size differences between M. galaxias (to 84 mm SVL) and W. “Southern Alps” (c. 55-65 mm SVL) may allow resource partitioning in retreat site and prey selection between the two species. In contrast, Whitaker et al. (1999) noted that in Nelson and Marlborough, M. kahutarae appear to exclude specimens of the sympatric Woodworthia geckos from their immediate territory. Several species of Oligosoma skinks are also present, particularly O. maccanni , which are found throughout the same rocky habitat and in the surrounding alpine tussockland.
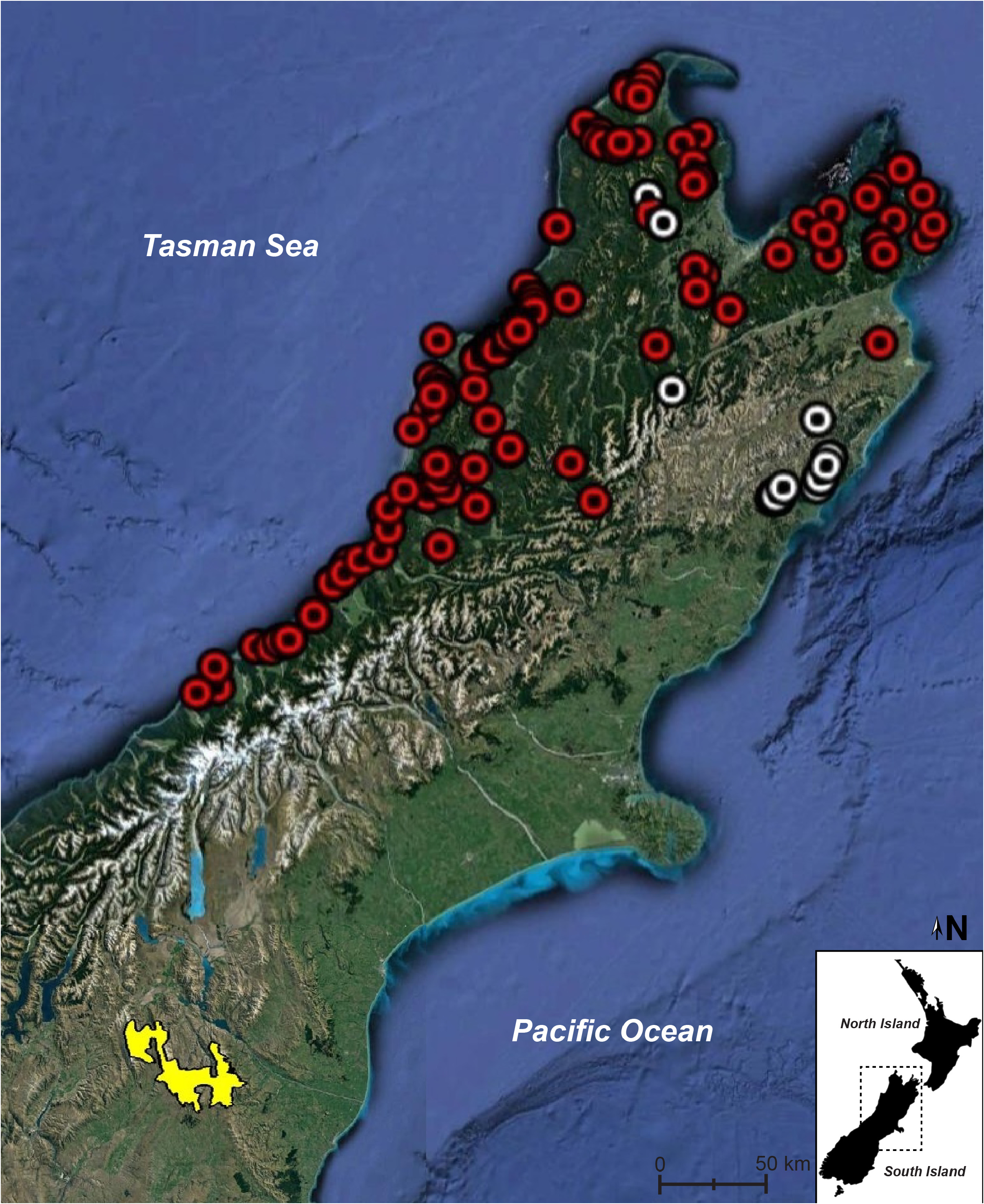
Phylogenetic affinities. Genetic analyses using sequences from the mitochondrial ND2 gene indicate that M. galaxias is sister to a clade comprising the sister species M. granulatus and M. kahutarae . Mokopirirakau granulatus occurs in the upper half of the North Island, and in Marlborough, Tasman, and Westland in the South Island. The closest M. kahutarae populations to M. galaxias are approximately 400 km to the north-east; whereas, the closest M. granulatus populations to M. galaxias are approximately 180 km away, but on the other side of the Southern Alps ( Fig. 7 View FIGURE 7 ). Thus, M. galaxias is completely isolated from its nearest relatives geographically, with multiple mountain ranges between their known ranges. The geographically closest Mokopirirakau populations are of the undescribed taxon M. “Roy’s Peak”, which occurs to the west and south-west (van Winkel et al. 2018; Knox et al. 2019).
Conservation. At the type locality this species is currently known only from eight rock tors and surrounding talus within a 1.4-ha area, and <20 individual animals have been recorded from three surveys. Although difficult to estimate, abundance at the type locality is likely to be <100 geckos, given the small area of habitat. Initial surveys from January 2018 to February 2020 (mostly within a 4 km radius of the known site) were undertaken in suitable weather (>10 °C, dry and calm nights), but did not locate new populations. However, in November and December 2020, three new populations were found, including two sites in another mountain range within Oteake Conservation Park. Given the November and December 2020 finds, it is deemed likely that other populations of this species will be discovered, as further survey work is undertaken in North Otago and South Canterbury.
Threats to M. galaxias include predation from introduced mammals, illegal collection, and climate change. In addition, fires and grazing by introduced herbivores may be a threat by reducing habitat quality and food availability. Introduced mammalian carnivores and rodents that may encroach into their high altitude, exposed habitat pose a threat, especially predators that hunt at night when the geckos may be emergent on rock surfaces. These are likely to include cats (Felix catus), ferrets ( Mustela furo ), stoats ( Mustela erminea ), weasels ( Mustela nivalis ), hedgehogs ( Erinaceus europaeus ), possums ( Trichosurus vulpecula ), and mice ( Mus musculus ) ( O’Donnell et al. 2017; Josh Kemp pers. comm.). Mice are likely to have population irruptions following tussock mast seeding, which will then drive secondary carnivore irruptions. Stoats are known to prey on alpine lizards ( O’Donnell et al. 2017), and mice and weasels can squeeze into tight crevices to access the geckos inside retreats. Illegal collection has emerged as a threat to several species of New Zealand lizards, particularly geckos ( Knox et al. 2013; Marshall et al. 2020) and if precise site information becomes publicly available then determined smugglers could impact the abundance of geckos at those sites. Climate change may also render any remaining habitat increasingly vulnerable to introduced mammals, as it is expected that more predators will infiltrate alpine areas as the climate warms ( O’Donnell et al. 2017).
Due to the species being discovered very recently, it is not yet listed in the most recent iteration of the Department of Conservation’s New Zealand Threat Classification System lists ( Hitchmough et al. 2016). Typically, newly discovered species are placed in the “Data Deficient” category. However, the system requires a precautionary ap- proach under which, provided some information exists, the species is placed into the most threatened category consistent with what is known, with uncertainty being indicated by a “Data Poor” qualifier. We recommend the species be assigned the conservation ranking of “Threatened—Nationally Endangered” with the qualifier “Data Poor” ( Townsend et al. 2008). This is because of its low abundance, likely decline resulting from impacts of predatory mammals, threats from climate change, and highly restricted range (four small locations). This species should be a high priority for the Department of Conservation for surveys to improve understanding of its distribution, conservation status and conservation management requirements.
In the IUCN Red List system, in the absence of clear information on population trend, Data Deficient seems the best recommendation for listing.





| T |
Tavera, Department of Geology and Geophysics |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |