
Pheretima monoporata, James, 2004
|
publication ID |
https://doi.org/ 10.5281/zenodo.4618925 |
|
persistent identifier |
https://treatment.plazi.org/id/9918E954-FFBF-E06C-099A-FA5951C7FCB8 |
|
treatment provided by |
Carolina |
|
scientific name |
Pheretima monoporata |
| status |
sp. nov. |
Pheretima monoporata , new species
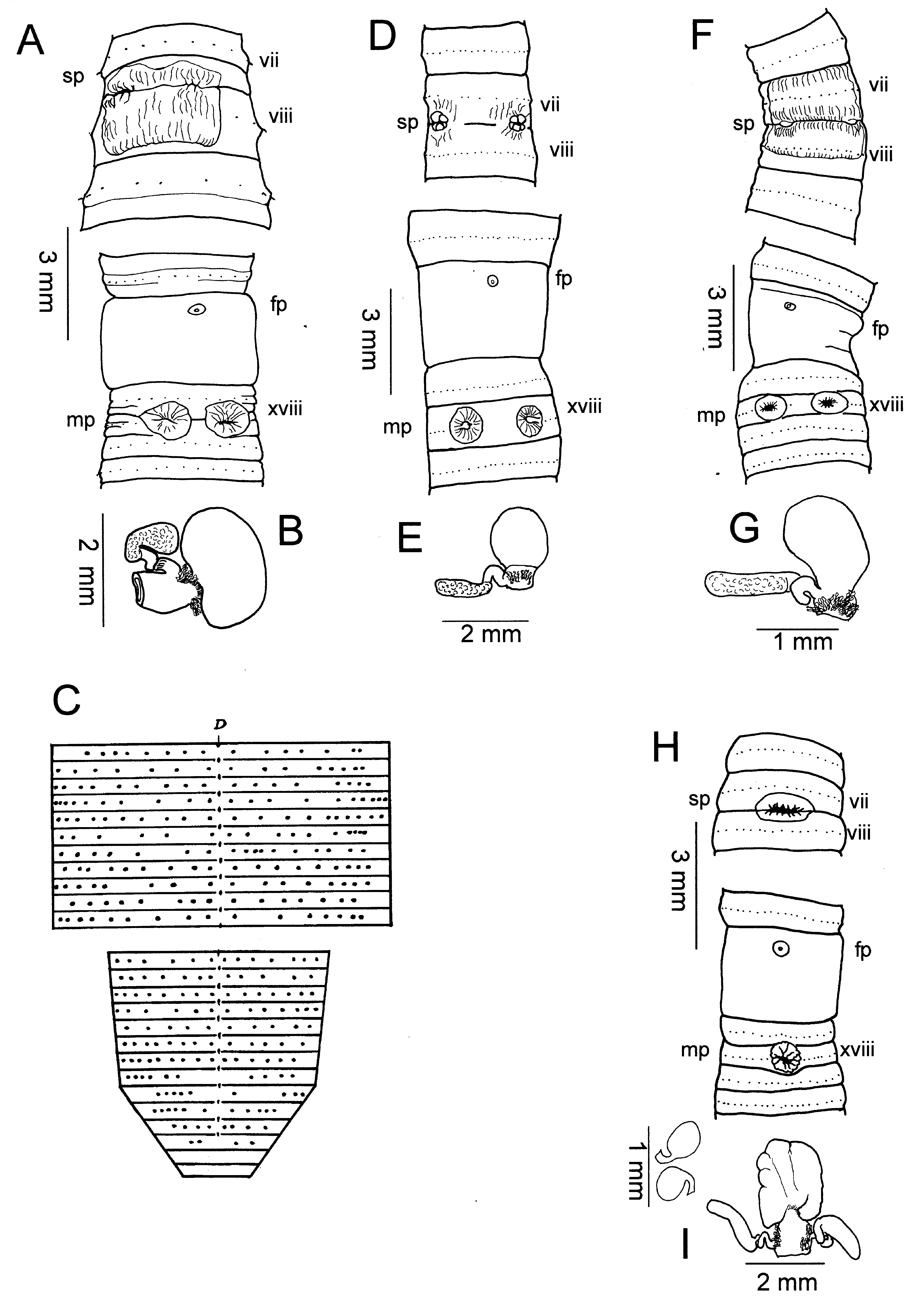
( Figs. 2H, I View Fig )
Material examined. – Holotype - adult, ( NMA 003981 ), Philippines, Mindanao Island, Bukidnon Province, Mt. Kitanglad Range , 16.5 km S, 4 km. E of Camp Phillips, 8 10.5' N, 124 51' E, 1900 m. elevation, coll. L. Heaney, 22 Mar.1993. GoogleMaps
Paratypes – 2 adults, 2 late juveniles ( FMNH 011066 View Materials ), Philippines, Mindanao Island, Bukidnon Province, Mt. Kitanglad Range , 16.5 km S, 4 km. E of Camp Phillips, 8 10.5' N, 124 51' E, 1900 m. elevation, coll. D. Balete, no date GoogleMaps .
Others – 1 adult ( KUNHM 002144 ), Philippines, Mindanao Island, Bukidnon Province, Mt. Kitanglad Range , 8 9' N, 124 44' E, 1800 m. elevation, coll. D. Balete, no date GoogleMaps ; 1 adult ( KUNHM 002145 ), Philippines, Mindanao Island, Bukidnon Province, Mt. Kitanglad Range , 18.5 km S, 4 km E of Camp Phillips, 8 9.5' N, 124 51' E, 2250 m elevation, coll. D. Balete, no date GoogleMaps .
Etymology. – The species name comes from the presence of a single spermathecal pore and male pores so closely approximated as to appear to be one pore.
Description. – Faint brown antero-dorsal pigmentation, body 60-70 x 2.2-2.9 mm (vii), 2.5-2.9 (x), 2.3-3.2 mm (xxv), 99, 101 segments (several specimens were amputees); body cylindrical in cross-section. First dorsal pore 12/13 (4), 13/ 14 (1), spermathecal pore single in 7/8, female pore single in xiv, openings of copulatory bursae paired in xviii, 0.05 circumference apart between 2 nd and 3 rd setal lines, 0 setae between openings. Setae regularly distributed around segmental equators; 26-34 setae on vii, 32-36 setae on xx; in vii ZZ: YZ = 2.0-4.0, in xx ZZ: YZ = 3-3.5, no ventral gaps. Clitellum annular xiv-xvi; no other genital markings ( Fig. 2H View Fig ).
Septa 4/5-6/7 thin, 7/8 muscular, 8/9-10/11 thin, 11/12,12/ 13 slightly muscular. Large gizzard in viii, esophagus with low vertical lamellae x-xiii, xiv, intestinal origin xvii, simple caeca originating in xxvii, extending forward to xxv, xxiii; typhlosole xxvii-lxxiii, lxv, lx, simple fold 0.4 lumen diameter; intestinal wall with 18-20 longitudinal blood vessels xxvii-xxxix.
Hearts x-xiii esophageal, commissural vessels vi, vii, ix lateral, those of ix either greatly different in size or one side lacking; viii to gizzard; supra-esophageal vessel x-xiv; extra esophageal vessel joins ventral esophageal wall in x, receives efferent parieto-esophageal vessel in xiii.
Dense tufts of nephridia on anterior faces of 5/6, 6/7; nephridia of intestinal segments located at septum/body wall junction on anterior and posterior faces of septa, during dissection nephridia come away on septa.
Ovaries and funnels free in xiii, spermatheca single, preseptal in vii with nephridia on duct; spermatheca with large ampulla, broad stout duct, two (4) or three (1) stalked diverticula attached to lateral sides of duct, each terminating in sausageshaped receptacle, stalks long, kinked with at least one loop closely adherent to spermathecal duct; spermatophores found within ampulla ( Fig. 2I View Fig ). Male sexual system holandric, testes and funnels enclosed in paired sacs in x, xi; seminal vesicles xi, xii each with digitate dorsal lobe; vasa deferentia slender, free from body wall en route to ental end of prostatic ducts; each prostate racemose, occupying xvi, xvii-xx, bilobed, stout muscular duct entering apex of egg-shaped copulatory bursa in xviii; coelomic and exterior surfaces of paired copulatory bursae muscular, lacking glandular or other projections; bulky angular penis present, no pads flanking opening.
Remarks. – Pheretima monoporata keys to P. ambonensis in Sims & Easton (1972), but differs from that species in having very closely spaced male openings, appearing as only one externally, whereas P. ambonensis has two distinct openings about 0.2 circumference apart. In P. monoporata the outer single opening, with its sphincter-like surrounding folds, conceals paired channels leading to the paired copulatory bursae. The ancestral state of two openings has given way to a fusion of the outermost aspects of the two, but there are still independent channels separated by a small distance. The intestinal origin of P. monoporata is in xvii, not xv, the body length is half that of P. ambonensis (120mm), and the setae are about half as numerous. The spermatheca of P. monoporata has longer diverticulum chambers and stalks. The single spermatheca appears to be derived from fusion of a pair of spermathecae, as in P. monotheca and P. ambonensis . The copulatory bursae, rather than the usual broadly attached half-ovals or hemispheres of other Mindanao species encountered so far (except P. alba ; see below), are attached by one apex of the egg-shape. The copulatory bursae lack internal pads, but have penes.
A spermatophore (Thekozyste) was reported from the spermathecae in the Sumatran Metaphire singalangi ( Michaelsen, 1930) , but here the spermatophores have a shorter neck and more rounded body. Thecacysts and other free structures related to sperm exchange were discussed in the context of some aquatic Oligochaeta in Michaelsen (1926; p. 27), but a full characterization of these structures is in Stephenson (1930). In the case of P. monoporata , the structure found in the spermatheca is most probably a true spermatophore formed in the copulatory pouch, because spermatophores are known to occur in P. montana , among other species ( Beddard, 1911).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |