
Pilaridrilus uliginosus, Fend, Steven V. & Lenat, David R., 2007
|
publication ID |
https://doi.org/ 10.5281/zenodo.180092 |
|
DOI |
https://doi.org/10.5281/zenodo.6252647 |
|
persistent identifier |
https://treatment.plazi.org/id/03953B40-FFB7-FFFD-FF35-E28FFC3DCCD0 |
|
treatment provided by |
Plazi |
|
scientific name |
Pilaridrilus uliginosus |
| status |
sp. nov. |
Pilaridrilus uliginosus View in CoL n. sp.
Holotype: USNM 1107791. A sagittally sectioned worm, stained in hematoxylin and eosin, collected by S. Fend and D. Lenat.
Type locality: North Carolina: Halifax Co.: Beech Swamp at SR 1003, N36º 05’ 13”, W77º 20’10”, 17 March 2007.
Paratypes: USNM 1107792-1107793. Collected at the type locality, 17 March 2007. 1 sagittally sectioned, 1 dissected on slide.
Other material: From the type locality, 1 February 2002. 8 dissected on slides.
Etymology: The genus is named after Prof. Pilar Rodriguez (University of the Basque Country, Spain), for her many contributions to lumbriculid systematics. The specific epithet refers to the swampy habitat.
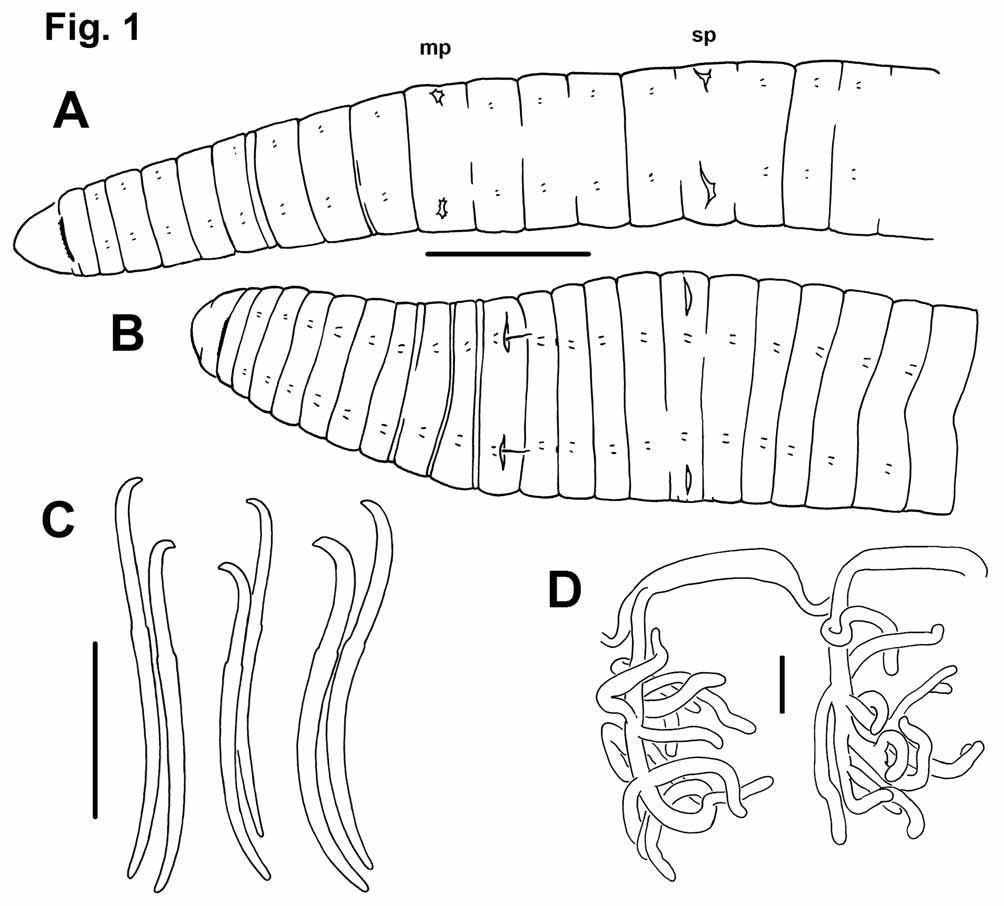
Description. Length of preserved worms 17 to 28 mm; diameter 0.8-1.1 mm in X, maximum diameter to 1.6 mm ( Figs. 1 View FIGURE 1 A–B). Prostomium rounded to nearly conical, as long as wide when fixed in Bouin’s solution ( Fig. 1 View FIGURE 1 A), shorter than wide when preserved in alcohol ( Fig. 1 View FIGURE 1 B). Secondary segmentation weak, limited to a narrow ring in two or three segments anterior to the clitellum.
Chaetae paired, in 4 bundles per segment, beginning in II. Chaetae sigmoid, simple-pointed, with slightly distal nodulus, 0.35–0.40 from the tip ( Fig. 1 View FIGURE 1 C). Chaetal length 180 to 216 Μm; dorsals the same size as or slightly smaller than ventrals. In posterior segments, one chaeta in each pair may be slightly thicker than the other ( Fig. 1 View FIGURE 1 C). Tips of some chaetae may appear slightly truncated, possibly from wear.
Epidermis in anterior segments 20–25 Μm thick; thickest part of clitellum 40–70 Μm. Clitellum distinct in some specimens, thickened and glandular; position variable, from the atrial or pre-atrial segment to at least the spermathecal segment (typically from IX or X to XVI or XVII); in one specimen, epidermis is thin but glandular through XX. Circular muscle of body wall up to 5 Μm thick; longitudinal muscles about 30–40 Μm thick. Pharynx without a distinct dorsal pad; thickened in II–IV dorsally, mid-II or III–IV ventrally. Pharyngeal glands (III)IV–VI.
First pair of nephridia on 6/7. Nephridia absent in VIII–XII, usually paired in XIII, and occurring irregularly in posterior segments. Each nephridium with small anteseptal funnel, a granular postseptal expansion (length 100–130 μm, diameter 30–35 μm), and a long, convoluted duct that may pass through two or more posterior segments ventral to the gut, and terminating in a short duct to a simple nephropore anterior to the ventral chaetae.
Anterior segments with one pair of convoluted commissural blood vessels. One pair of blind, branched lateral vessels joins dorsal vessel in anterior part of each segment posterior to XVIII or XX; usually about 10 branches per vessel ( Fig. 1 View FIGURE 1 D).
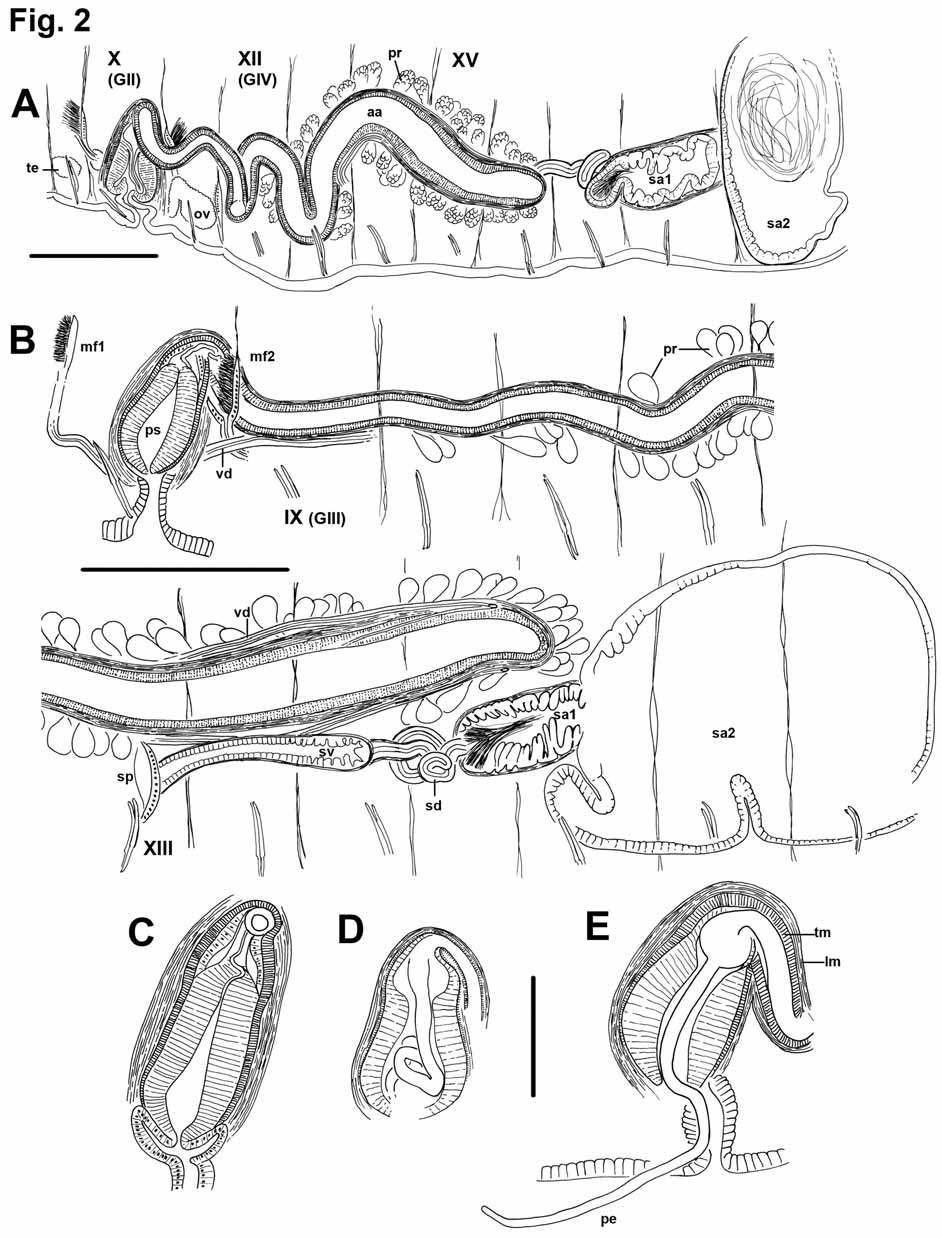
One pair of male pores, usually in X (in VIII in two specimens), posterior to ventral chaetae ( Figs. 1 View FIGURE 1 A–B). Spermathecal pores in shallow, transverse folds slightly lateral to ventral chaetae, in posterior part of XV, near 15/16 (XIII in the worms having male pores in VIII). Female pores paired, on chaetal line, usually at intersegmental groove 11/12. Paired testes usually on anterior septa in IX and X; those in X usually larger. Sperm sacs may extend anteriorly to VII or VI, and posteriorly as far as XXIV. Ovaries usually in XI, but may extend back through XII within egg sacs. Egg sacs may extend as far as XXVI. Female funnels are simple tubes, with posterior lip higher than the anterior.
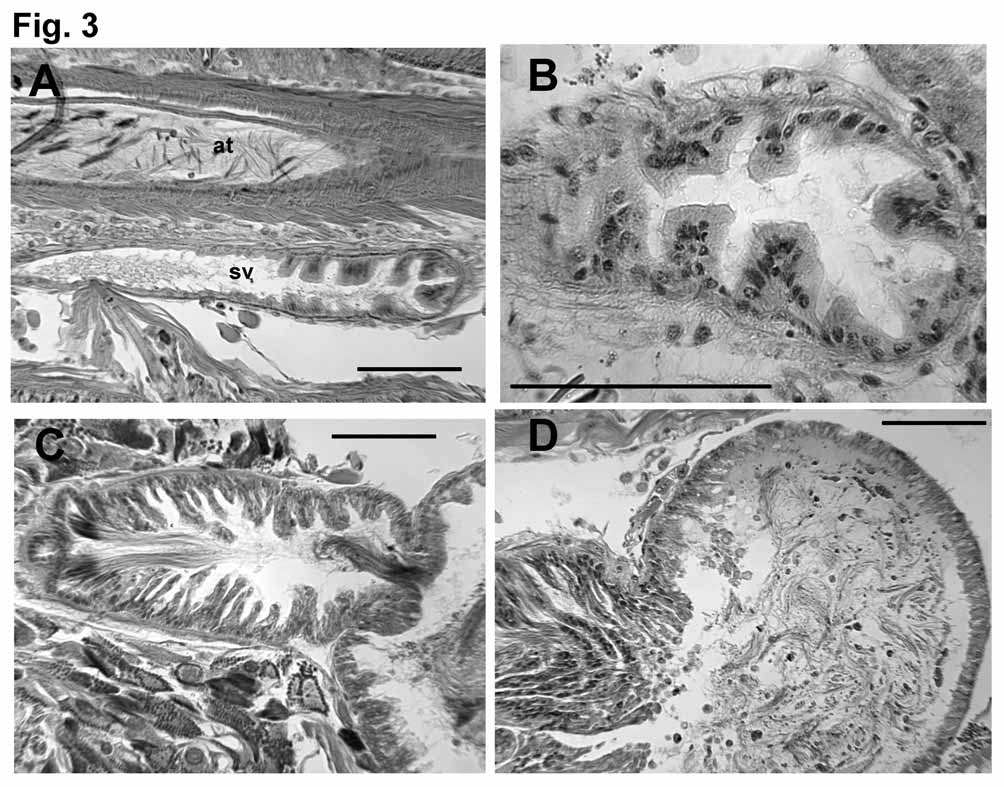
Ectal end (at the pore) of spermathecal duct is widened into a narrow vestibule, 200–500 μm long, and up to 110 μm wide; vestibule may be entirely in XV or extend through one or two segments to as far as 17/18 ( Fig. 2 View FIGURE 2 B, 3A). Epithelium of vestibule becomes uneven in ental half, with clumps of elongated cells separated by deep notches ( Fig. 3 View FIGURE 3 B); a thin (2–5 μm) circular muscle layer, and sparse, irregular outer muscle layer surround the vestibule. Ental to the vestibule, a thin (30–50 μm) tubular section of the spermathecal duct is 450 to 670 μm long, but usually tightly convoluted, forming a compact mass; this section of duct has cuboidal epithelium and a 5–6 μm muscle coat, which constricts it at the junction with the vestibule. Spermathecal ampulla in two parts. Ectal chamber 200–430 long by 160–280 wide, epithelium produced or folded into narrow, multicellular lamellae, which fill the chamber. Sperm heads are lined up along lamellae, concentrated near ectal end ( Figs. 2 View FIGURE 2 A–B, 3C). Ectal chamber surrounded by a circular muscle layer 2–6 μm thick, and an outer longitudinal muscle to 10 μm. Ental chamber (main part) of spermathecal ampulla irregular to ovate, length about 500– 1000 μm, width 500–650 μm; may extend through two segments, to as far as XXI. Epithelial cells somewhat elongate (to 25 μm) and uneven near junction with ectal chamber, but thin (12 μm) and regular entally; cells do not contain large vacuoles. Sperm in ental chamber sparse and unordered, or with heads directed toward wall; usually not darkly-staining ( Fig. 3 View FIGURE 3 D).
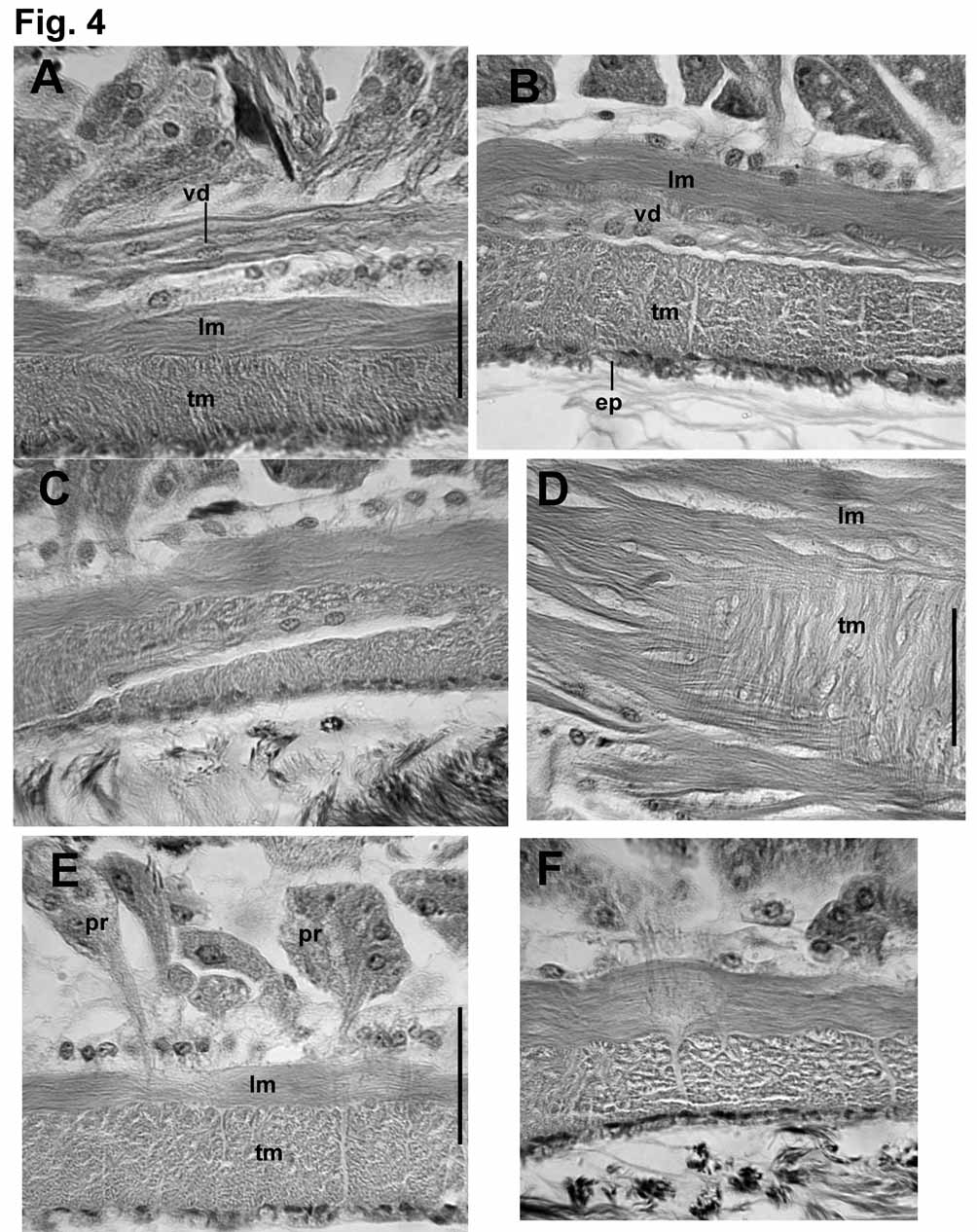
Male funnels on 9/10 and 10/11; the anterior about 160 μm high, flat on septum and directed anteriorly; the posterior may be larger (to 290 μm) than the anterior, with more sperm, and may be convoluted and/or folded back into sperm sac. Vasa deferentia thin (12–18 μm) and difficult to follow, ciliated, and covered by a thin muscle layer (1–2 μm) in the ectal half, near the atrium ( Fig. 4 View FIGURE 4 A). Both vasa run parallel to but separate from atria for most of their length, appressed to the atria in about the ental 1/5, and penetrating the longitudinal muscle near the ental end. Vasa then double back, running through both muscle layers ( Figs. 4 View FIGURE 4 B–C), and apparently enter the atrial lumen near the ental 1/4 ( Figs. 2 View FIGURE 2 B, 4C). The posterior vasa deferentia run directly to the sperm sacs in X, without penetrating 10/11 or forming a loop into XI.
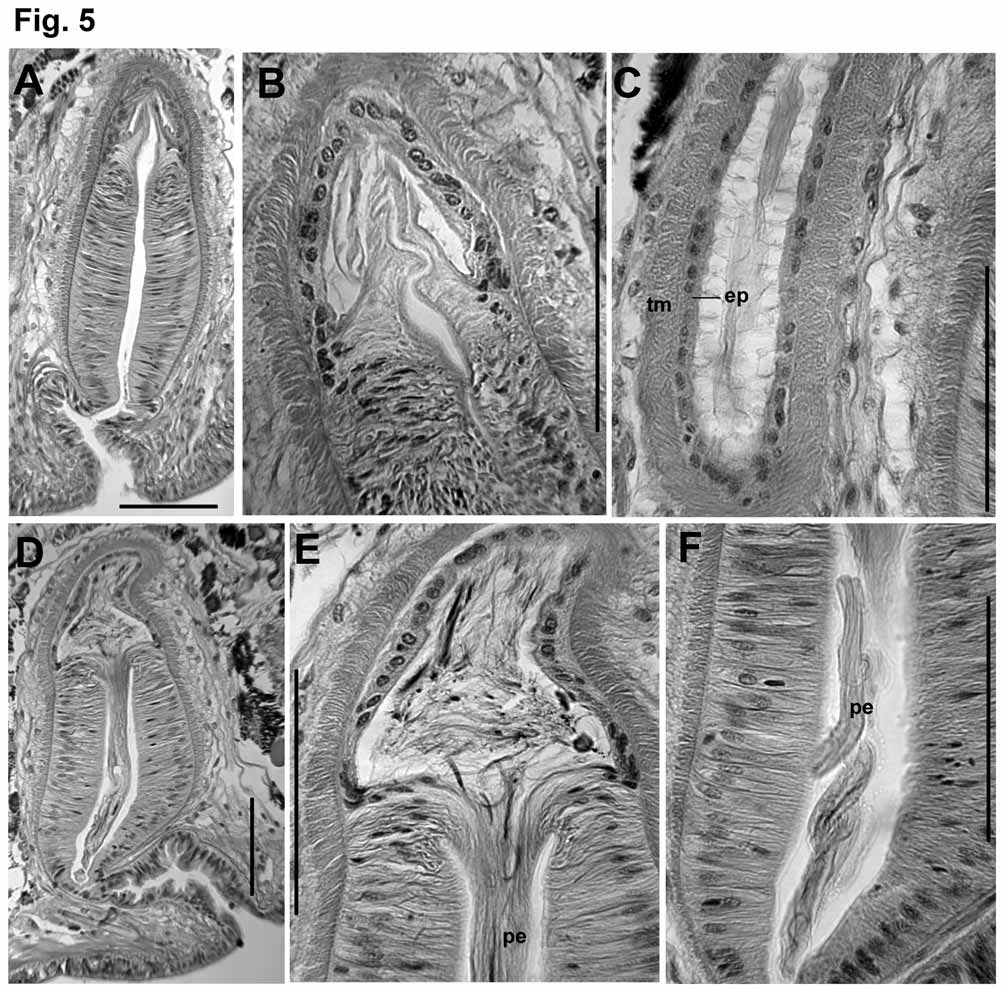
Atria usually pass through several septa, to as far as XIX. Atria elongate-petiolate, with a narrow, tubular ampulla having thickened muscle layers and a dense prostate covering, and a thinner, ectal duct. Total atrium length 1500–3200 μm; width of ampulla 180–250 μm; width of ectal duct 50–110 μm. Atrial muscle in two layers. The inner (transverse-circular) muscle layer has fibers arranged orthogonally to the longitudinal axis; the fibers appear separate and unordered, in contrast to the lamellar arrangement of the penial musculature; thickness of the transverse layer 7–15 μm in the ectal duct, and 20–50 μm in the ampulla. The outer layer is composed of unordered longitudinal muscle fibers; thickness of longitudinal layer 5–14 μm in the ectal duct, 12–25 μm in the ampulla. Epithelium thin but distinct (2–7 μm) in the ectal duct; very thin and inconspicuous in the ampulla. Longitudinal muscle covered with an even, unicellular layer of peritoneum, and densely packed but separate prostate glands. Prostate glands are pyriform-petiolate bundles, each with many cells, 70– 150 Μm long. Prostate cells granular, with indistinct boundaries; conjoined bundles of ducts penetrate longitudinal muscle of atrial ampulla via longitudinal, lenticular openings; then penetrate circular muscle via transverse openings ( Figs. 4 View FIGURE 4 D–F). Transition between the atrial duct and ampulla is gradual; prostate glands may begin by the ectal 1/5 of the atrium, but the thickening of muscle layers and widening of the ampulla begins Penial structures ovate to bell-shaped, length about 350–450 μm, greatest width about 170–230 μm ( Figs. 2 View FIGURE 2 C–E, 5A, 5D). The main part (here termed the “penial chamber”) is a thick tube of narrow, columnar epithelium (30–60 μm thick), surrounding a narrow lumen; the thick epithelial layer is covered by a thin layer of transverse-circular muscle and a loose outer layer of longitudinal muscle and peritoneal cells. In contrast to the atrium, circular muscle fibers are arranged in horizontal lamellae. A very long, whiplike penis may be folded within the penial chamber ( Figs. 2 View FIGURE 2 D, 5D–F) or extend out the male pore ( Figs. 1 View FIGURE 1 B, 2E), but may instead be withdrawn into the base of the penial structure, or into the atrial duct ( Fig. 2 View FIGURE 2 C, 5A–C). Extended penis length in one worm 740 μm (510 μm outside the pore); diameter 24 μm near base within the sac; 18 μm near middle. In another specimen, a partially extended penis is folded twice within the penial chamber, to a total length of 540 μm. Retracted penes, or at least the cuticular lining (diameter about 10 μm), can extend at least 300 (maximum 790) μm entally within the atrial duct, within a network of thin “fibers” ( Figs. 5 View FIGURE 5 C). Ental end of penial structure contains an internal conical to globular chamber, apparently the widened end of the atrial duct ( Figs. 5 View FIGURE 5 B, 5E); this part of the structure subtends the extended penis ( Figs. 2 View FIGURE 2 E, 5D– E), or may contain the coiled, retracted penis. Cells at the transition between atrial epithelium and the columnar epithelium of the penial chamber are very elongate, and apparently extend to form the actual penis ( Fig. 5 View FIGURE 5 E); when the penis is retracted, cell extensions are withdrawn into the terminus of the atrial duct ( Fig. 5 View FIGURE 5 B). Cell nuclei are visible only at the base of the extended penis. Ectally, the penial structure terminates within an irregular fold 100–200 μm deep; the fold is thin-walled and convoluted, and ectally forms the male pore ( Figs. 5 View FIGURE 5 A, 5D).
Remarks. Most of the worms used in the description were contorted and poorly fixed, having been preserved in alcohol. Some characters, such as the unusual path of the vasa deferentia, could not be confirmed in those specimens. The three specimens collected in 2007 were fixed in Bouin’s solution, and form the basis for histological observations. Two specimens, including the dissected paratype ( Fig. 2 View FIGURE 2 B), had the entire reproductive system shifted forward two segments (testes beginning in VII, male pores in VIII; spermathecal pores in XIII). In a single specimen, one of the atria was directed anteriorly, into the anterior sperm sac; in all other cases the atria extended backward into posterior sperm sacs.
The very distinctive placement of spermathecal pores ( Figs. 1 View FIGURE 1 A–B) makes mature P. uliginosus easy to identify as unmounted or whole-mounted specimens. Cleared, immature worms in good condition should also be distinguishable from sympatric taxa on the basis of blood vessels in posterior segments. Immature Pilaridrilus can be distinguished from the morphologically similar Eclipidrilus species by the single pair of branched vessels in posterior segments ( Fig. 1 View FIGURE 1 D), as opposed to two pairs in Eclipidrilus ( Fend 2005) ; the simplepointed chaetae distinguish it from the Nearctic species of Lumbriculus .
| USNM |
Smithsonian Institution, National Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.