
Uroplatus fivehy, Ratsoavina & Glaw & Raselimanana & Rakotoarison & Vieites & Hawlitschek & Vences & Scherz, 2020
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4895.2.5 |
|
publication LSID |
lsid:zoobank.org:pub:F042B4D5-1902-4BE9-A797-00F5AF0659F5 |
|
DOI |
https://doi.org/10.5281/zenodo.4361887 |
|
persistent identifier |
https://treatment.plazi.org/id/AC24C7D3-1AB5-451F-9DE2-935DE7174FDD |
|
taxon LSID |
lsid:zoobank.org:act:AC24C7D3-1AB5-451F-9DE2-935DE7174FDD |
|
treatment provided by |
Plazi |
|
scientific name |
Uroplatus fivehy |
| status |
sp. nov. |
Uroplatus fivehy sp. nov.
LSID: urn:lsid:zoobank.org:act:AC24C7D3-1AB5-451F-9DE2-935DE7174FDD
Figs 3 View FIGURE 3 , 4 View FIGURE 4 , 6–8 View FIGURE 6 View FIGURE 7 View FIGURE 8
Remarks. This new species corresponds to the lineage previously named Uroplatus sp. A by Raxworthy et al. (2008), Uroplatus sp. 4 by Ratsoavina et al. (2011), and Uroplatus ebenaui [Ca4] by Ratsoavina et al. (2012, 2013, 2015, 2017, 2019a,b).
Holotype. ZSM 1721 View Materials /2012 (field number FGZC 3590 ), adult male, collected on 26 November 2012 by F. Glaw, O. Hawlitschek, T. Rajoafiarison, A. Rakotoarison, F. M. Ratsoavina and A. Razafimanantsoa on the Sorata Massif , Sava Region, northern Madagascar, in the vicinity of the campsite, near 13.6805°S, 49.4451°E, 1417 m a.s.l. GoogleMaps
Paratypes. A total of 12 specimens: ZSM 1722 View Materials /2012 (field number FGZC 3591 ), adult female, with same collection data as the holotype GoogleMaps ; UADBA-R 70851 ( FGZC 3694 ), adult male, collected by same collectors as holotype on 29 November 2012 on the Sorata Massif at 13.6868°S, 49.4412°E, 1272 m a.s.l. GoogleMaps ; UADBA-R 70850 ( FGZC 3715 ), adult male, collected by same collectors as holotype on 30 November 2012 on the Sorata Massif near a creek at 13.6829°S, 49.4403°E, 1325 a.s.l GoogleMaps .; ZSM 634 View Materials /2009 ( ZCMV 11308 ), adult male, and UADBA-R 70966 ( ZCMV 11309 ), adult female, collected between 22–25 June 2009 by F. Ratsoavina , R. D. Randrianiaina, E. Rajeriarison , T. Rajoafiarison, D . R. Vieites and M. Vences near Angozongahy campsite on the western side of the Makira plateau in the Northern Central East of Madagascar, near 15.437°S, 49.1167°E, 1009 m a.s.l. GoogleMaps ; UADBA 29428 View Materials ( APR 05955) and UADBA 29427 View Materials ( APR 05921), two adult males collected on 22 November 2004 and UADBA 29426 View Materials ( APR 05859), subadult, collected on 19 November 2004 by A.P. Raselimanana at Anjiabolo site (ca. 16.2800°S, 48.8017°E; respectively at 1100 m and 1000 m a.s.l) in Marotandrano Special Reserve , Sofia Region, Northern Central East Madagascar GoogleMaps ; UADBA 29430 View Materials ( APR 05984), UADBA 29431 View Materials ( APR 05985), two males and UADBA 29429 View Materials ( APR 05983), adult female, collected on 25 November 2004; and UADBA 29432 View Materials ( APR 06118), adult male, collected on 29 November 2004 by A.P. Raselimanana at Riamalandy site (16.2850°S, 48.8150°E, between 825–875 m a.s.l.) in Marotandrano Special Reserve GoogleMaps .
Etymology. The species epithet fivehy is a noun in apposition to the genus name, meaning “paddle” in local Malagasy dialect, and referring to the paddle-like shape of its tail.
Diagnosis. Uroplatus fivehy sp. nov. is assigned to the Uroplatus ebenaui species group based on its relatively small size, leaf-like, laterally-compressed body shape, short tail, and triangular head with supraocular spines. The following unique combination of characters characterizes the new species: (1) short (TAL/SVL 0.21–0.41) and slender (TAW/SVL 0.06–0.13) tail, (2) oral mucosa without black pigment, and (3) relatively smooth (not spiny) appearance in males. Additionally, it is distinguished by high genetic pairwise-distances> 10% in DNA sequences of the mitochondrial 16S gene from all other nominal species of Uroplatus .
By its overall leaf-like appearance, the species can easily be distinguished from all members of the U. fimbriatus species group, from U. lineatus , and from the phenetic U. alluaudi species group. Within the U. ebenaui species group, U. fivehy sp. nov. can be distinguished from U. finaritra and U. malama by smaller body size (SVL 60.8–72.4 mm versus 77.5–95.3 mm); from U. malama , U. phantasticus , U. finiavana and U. finaritra by shorter tail (TAL/SVL 0.21–0.41 versus 0.48–0.76); from U. fangorn , U. ebenaui , U. fetsy , and U. kelirambo by its unpigmented oral mucosa (pale pinkish all over versus black or deep red pigment present); from U. kelirambo by more expanded tail edges (versus strongly spear-shaped; TAW/SVL 0.06–0.13 versus 0.04–0.05 in U. kelirambo ). U. fivehy sp. nov. is morphologically most similar to U. fiera and to U. fotsivava , which however concordantly differ in mitochondrial and nuclear DNA sequences and are not its immediate relatives based on molecular phylogenetic analysis; it differs from U. fotsivava by a generally longer and wider tail, although the values do overlap (TAL/SVL 0.21–0.41 versus 0.15–0.32; TAW/SVL 0.06–0.13 versus 0.05–0.10), and from U. fiera possibly in a slightly larger ratio TAL/TAW (0.22–0.37 versus 0.19–0.21).
Description of the holotype. Adult male in good condition with an intact tail and everted hemipenes. SVL 63.1 mm, tail length 25.9 mm, maximum tail width 5.7 mm, for further measurements see Table 1 View TABLE 1 . Head triangular in dorsal view; canthus rostralis recognizable and almost straight; snout sloping strongly and continuously downward anteriorly; snout weakly depressed, short; eyes large (eye diameter 4.6 mm), not bulging above dorsal surface of cranium, directed laterally, pupil vertical with crenate borders; ear opening very small but distinct (horizontal diameter 0.6 mm), its opening facing posterolaterally, but also posteroventrally (ear opening clearly visible in lateral view but not in dorsal view); nostrils laterally oriented; body somewhat laterally compressed, without lateral dermal fringes; limbs without fringes and practically without spines on forelimbs and hindlimbs, except for a small flap on the knee; forelimb reaches beyond tip of snout when adpressed forward and does not reach the groin when adpressed backwards along body (forelimb length/axilla–groin distance 27.7/ 29.7 mm = 93%), hindlimb reaches beyond axilla when adpressed forward along body (hindlimb length/axilla–groin distance 37.4/ 29.7 mm = 126%); original tail length 41% of snout–vent length, membranous borders of the denticulated tail rather asymmetrical, broadening on either side and then narrowing again toward the tip), completely absent from the slightly spatulate tail-tip.
Nares separated from each other by eight small granular scales, from the first supralabial by one scale, and from the rostral scale by two scales; the first supralabial taller than the others; rostral entire, much wider than tall; mental scale small, not differentiated from infralabial scales; 21/20 (right/left) supralabials and 19/21 infralabials (as defined in methods above); scales at the periphery of the chin (toward the infralabial scales) slightly larger than central chin scales; the dorsal scales of head, neck, body, limbs and tail small, granular, juxtaposed and largely of homogeneous size interspersed with very few larger and partly raised tubercles; the numerous curved transverse lines on the head and dorsal side of the body are not marked by significantly enlarged scales. Very few spines on head and hindlimbs, a dermal flap on each knee, bearing no spine; no flaps or spines on elbow and forelimb; a prominent pointed flap on the posterior portion of each upper eyelid (supraocular spine); posterior border of eye fringed. Axillary pits not recognizable. Scales of the ventral abdomen distinctly larger than dorsal body scales and arranged more homogeneously.
Hemipenes everted, bearing two lobes. Calyx with protuberance bearing honeycomb-like structures, especially on the asulcal side. Area of sulcus spermaticus is smooth. Each lobe with a dense field of pointed papillae at its apex.
Coloration of the holotype. After eight years of preservation in 70% ethanol the color pattern remains similar to that of the living animal ( Figs 6H View FIGURE 6 , 8 View FIGURE 8 ) but its vividness and intensity have slightly faded. All dorsal surfaces are mottled beige and blackish in base color. The tail has a beige pattern on its dorsal pygal section, bordered by blackish color. The dorsum has rather distinctly defined posteroventrally oriented lighter and darker beige chevrons. The dorsal surfaces of the head are dark brown with a series of six distinct transverse narrow beige lines, running either straight or converging anteriorly or posteriorly. No dark band between the eyes, but a dark V-shaped area on the nape is well distinguishable. Additional dark flecks are present on the mid-dorsum, and several smaller asymmetrical spots on other parts of the body, including legs and nape. One light cream marking is present posteriorly below each eye, descending to the supralabials. The supralabials and the infralabials are generally dark gray in color. The venter, including the ventral limbs except for the lower hindlimbs, is slightly lighter beige than the dorsum, and less mottled. The pectoral region bears two elongated, poorly delimited whitish spots. The postpygal section of the tail is of similar color dorsally and ventrally. The oral mucosa is unpigmented and whitish in preservative. The iris of the eye is gray, and the vertical pupil is blackish.
Variation. As described for U. fangorn above, there is a substantial color variation ( Figs 6–7 View FIGURE 6 View FIGURE 7 ), and sexual dimorphism in the edges of the tail. The color of the eye varies from beige/gray to dark brown and reddish-brown. Dorsal color can be uniform beige to reddish-brown with or without white lichen-like markings. Two sharply delimited vertebral black patches on the anterior and posterior part of the dorsum are present in several individuals. Relative tail length and tail width are characterized by substantial variability among individuals ( Fig. 4 View FIGURE 4 ).
Here, we have included in the paratype series and in the morphological comparisons a series of individuals from Marotandrano Special Reserve that we attribute to this species. However, it must be stressed that the species identity has been confirmed by DNA sequences only for one of these (UADBA 29428 = APR5955). In the unlikely case that some of these individuals will turn out to belong to another species of Uroplatus , the morphometric variation of U. fivehy may be narrower than reported here.
Distribution, natural history and conservation status. According to the material collected and examined by us, U. fivehy is known from (1) the type locality on the Sorata Massif, (2) the western slope of the Makira Reserve, and (3) Marotandrano Special Reserve. In addition, mitochondrial DNA sequences published by Ratsoavina et al. (2012) suggest its occurrence in (4) the Anjanaharibe-Sud Massif, and mitochondrial DNA sequences from Raxworthy et al. (2008) furthermore suggest the occurrence of the species in (5) Marojejy, (6) Ankitsika and (7) Betaolana. The confirmed elevational range from our own samples is between 1009 and 1417 m a.s.l., and the record from Ankitsika would extend this range downwards to 830 m a.s.l., characterizing the species as rather wide-ranging from the Northern Central East to the North East of Madagascar from mid-elevations to highlands (see also Ratsoavina et al. 2013). All specimens were found in intact rainforest, at night, perched on branches of trees.
At Marotandrano, an adult male was found hidden in a Pandanus leaf axil during the day in closed canopy humid forest, and two adult individuals (male and female) were found at night close to each other in the same tree on 25 November 2004, suggesting it was probably the reproduction period for this species. Individuals were often observed active on small branches, leaves, and lianas at 0.5–3 m above the forest floor. All individuals were collected on slope or ridge forest. In Marotandrano, the species appeared to be quite common in forests associated with vine-like bamboo and shrub vegetation.
A minimum convex polygon of the known distribution of this species covers an area of ca 15,100 km 2, qualifying the species for a status of Vulnerable under the IUCN Red List criterion B ( IUCN 2012). This species is known from several protected areas (Marotandrano Special Reserve, Makira Reserve, COMATSA Nord, Marojejy National Park, Anjanaharibe-Sud National Park) but cumulatively just seven locations (B subcriterion a). These locations, and especially the unprotected areas between them, are experiencing substantial ongoing declines in the extent and quality of habitat (B subcriterion b(iii)). Therefore, we propose to list this species as Vulnerable under IUCN criterion B1ab(iii).
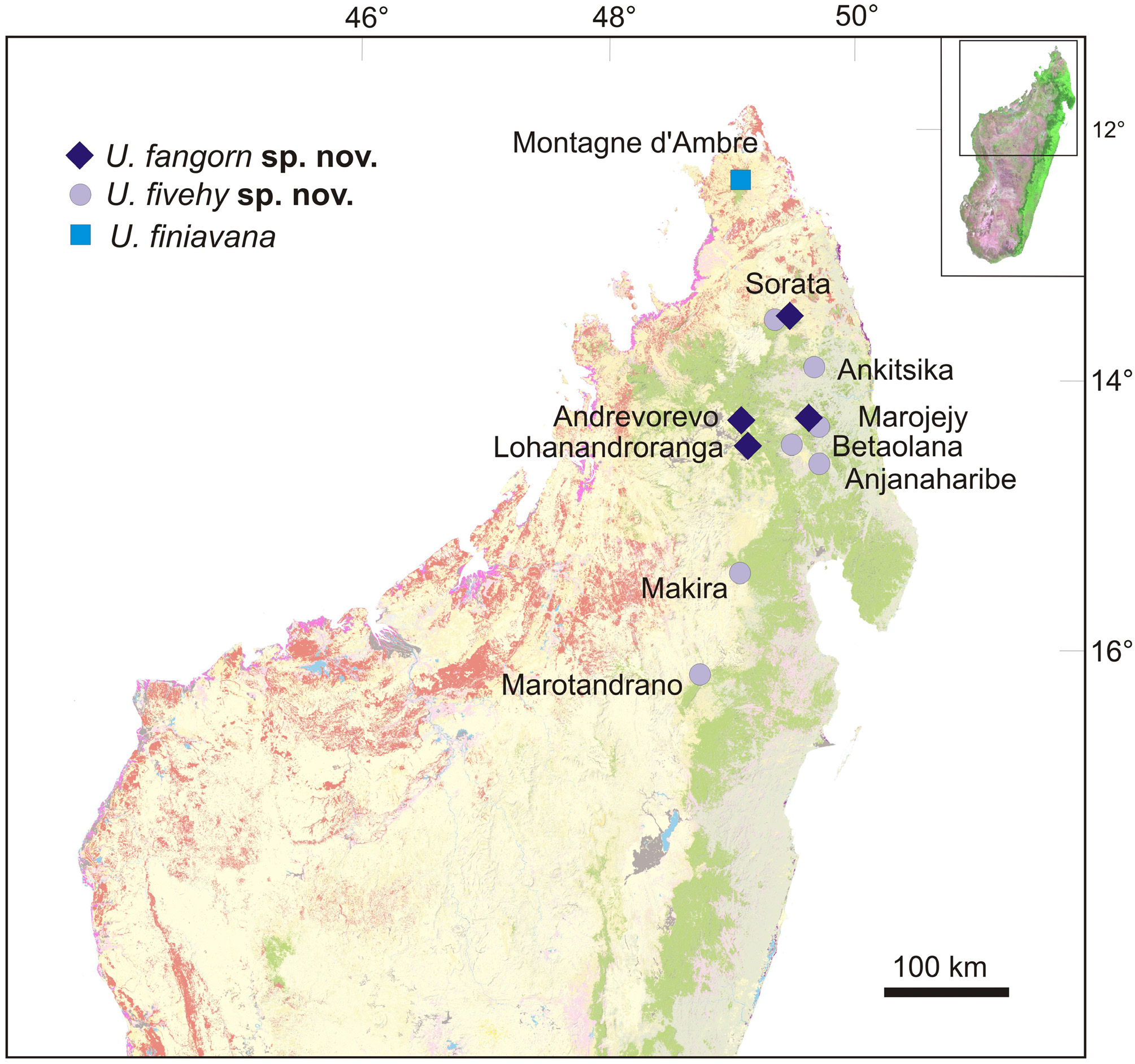




TABLE 1. Measurements in mm, and one scale count (internarial scales, INS) taken from Uroplatus fangorn sp. nov. and U. fivehy sp. nov., and their closest relative, U. finiavana. HT, holotype; PT, paratype; NM, not measured (for tail measurements, referring to specimens with missing or regenerated tail). See Materials and Methods for other abbrevia- tions.
| Locality | Sex | Type status | Museum number | Field number | SVL | TAL | TAW | HL2 | HL | HW | FORL | HIL | INS |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| U. finiavana | |||||||||||||
| Mt. d’Ambre | male | HT | ZSM 328/2004 | FGZC 625 | 59.0 | 35.3 | 5.7 | 18.1 | 19.2 | 13.7 | 25.5 | 34.5 | NM |
| Mt. d’Ambre | male | PT | ZSM 326/2004 | FGZC 623 | 52.0 | NM | NM | 15.8 | 16.3 | 11.8 | 22.1 | 32.7 | NM |
| Mt. d’Ambre | male | PT | ZSM 327/2004 | FGZC 624 | 53.8 | 35.1 | 7.6 | 16.4 | 16.9 | 12.4 | 23.7 | 27.3 | NM |
| Mt. d’Ambre | male | PT | ZSM 329/2004 | FGZC 626 | 60.0 | NM | NM | 17.4 | 18.3 | 13.5 | 24.1 | 33.4 | NM |
| Mt. d’Ambre | male | PT | ZSM 1132/2003 | FGMV 2002.2387 | 61.0 | NM | NM | 17.8 | 18.8 | 13.2 | 27.0 | 35.8 | NM |
| Mt. d’Ambre | male | PT | ZSM 1135/2003 | FGMV 2002.3098 | 58.0 | NM | NM | 17.2 | 18.8 | 13.1 | 26.1 | 34.6 | NM |
| Mt. d’Ambre | male | ZSM 205/2018 | MSZC 0443 | 61.5 | NM | NM | 16.5 | 18.5 | 13.6 | 25.4 | 34.7 | 8 | |
| Mt. d’Ambre | male | ZSM 206/2018 | MSZC 0483 | 43.4 | 22.9 | 6.3 | 12.4 | 14.3 | 10.1 | 17.8 | 23.3 | 10 | |
| Mt. d’Ambre | male | ZSM 207/2018 | MSZC 0696 | 55.6 | NM | NM | 15.1 | 17.1 | 11.8 | 24.6 | 30.0 | 9 | |
| Mt. d’Ambre | male | ZSM 208/2018 | MSZC 0731 | 62.6 | 29.5 | 10.0 | 16.0 | 19.4 | 13.9 | 25.3 | 29.5 | 8 | |
| Mt. d’Ambre | female | PT | ZSM 1134/2003 | FGMV 2002.2390 | 52.0 | 24.8 | 5.6 | 15.1 | 15.2 | 11.4 | 22.4 | 30.1 | NM |
| Mt. d’Ambre | female | PT | ZSM 322/2004 | FGZC 619 | 65.2 | NM | NM | 18.2 | 19.2 | 13.4 | 28.4 | 36.0 | NM |
| Mt. d’Ambre | female | PT | ZSM 323/2004 | FGZC 620 | 56.0 | 29.3 | 7.4 | 16.5 | 17.1 | 13.5 | 23.7 | 33.5 | NM |
| Mt. d’Ambre | female | PT | ZSM 324/2004 | FGZC 621 | 62.6 | 32.3 | 6.6 | 18.0 | 19.1 | 13.5 | 23.5 | 36.1 | NM |
| Mt. d’Ambre | female | PT | ZSM 325/2004 | FGZC 622 | 54.7 | 23.1 | 7.0 | 15.3 | 16.3 | 12.0 | 21.0 | 32.4 | NM |
| U. fangorn sp. nov. | |||||||||||||
| Marojejy | male | HT | ZSM 79/2005 | ZCMV 2030 | 62.5 | 19.8 | 4.9 | 16.2 | 18.0 | 13.1 | 25.5 | 33.8 | 7 |
| Andrevorevo | male | PT | ZSM 644/2014 | DRV 6280 | 59.5 | 25.6 | 4.2 | 16.2 | 17.7 | 13.7 | 25.5 | 33.5 | 6 |
| Andrevorevo | female | PT | UADBA 70054 | DRV 6281 | 63.9 | 17.6 | 5.2 | 12.0 | 18.2 | 14.8 | 24.8 | 34.2 | 6 |
......continued on the next page
| R |
Departamento de Geologia, Universidad de Chile |
| T |
Tavera, Department of Geology and Geophysics |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.