
Opisthotropis andersonii ( Boulenger, 1888 )
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4247.4.3 |
|
publication LSID |
lsid:zoobank.org:pub:B926664F-8D4E-4CBA-8EEA-317B5AB46357 |
|
DOI |
https://doi.org/10.5281/zenodo.5674721 |
|
persistent identifier |
https://treatment.plazi.org/id/1B666942-FFDF-FF97-FF59-FB31FBBC56A5 |
|
treatment provided by |
Plazi |
|
scientific name |
Opisthotropis andersonii ( Boulenger, 1888 ) |
| status |
|
Opisthotropis andersonii ( Boulenger, 1888)
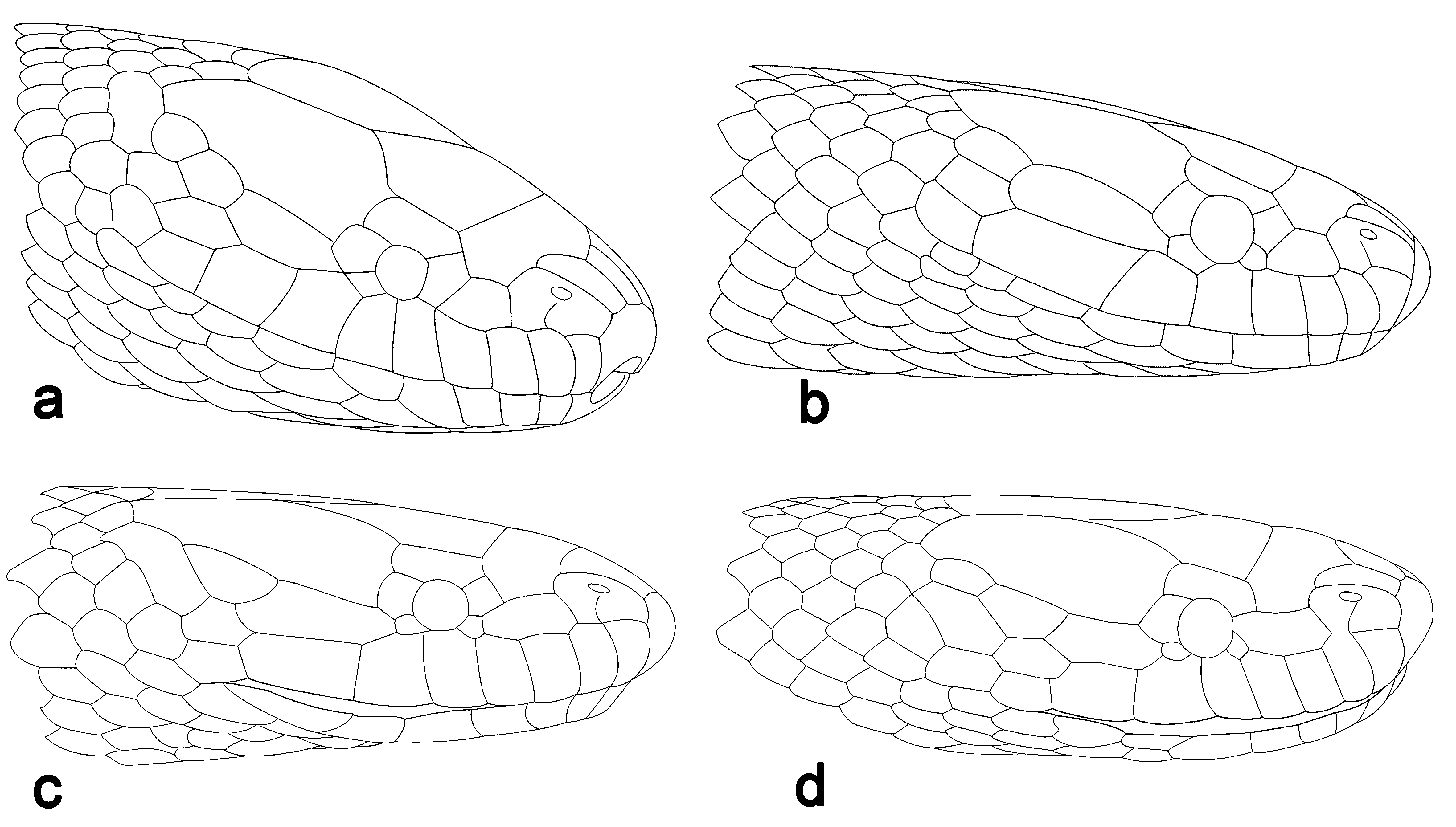
Figs. 4 View FIGURE 4 and 6 View FIGURE 6 c, d
Calamohydrus Andersonii Boulenger, 1888 . Type locality: Hong Kong. Opisthotropis andersonii: Boulenger 1893
Opisthotropis andersoni: Smith 1943
Opisthotropis andersonii: Zheng 1992 ; Zhao et al. 1998; Yang et al. 2013
Specimens examined. Ten specimens from China: SYS r001423, adult female, collected by Anthony Lau on 29 January, 2015, from Tai Tam (22°15′32.52″N, 114°12′6.6″E; 252 m a.s.l.) GoogleMaps ; SYS r001424, adult female, collected by Yik-Hei Sung on 20 May, 2015, from Tai Mo Shan (22°24′36.18″N, 114°7′12.12″E; 740 m a.s.l.) GoogleMaps . SYS r000607, adult male, collected by Jian-Huan Yang (JHY hereafter) and Run-Lin Li (RLL hereafter) on 8 March, 2012; SYS r001017, juvenile, and SYS r001020, adult female, collected by JZ and RLL on 15 and 16 September, 2014; SYS r001082, adult male, collected on 13 March 2015 by JW and ZTL, all from Mt. Wutong (22°34′54.8″N, 114°12′2.7″E; 250–300 m a.s.l.) GoogleMaps . SYS r000803, adult male, collected by JZ and YLL on 5 April, 2013 from Mt. Yangtai (22°38′36.09″N, 113°59′7.45″E; 150 m a.s.l.), Shenzhen City , Guangdong Province GoogleMaps . SYS r000893, adult male, collected by JZ and YLL on 3 November, 2013 from Dapeng Peninsula (22°30′17.55″N, 114°32′23.01″E; 102 m a.s.l.), Shenzhen City , Guangdong Province GoogleMaps . SYS r001382, an adult female, and SYS r001383, a juvenile, by JW on 16 October, 2015 from Mt. Maofeng (23°18′22.79″N, 113°22′58.03″E; 55 m a.s.l.). Further information of collection localities is listed in Table 1 View TABLE 1 . GoogleMaps
Description. Measurements, scalation and body proportions of eight adult specimens are listed in Table 3. The examined specimens roughly conform to the previous descriptions ( Boulenger 1888; Zhao et al. 1998), and show significant variation. Body cylindrical; adult specimens measuring 378–462 mm in TL; tail short, TaL of 15–20% of TL; Head small, indistinct from neck; rostral concave, visible from above, slightly enlarged, more than twice as broad as deep, RW 31–37% of HW; nasal directed dorsolaterally, rectangular or pentagonal, in contact with first two supralabials ventrally, with loreal and prefrontal posteriorly, with internasal dorsally, with rostral anteriorly; a short vertical cleft below the nostril and dividing nasal into anterior and posterior parts, pointing to the posterior upper-angle of first supralabial (n=3), to the anterior upper-angle of second supralabial (n=3), or connecting with the suture between first and second supralabials (n=4); a single loreal, more than twice as long as deep (LoL/LoD 2.1–2.9), not in contact with internasal, in contact with second, third and fourth supralabials (in contact with second, third, fourth and fifth in left side of head of SYS r000803), whereas only in contact with third and fourth supralabials in original description ( Boulenger 1888); in specimens SYS r001423, SYS r001020, 0 893 and 0 607, loreal not entering the orbit, preoculars 2–3 (3 only on right side in SYS r001423), upper one significantly larger than the lower ones; in SYS r001424, SYS r000803, 1382, 1082, 1017 and 1383, loreal entering the orbit, preocular absent in SYS r000803, 1382 and 1383, present a small preocular under the loreal in SYS r001424, SYS r001082 and SYS r001017; postoculars 1–2; anterior temporal single, significantly prolonged (anterior temporal broken into three in specimen SYS r001423); posterior temporals two, only one on left side of head in specimen SYS r001423; supralabials 8/ 8 in all adults, 7/ 7 in two juveniles, commonly first three rectangular, the last one significantly smaller than the adjacent preceding supralabial; the fifth, or fourth and fifth, or fourth, fifth and sixth supralabials touching the orbit; 7–10 infralabials, the first one in contact with its fellow behind the mental; two pairs of chin shields; anterior chin-shields large, in contact with each other, and in contact with the first four or first five infralabials on both sides; posterior chin shields small and separated by small scales; dorsal scale rows 17:17:17; dorsal scales weakly keeled throughout the body in previous description ( Zhao et al. 1998; Teynie et al. 2014), whereas smooth on the anterior neck in our specimens, first presenting barely perceptible weak keel on 10th vertebral scale in SYS r001423, on 13th in SYS r001424, 15th in SYS r000803 and 1382, 17th in SYS r001020, 18th in SYS r001082, 21st in SYS r000607, 30th in SYS r001017, 46th in SYS r001383, 58th in SYS r000893, and gradually visible posteriorly; dorsal scales moderately and visibly keeled at mid-body to posterior part of body, the outermost row smooth; dorsal scales rather strongly keeled on tail; V 141–174; cloacal plate divided; subcaudals 43–59, paired.
Coloration in life. Dorsal surfaces of head, body and tail olive-green to olive-brown, usually each scale with black lateral edges, forming distinct or barely perceptible longitudinal black stripes parallel to each other, crossing each scale ( Fig. 4 View FIGURE 4 ); sometimes dorsal scales with light-colored posterior edge; the most outer a scale row yellow, usually with black upper edge; mental and infralabials black-brown; venter yellow in adult specimens, yellowishwhite in juvenile specimens; chin shields, throat, subcaudals usually with black markings; in SYS r000807, each ventral scale with a middle black spot, formed a longitudinal row along medioventral line.
Coloration in ethanol. Dorsal surfaces blackish-brown, longitudinal stripes indistinct or even not visible; dorsal scales more or less with pale-colored posterior edges; pale-yellow to whitish beneath, more or less with grey black spots.
Revision of diagnosis. This species is characterized by the combination of (1) TL 378–462 mm in adult specimens; (2) TaL 15–20% of TL; (3) rostral concave, slightly large, RW 31–37% of HW; (4) nasal in contact with first and second supralabials; (5) loreal more than twice as long as deep; (6) loreal entering orbit or not, in contact with second to fourth or to fifth supralabials; (7) supralabials 7–8, the last one significantly smaller than the adjacent preceding one; (8) infralabials 7–10; (9) dorsal scale rows 17:17:17; (10) dorsal scales smooth on the neck, feebly keeled on the middle part of body, rather strongly keeled on posterior part of body, strongly keeled on the tail; (11) ventrals 141–174; subcaudals 43–60; (12) olive-green to olive-brown above, with distinct or barely perceptible longitudinal black stripes parallel to each other, crossing each scale; yellow beneath, with black markings in life.
Distribution and habitat. Opisthotropis andersonii has been recorded in Hong Kong and adjacent Shenzhen City, extending north to Guangzhou City, China, and in northern and central Vietnam ( David et al. 2011). It is a low mountain dweller, commonly found in montane fast or slow-flowing streams, tiniest trickles covered with bare rocks, pools and ditches, surrounded by moist subtropical evergreen broadleaved forests.
Remarks. Opisthotropis andersonii was described originally from a single juvenile specimen with TL 240 mm, TaL 45 mm, which was obtained, through the kind mediation of Dr. J. Anderson from the Directors of the City Hall Museum, Hong Kong, labelled as from Hong Kong, presented to the City Hall Museum by C. Ford ( Boulenger 1888). As the City Hall Museum has been closed for a long time and we have been unable to examine this preserved specimen.
According to the original description of Boulenger (1888), O. andersonii possessed a preocular, two suboculars and a postocular. Zheng (1992) examined two specimens (BM.217 and 218, collected on 1956) deposited in the British Museum, London and considered that O. andersonii possessed two preoculars and two postoculars, no subocular. Obviously, Zheng (1992) treated two suboculars (i.e., pre-subocular and post-subocular) considered by Boulenger (1888) as lower preocular and lower postocular, respectively. We follow the descriptions of Zheng (1992). Opisthotropis andersonii is oviparous, probably feeds on semi-aquatic earthworms, and sometimes does well in captivity on a diet of earthworms ( Karsen et al. 1998).

| SYS |
Zhongshan (Sun Yatsen) University |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Opisthotropis andersonii ( Boulenger, 1888 )
| Wang, Ying-Yong, Guo, Qiang, Liu, Zu-Yao, Lyu, Zhi-Tong, Wang, Jian, Luo, Lin, Sun, Yan-Jun & Zhang, Yan-Wu 2017 |
Opisthotropis andersonii:
| Zheng 1992 |
Opisthotropis andersoni:
| Smith 1943 |
Opisthotropis andersonii:
| Boulenger 1893 |
Calamohydrus
| Andersonii Boulenger 1888 |