
Magelona minuta Eliason, 1962
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4527.4.5 |
|
publication LSID |
lsid:zoobank.org:pub:B6EE38BD-7333-4013-B9CA-111A406BCFC5 |
|
DOI |
https://doi.org/10.5281/zenodo.5980871 |
|
persistent identifier |
https://treatment.plazi.org/id/03B8C123-ED59-E123-FF75-3091FB3BA30C |
|
treatment provided by |
Plazi |
|
scientific name |
Magelona minuta Eliason, 1962 |
| status |
|
Magelona minuta Eliason, 1962 View in CoL
Figures 2–10 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 View FIGURE 8 View FIGURE 9 View FIGURE 10
Magelona rosea: Southern (1914) View in CoL west Ireland (see Eliason, 1962)
Magelona rosea: Eliason (1920) View in CoL west Sweden (see Eliason, 1962)
Magelona papillicornis var. rosea: Thorson (1946) View in CoL Denmark (see Eliason, 1962)
Magelona rosea: McIntyre (1958) View in CoL ? (see discussion re M. filiformis minuta View in CoL )
Material examined. Holotype: Sweden, Öresund — Station 0 68 69, mud, a little sand, shells, 16 m ( NHMG Polych. 11491; af, f), 12/4/1961.
Non-type material: Hornbaek Bay, Northwestern part of Sound, Öresund, sandy bottom, 15–18 m (USNM 52510; 2af, 1 of which is dissected and slide mounted onto 4 slides (majority of specimen dissected into three fragments and mounted on slide 4: fragment 1—prostomium and 8 thoracic chaetigers; fragment 2—26 abdominal chaetigers, and fragment 3—8 posterior-most abdominal chaetigers. Individual parapodia dissected from specimen and mounted on slides 1–3 as follows: left-hand parapodia of chaetigers 1–8, 9 th and 10 th chaetiger (both sides), right-hand parapodia of chaetiger 16 and left-hand parapodia of chaetiger 35, see Fig. 2 View FIGURE 2 ), collected by K Muus.
Irish Sea, Cardigan Bay — Station 46 (52 o 19.2’N, 0 4 o 37.0’W), sandy gravel, 30 m (NMW.Z.1991.075.1583; c), collected by NMW, 01/08/1991; Aberporth — Station 47 (52 o 09.6’N, 0 4 o 32.5’W), muddy sand, 15 m (NMW.Z.1991.075.1584; 16c, 5af), collected by NMW, 01/08/1991. Celtic Sea, Celtic Deep — Station 59 (51 o 32.0’N, 0 5 o 56.5’W), sand, 109 m (NMW.Z.1991.075.1587; c), collected by NMW, 03/08/1991; Station 60 (51 o 15.8’N, 0 5 o 59.8’W), muddy sand, 93 m (NMW.Z.1991.075.1589; 4c, 5af), collected by NMW, 03/08/1991; Station 40 (51 o 21.245’N, 0 6 o 23.988’W), sandy mud, 123.2 m (NMW.Z.2005.014.0110, af; NMW.Z.2005.014.0111, af, c; NMW.Z.2005.014.0108, af), collected by NMW, 27/07/2005; Station 42 (51 o 21.899’N, 0 6 o 16.877’W), sandy mud, 105 m (NMW.Z.2005.014.0117, c, 2af; NMW.Z.2005.014.0122; c), collected by NMW, 27/07/2005. South Devon, Dart Estuary — Station D5 (~ 50° 21’N, 03°35’W), (NMW.Z.2003.013.0090, af; NMW.Z.2003.013.0091, 2af), collected by D Levell, 03/1987. Scotland, Northern North Sea, Tern Oilfield — Station 6000 (61 o 18’N, 0 0 o 54’E), 160 m (NMW.Z.1987.067.846; af), collected by IOE, 08/1987.
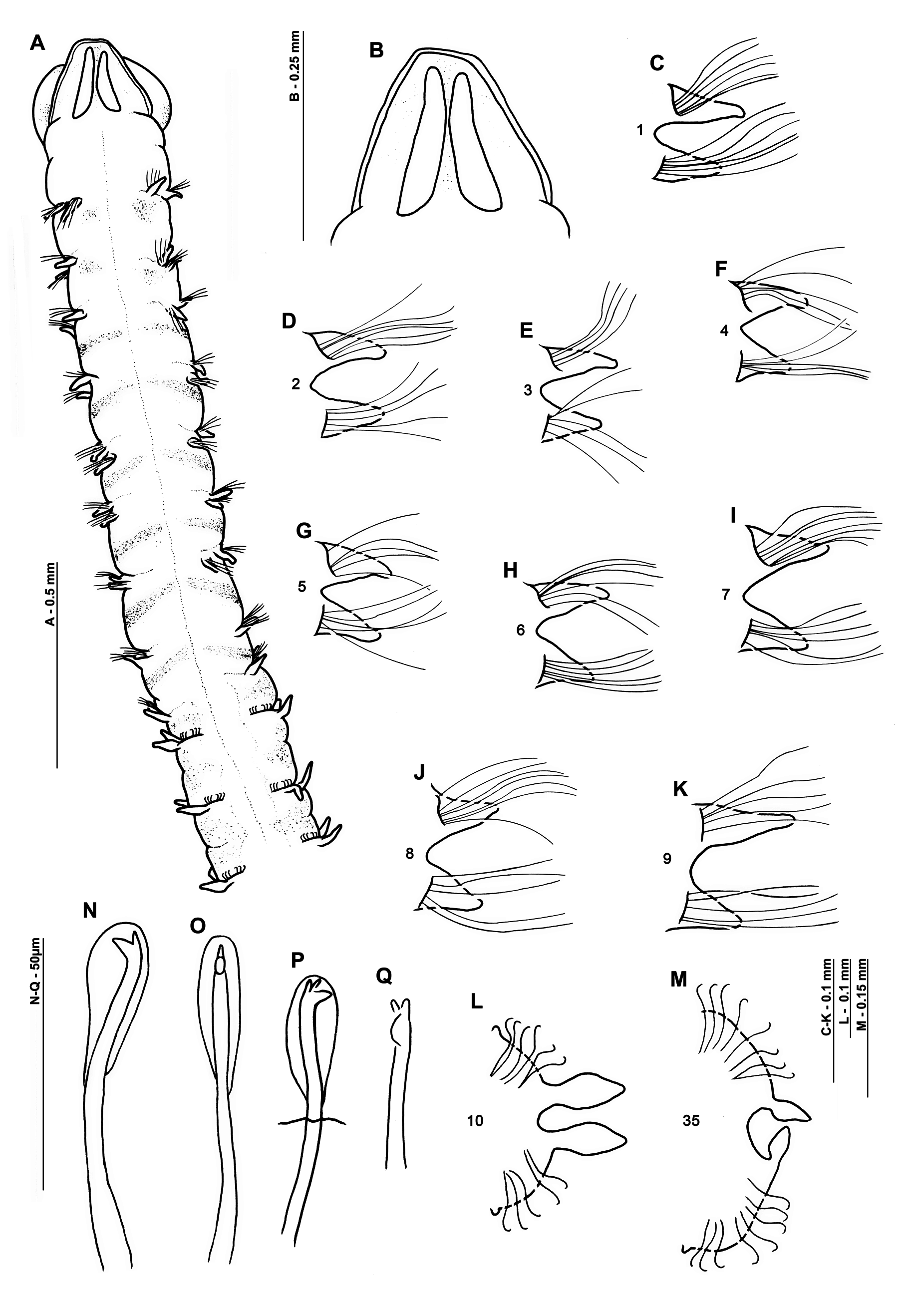
Diagnosis. Small, slender species. Prostomium width similar to length, sub-trapezoidal, without prostomial horns. Notopodia and neuropodia of chaetigers 1–9 with broad triangular postchaetal and low prechaetal lamellae. Thoracic chaetigers with capillary chaetae. Abdominal lateral lamellae slender foliaceous, slightly basally constricted. Hooded hooks bidentate, in 2 groups, vis-à-vis. No lateral pouches. Pygidium with a pair of long, slender cirriform lateral cirri.
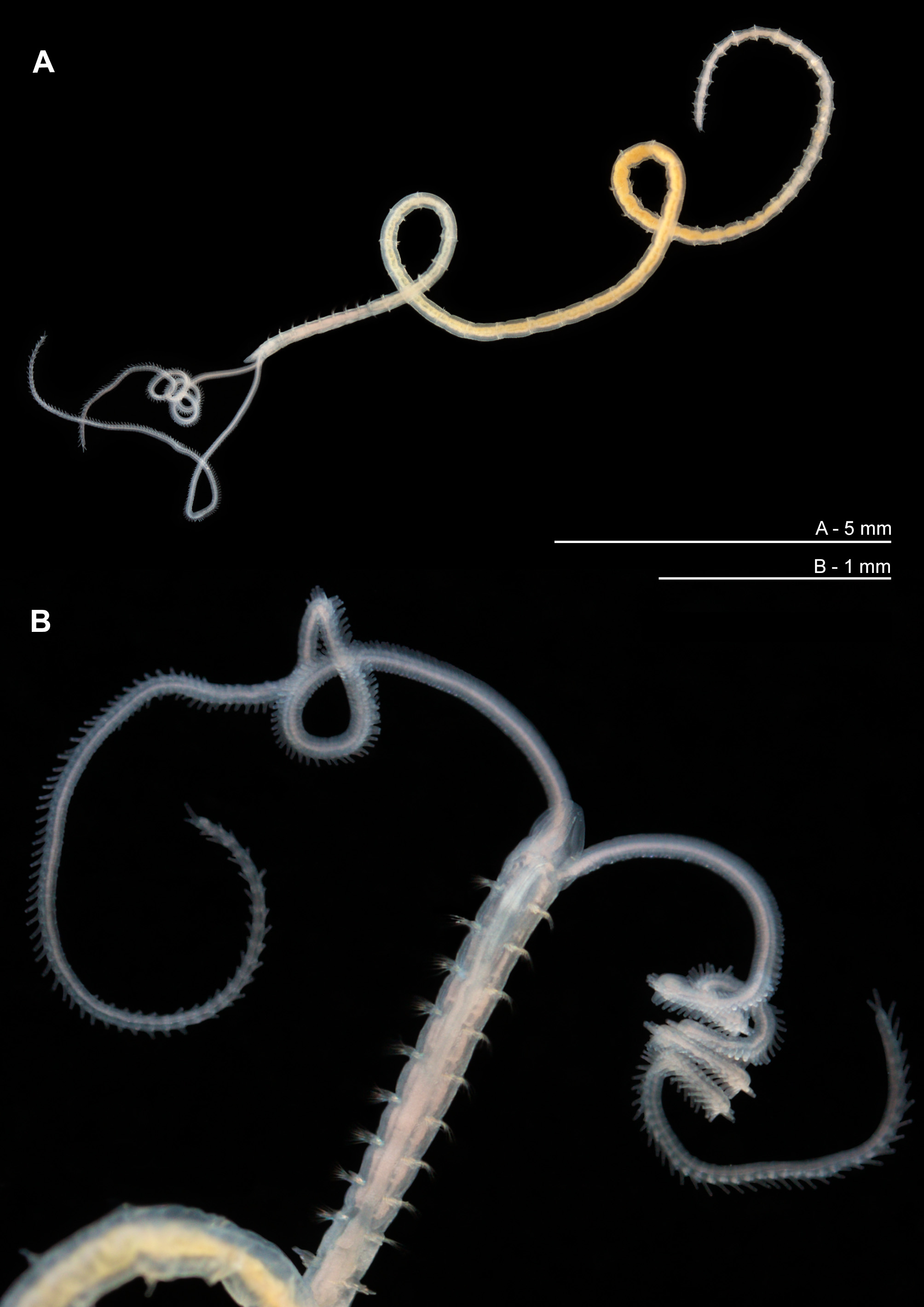
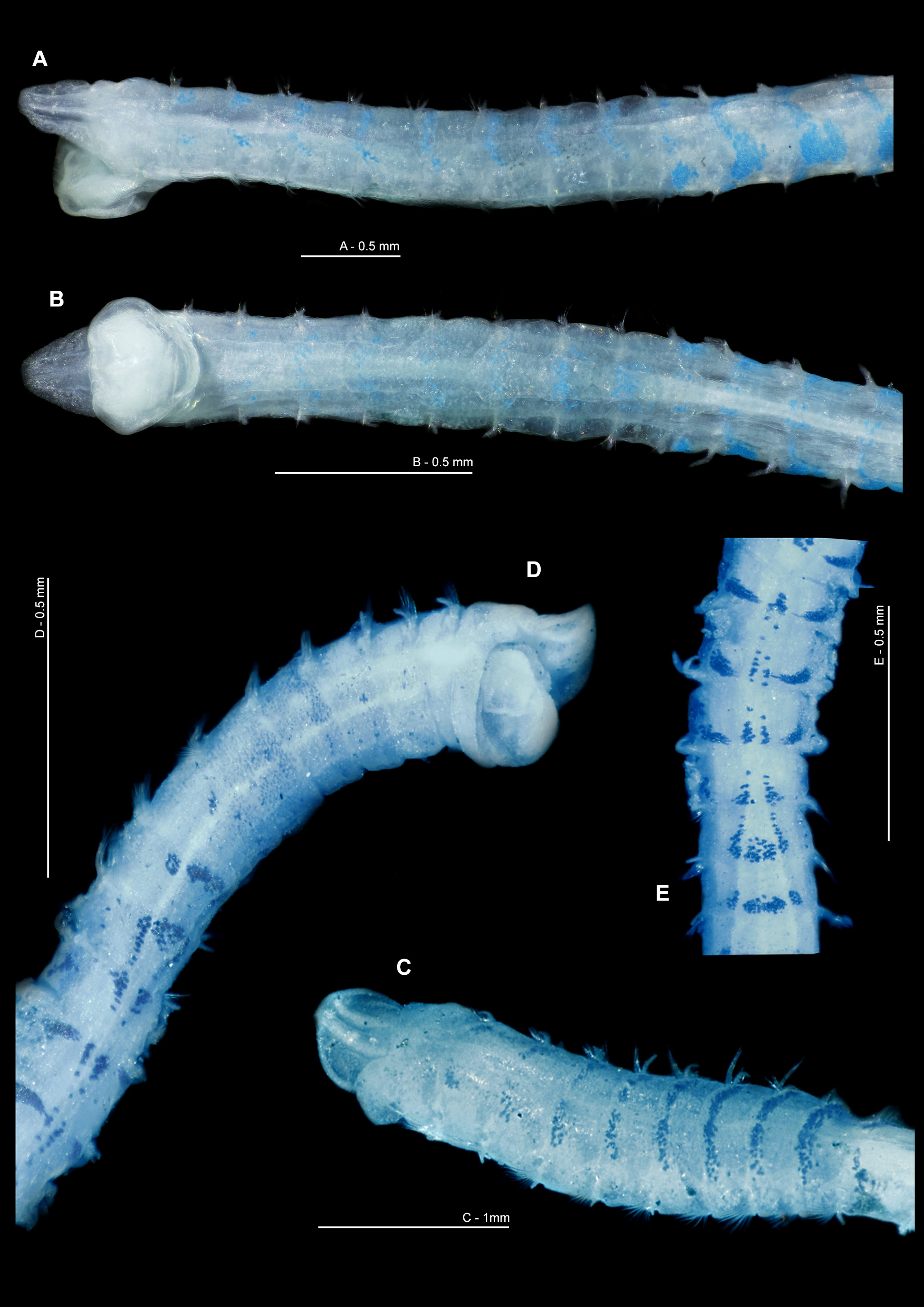
Description. Small, slender species ( Figs 3 View FIGURE 3 , 5 View FIGURE 5 , 9 View FIGURE 9 ), thorax of similar width to abdomen. Holotype, incomplete in two fragments: prostomium 0.3 mm wide, 0.25 mm long; thorax 1.9 mm long (including prostomium), 0.3 mm maximum width; abdomen 0.4 mm wide, total length of anterior fragment 5.5 mm for ~19 chaetigers; abdominal fragment 5 chaetigers long. Fluid preserved USNM 52510 specimen (af): prostomium 0.25 mm wide, 0.3 mm long; thorax 1.55 mm long (including prostomium), 0.3 mm maximum width; abdomen 0.25 mm wide; total length 3.25 mm for ~17 chaetigers. Slide mounted USNM 52510 specimen (almost complete): prostomium 0.3 mm wide, 0.25 mm long; thorax 0.3 mm wide; chaetigers 1–8 1.54 mm in length (including prostomium); abdomen 0.25 mm wide, 8.95 mm long; approximate total length 12 mm for 42 chaetigers. Total length of other entire specimens from Southern Irish and Celtic Seas (19) 9.7–16.5 mm long, for 51–67 chaetigers. N.B. all width measurements excluding parapodia.
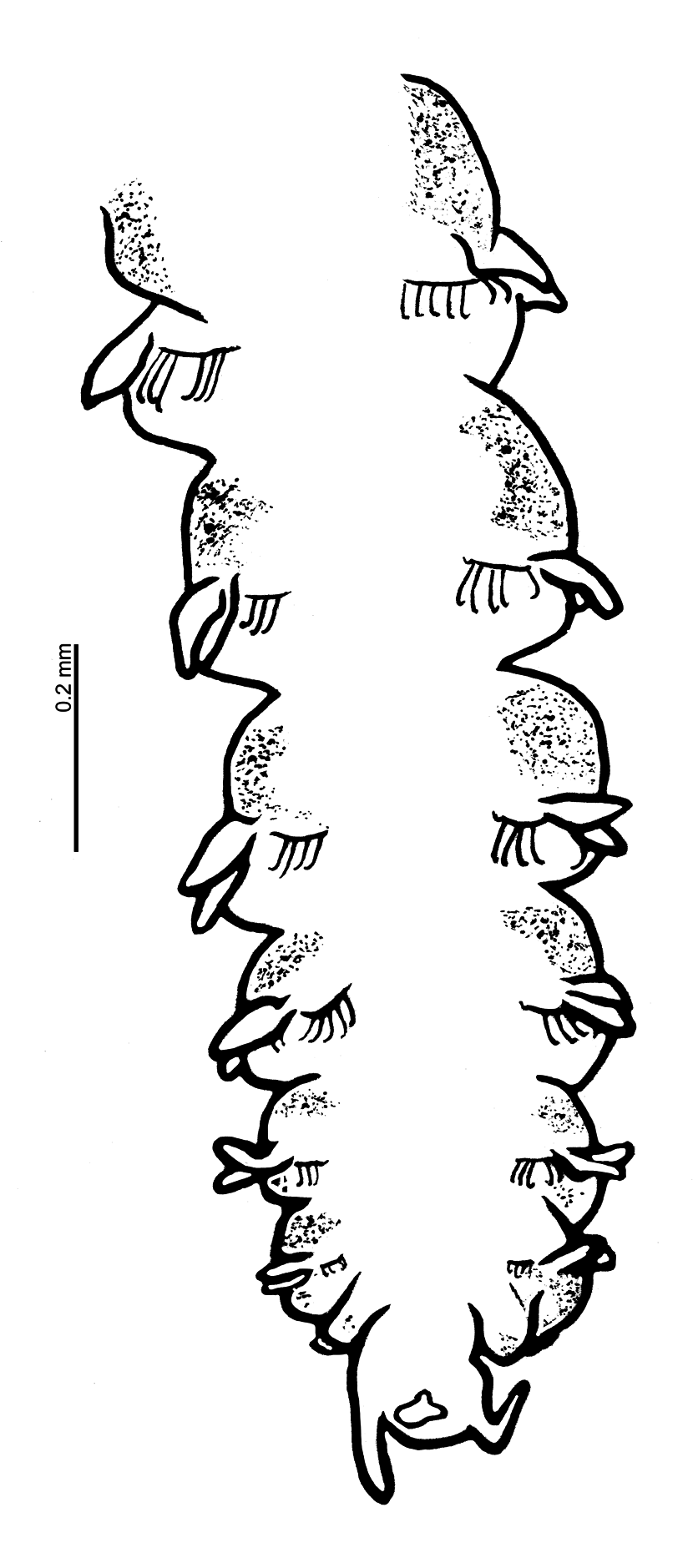
Prostomium of similar width to length (L:W ratio 0.8–1.2, Figs 3A, 3B View FIGURE 3 , 5B View FIGURE 5 , 9 View FIGURE 9 A–C), sub-trapezoidal, anterior margin smooth and straight. Lateral margins rounded in freshly preserved material ( Fig. 9C View FIGURE 9 ), those of the holotype straighter, although appearing as though laterally compressed inwards. Prostomial horns and eyes absent. Two longitudinal dorsal muscular ridges, diverging at both ends ( Fig. 3B View FIGURE 3 ); no obvious prostomial markings either side. No palps retained on holotype, those of Southern Irish and Celtic Seas arising ventrolaterally from base of prostomium, long, reaching chaetigers 17–24 (3–5.44 mm long). Non-papillated region reaching approximately chaetiger 2. Thereafter, each palp densely papillated basally, with one row medially and distally either side of a median non-papillated longitudinal line (previously termed groove). Distal papillae longer and spikier in appearance ( Fig. 5B View FIGURE 5 ). Proboscis of holotype and fluid preserved anterior fragment (USNM 52510; Figs 9A, 9B View FIGURE 9 ) partially everted, oval to heart-shaped, appearing relatively smooth.
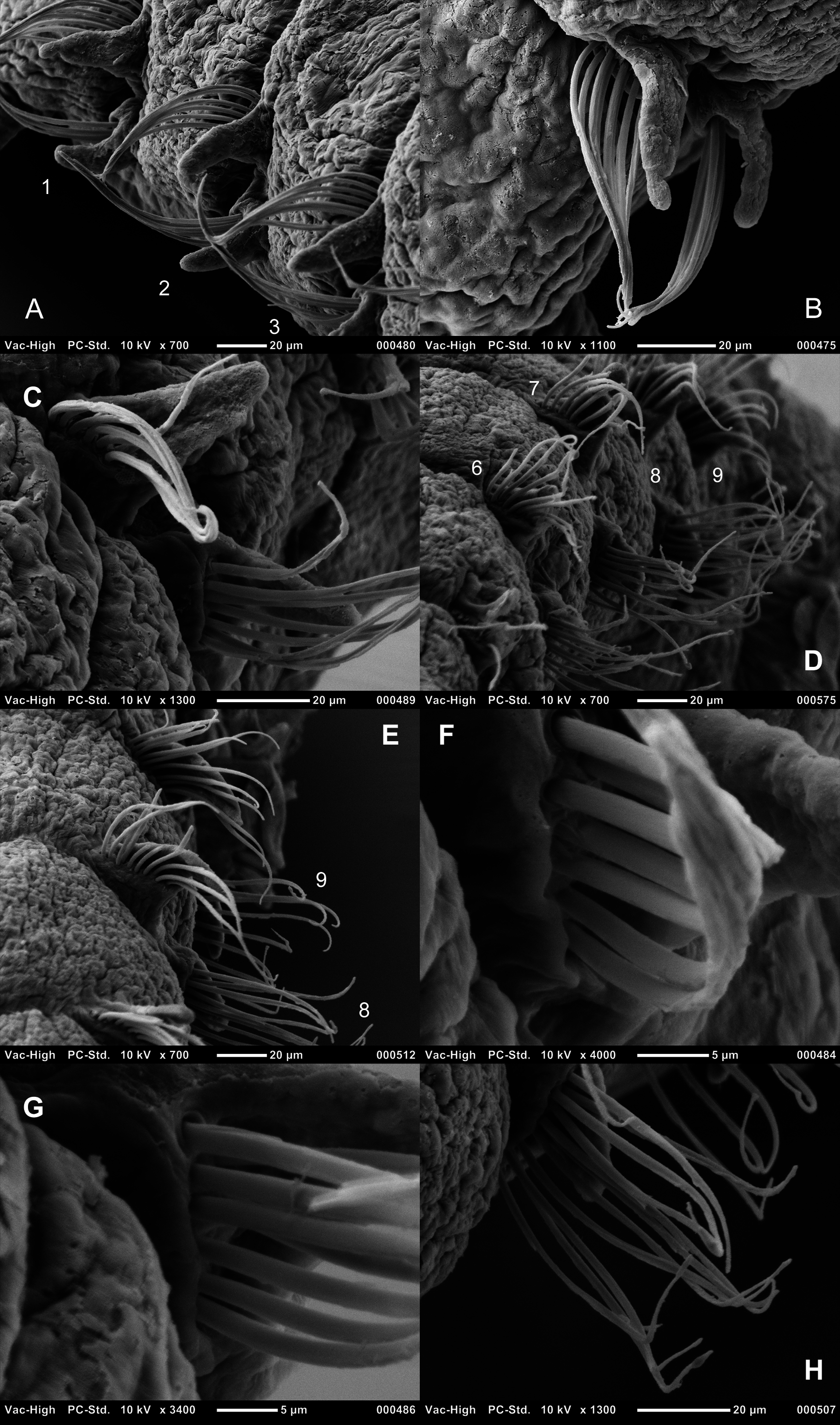
Achaetous first segment approximately one and a half times the size of chaetiger 1 ( Fig. 3A View FIGURE 3 ). Chaetigers 1–8 similar ( Figs 3 View FIGURE 3 C–J, 4A–D, 6A–E); parapodia biramous. Noto- and neuropodial postchaetal lamellae broad based triangular of similar length in both rami, smooth-edged, with low indistinct prechaetal lamellae. Chaetiger 9: parapodia similar to preceding chaetigers; prechaetal lamellae more vertically positioned, notopodial lamellae marginally longer than neuropodial ( Figs 3K View FIGURE 3 , 6E View FIGURE 6 ). No superior dorsal processes present on thoracic chaetigers.
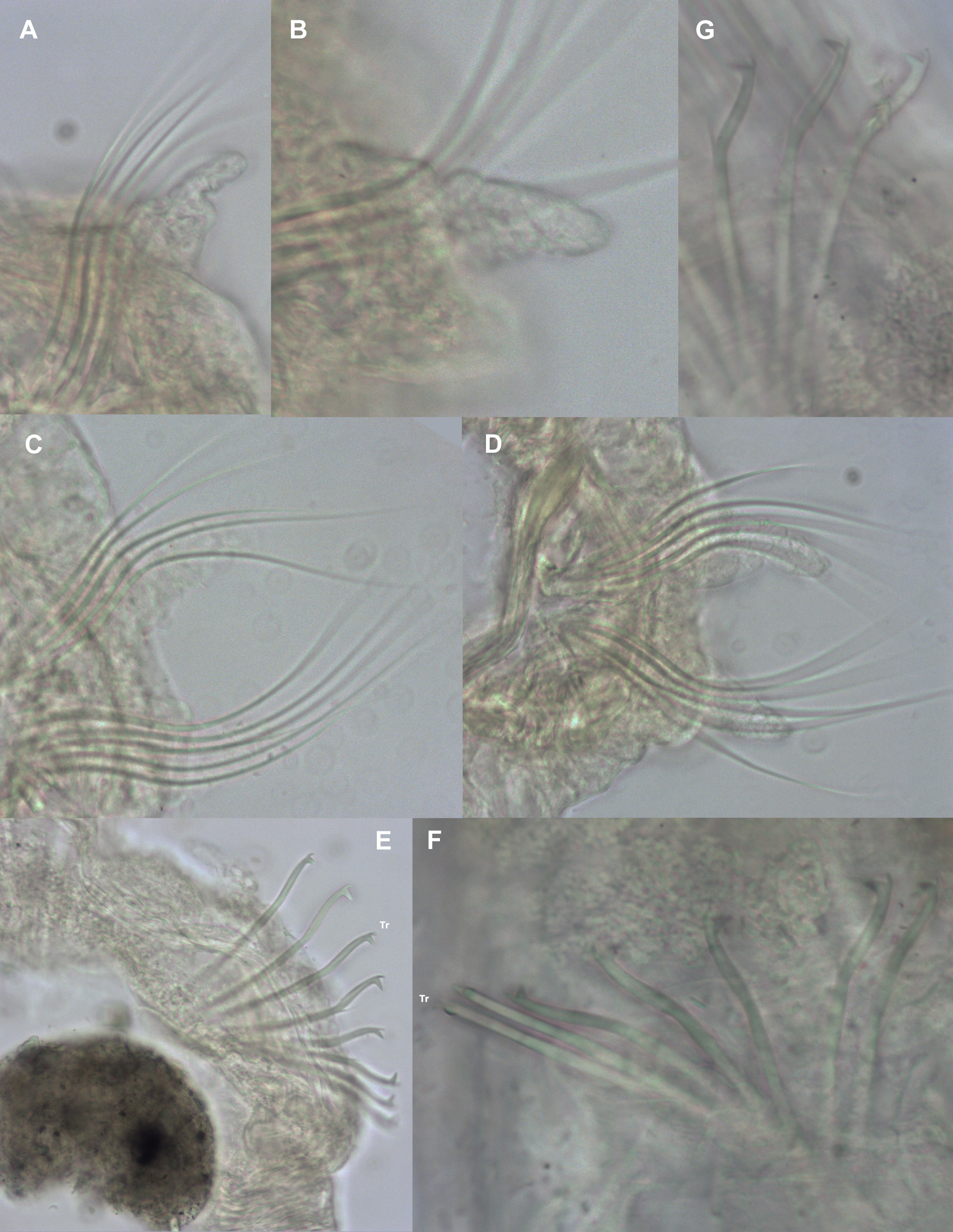
Thoracic chaetae all smooth edged unilimbate simple capillaries ( Figs 4 View FIGURE 4 A–D, 6). Slide mounted USNM 52510 specimen with 5–7 notochaetae and 5–6 neurochaetae. Other specimens from the Irish Sea with approximately 8– 11 notochaetae (becoming more numerous towards chaetiger 8) and 7–11 neurochaetae ( Table 1). Noto- and neurochaetae of a similar length.
Abdominal chaetigers with slender foliaceous lateral lamellae, of about equal size in both rami, slightly basally constricted ( Figs 3L, 3M View FIGURE 3 , 7A, 7B View FIGURE 7 ); becoming increasingly lanceolate ( Figs 7C, 7D View FIGURE 7 ) towards posterior region. Lateral lamellae do not extend postchaetally behind chaetal rows; small inferior and superior processes present at inner margins ( Figs 3L, 3M View FIGURE 3 , 7B, 7G View FIGURE 7 ), more obvious in anterior abdomen. Chaetigers twice as long as wide from around chaetiger 13 for holotype.
Abdominal chaetae bidentate hooded hooks with a secondary tooth above the main fang ( Figs 3N, 3O View FIGURE 3 , 4 View FIGURE 4 E–G, 7E–H), emerging from definite ridge ( Figs 3L, 3M View FIGURE 3 , 4E View FIGURE 4 , 7 View FIGURE 7 ). Hooks in two approximately equal groups, vis-à-vis. An occasional tridentate hooded hook observed ( Figs 3 View FIGURE 3 P–Q, 4E: 3 rd hook from lamella, 4F: outermost hook on lefthand side). Slide mounted specimen (USNM 52510) with around 6 hooks per ramus in anterior abdomen, increasing to around 10 hooks per ramus in the medial and posterior abdomen. Irish Sea specimens with approximately 6–10 hooks per ramus in anterior and medial abdomen; increasing to approximately 10–12 on posterior chaetigers ( Fig. 7 View FIGURE 7 ). Angle between main fang and secondary tooth between ~75°–105° ( Figs 3N View FIGURE 3 , 4 View FIGURE 4 E–G, 7E–H, Table 2). Lateral pouches (anteriorly and posteriorly open) absent. Posterior chaetigers tapering towards pygidium ( Fig. 8 View FIGURE 8 ). Holotype posteriorly incomplete. Pygidia of specimens from Irish and Celtic Seas with two long, slender, lateral anal cirri; small anus, ventrally located ( Fig. 8 View FIGURE 8 ).
Colour. Live material generally pale pinkish white; colouration of gut largely conspicuous, yellow/orange ( Fig. 5 View FIGURE 5 ). Fluid preserved material white in colour: USNM 52510 specimen more translucent than holotype and comparative material, although smaller in size. Speckled white patches (more conspicuous and yellow on the former specimen due to its translucent nature) present dorsally on chaetigers 1–3, adjacent to parapodia, becoming more distinct arched bands on chaetigers 4–9, only slightly interrupted medially ( Fig. 3A View FIGURE 3 ). Similar speckled areas ventrally, fainter, becoming more visible from chaetiger 6. Dorsal bands continuing into anterior abdomen. Colouration previously noted by Eliason: “Tieren läuft indessen zwischen und ein wenig hinter den vorderen Notopodien ein bisweilen abgebrochenes Band von graugelben Zellanhäufungen” (approximate translation: “In between, and slightly behind the anterior notopodia, an occasional broken band of greyish-yellow cell accumulations are present”). Speckled bands not as conspicuous in live material ( Fig. 5B View FIGURE 5 ), however, more noticeable in Rose Bengal and methyl green stained specimens. Large yellow/cream abdominal interparapodial patches extending the length of the abdomen. Methyl green staining pattern most noticeable on dorsal markings as noted above, but gaining intensity along the length of the thorax ( Fig. 9C View FIGURE 9 ) and apparent on abdominal interparapodial patches. Ventral staining on chaetigers 1–5 similar albeit paler ( Figs 9B, 9D View FIGURE 9 ). Faint interparapodial patches on posterior thorax, similar to those of the abdomen ( Fig. 9D View FIGURE 9 ). Two ventral longitudinal lines span the length of the thorax, converging at the thoracic/abdominal junction ( Figs 9D, 9E View FIGURE 9 ). Methyl green stain persisting diffusely over much of the thorax particularly between chaetigers 4–6 for some time after initial staining. Rose Bengal staining pattern similar, but ventral longitudinal lines less conspicuous.
Habitat. Occurs in muddy and sandy sediments ( Eliason, 1962; authors pers. comm.), although the former author noted specimens occurring in only sand or sand with a little mud (Skagerrak and the Kattegatt sea area). Comparative material from the current study records the species from sandy mud, muddy sand, sand and sandy gravel, from depths of 15– 160 m. Eliason (1962) recorded depths of at least 13 to 53 m, however, Fiege et al. (2000) includes records of up to 1000 m depth in the Mediterranean. The Global Biodiversity Information Facility’s ( GBIF Secretariat, 2017) database contains records of M. cf. minuta over 4000 m deep from Greenland. However, the deeper records of the species have not been verified. Mortimer & Mackie (2014) described M. minuta as an offshore muddy sediment species, primarily based on personal observations from the Irish Sea.
Distribution. Current confirmed distribution from Norway to off West Africa (Marine Invertebrates of Western Africa Project, University Museum of Bergen, J.A. Kongsrud and second author pers. comm.). Further records from Ireland ( Guiry & Guiry, 2011), the Mediterranean Sea ( Fiege et al., 2000; Faulwetter, 2010) the North Atlantic Ocean ( Bellan, 2001; Iberfauna, 2008) and Spain (Ramos, 2010) also exist.
Diet. Diatoms, silicoflagellates and minimal amounts of sand. Additionally, several fragments presumed to be of animal origin i.e. ostracods and bivalve larvae, observed for slide mounted specimen (USNM 52510, Fig. 10 View FIGURE 10 ).
Remarks. The original description of M. minuta includes material from Eliason (1920) and Thorson (1946), and a further three specimens (“Neue Funde”) from the Öresund area. One collected in 1946 from 18 m (Stn 0 60 68) and a further two collected in 1961 from 16 m (Stn 0 68 69), one of which was designated as the holotype (Polych. 11491). In a redescription of M. papillicornis, Jones (1977) listed three specimens as paratypes of M. minuta (see p252, footnote), two housed at the Gothenburg Natural History Museum (Polych. 9700 from Skagerrak & Polych. 11492 from Öresund) and one in the Smithsonian (USNM 52510, from Öresund), despite Eliason never formally designating any within the original description. Fiege et al. (2000) noting Jones’ remarks listed the latter as paratype material, and used it as the basis of a drawing of the anterior region. However, it was never listed as type material in Smithsonian catalogues (Kathryn Ahlfeld pers. comm.), and no collection date was recorded for verification with the original description. Records of NHMG indicate several paratype specimens of the species (including that mentioned by Jones, Polych. 11492) and these may have been the specimens referenced in regards to Skagerrak and Kattegatt in the original description. However, these specimens were all lost in a loan to France back in 1972 (pers. comm. Kennet Lundin, NHMG), including the Skagerrak specimen (Polych. 9700) mentioned by Jones (1977, but which was never recorded as a paratype in NHMG records). The holotype is unfortunately in a poor condition, something also noted by Fiege (pers. comm.). In light of this and in the absence of any formally designated paratype material, the Smithsonian specimens (USNM 52510) were chosen to be drawn and photographed within this re-description. These specimens were originally identified by Eliason himself and come from the type locality (18 m depth). We share Jones’ (1977) opinion that the slide-mounted specimen may have been the basis for the figures included in the original description.
Reexamination of the species has revealed further morphological features not previously described by Eliason, including prostomial and lamellar shape and the presence of sporadic tridentate hooded hooks. Examination of comparative material from British and Irish waters has allowed further features, such as the pygidium and palps, to be more fully described. These specimens agree well with the holotype in most respects, although a small variation in chaetal number exists; the slightly larger comparative material possessing marginally more thoracic chaetae. One of the notable differences compared with the partial redescription by Fiege et al. (2000) concerns prostomial shape. At that time it was figured with a rounded anterior margin, whilst reexamination of the same anterior fragment shows it to be straight, in agreement with the holotype and other observed live material. In general, the prostomial width is similar to its length, with slightly rounded lateral edges. Observed variations in L:W ratios are in most part due to the lateral compression of prostomial margins, particularly in older specimens.
This redescription has highlighted the morphological similarities between M. minuta and M. filiformis . Both species possess distinctive transverse bands, particularly in the thoracic region, and have a similar prostomial shape. Whilst they can be distinguished in terms of presence/absence of thoracic superior dorsal processes, dentition of abdominal hooded hooks and lamellae shape, this morphological closeness is likely to have caused previous taxonomic confusions, particularly with juvenile specimens (see discussion).
Magelona minuta View in CoL belongs to a ‘ Magelona View in CoL papillicornis’ group of species, which are all small and slender, with sub-triangular prostomia of similar width to length, and possess triangular postchaetal thoracic lamellae without superior dorsal processes. Members of the group are: M. papillicornis View in CoL , M. californica View in CoL , M. minuta View in CoL , Magelona pettiboneae Jones, 1963 View in CoL , Magelona pygmaea Nateewathana & Hylleberg, 1991 View in CoL and Magelona fauchaldi Shakouri, Mortimer & Dehani, 2017 View in CoL . Magelona minuta View in CoL is distinguished from the latter two species by dentition of the hooded hooks, possessing bidentate as opposed to tridentate and polydentate respectively. Jones (1977) stated that the particularly low and broadly attached triangular thoracic lamellae of M. minuta View in CoL could be used to further differentiate this species from remaining bidentate members.



| NHMG |
Goteborgs Naturhistoriska Museet |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Magelona minuta Eliason, 1962
| Mills, Kimberley & Mortimer, Kate 2018 |
Magelona fauchaldi
| Shakouri, Mortimer & Dehani 2017 |
Magelona pygmaea
| Nateewathana & Hylleberg 1991 |
M. papillicornis
| , Jones 1977 |
Magelona pettiboneae
| Jones 1963 |
Magelona minuta
| Eliason 1962 |
M. minuta
| Eliason 1962 |
Magelona minuta
| Eliason 1962 |
M. minuta
| Eliason 1962 |
M. filiformis minuta
| Wilson 1959 |
Magelona rosea
| : McIntyre 1958 |
Magelona papillicornis var. rosea:
| Thorson 1946 |
M. californica
| Hartman 1944 |
Magelona rosea:
| Eliason 1920 |
Magelona rosea:
| Southern 1914 |
Magelona
| F. Muller 1858 |