
Pheretima quincunxia, James, 2004
|
publication ID |
https://doi.org/ 10.5281/zenodo.4618925 |
|
persistent identifier |
https://treatment.plazi.org/id/9918E954-FFB2-E06A-0984-F93957D6FDB8 |
|
treatment provided by |
Carolina |
|
scientific name |
Pheretima quincunxia |
| status |
sp. nov. |
Pheretima quincunxia , new species
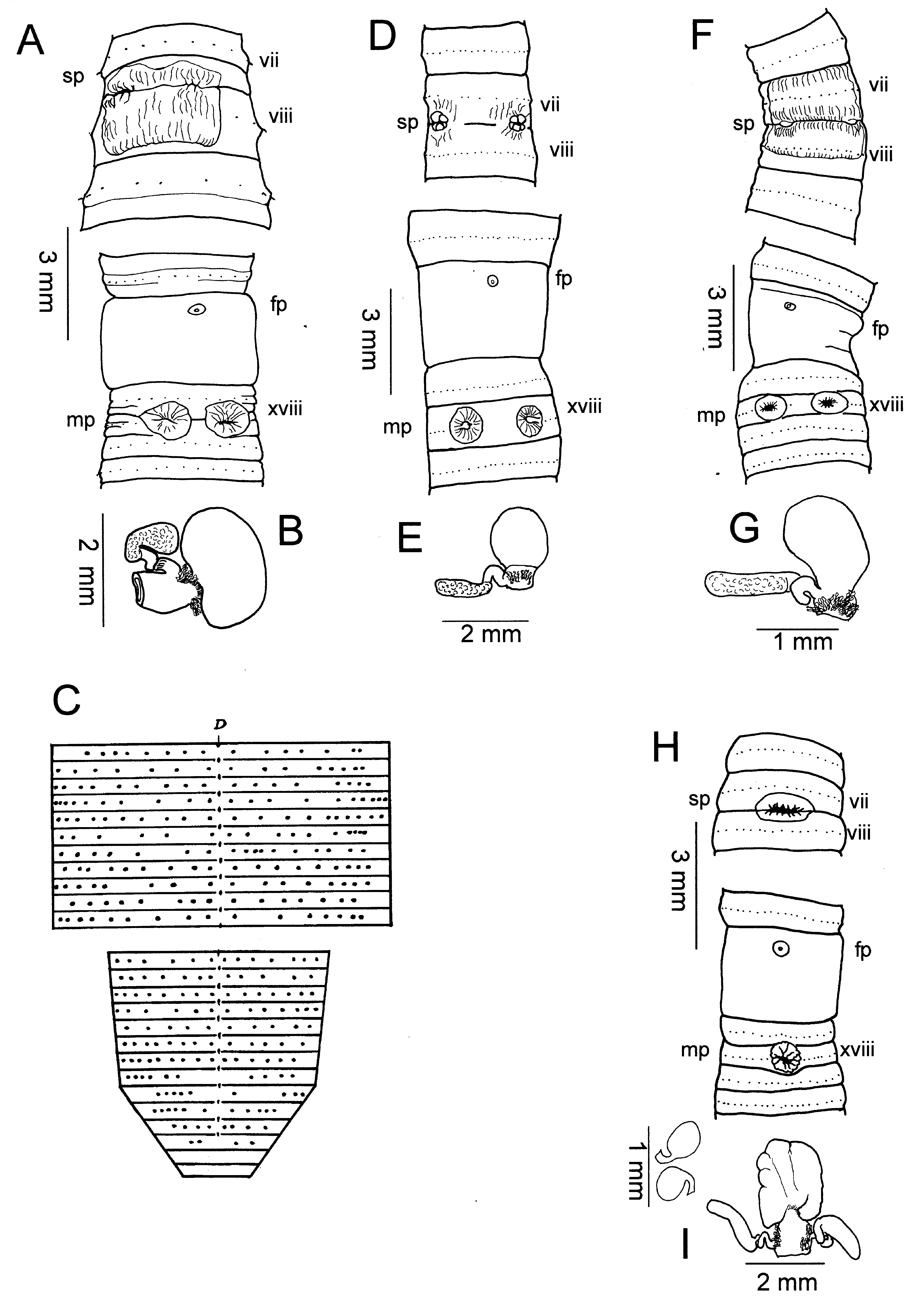
( Figs. 2 View Fig A-C)
Material examined. – Holotype - adult ( NMA 003978 ) Philippines, Mindanao Island, Bukidnon Province, Mt. Kitanglad Range , 18.5 km S, 4 km. E of Camp Philips, 2250 m. elevation, coll. L. Heaney, 2 Apr.1993.
Paratypes – four adults ( FMNH 011064 View Materials ) Philippines, Mindanao Island, Bukidnon Province, Mt. Kitanglad Range , 18.5 km S, 4 km. E of Camp Philips, 2250 m. elevation, coll. L. Heaney, 2 and 4 Apr.1993 .
Etymology. – The species is named for its approximately quincunxial setal arrangement.
Description. – Worms unpigmented, body 65-86 x 3.9 mm (vii), 4.8 (x), 4.6mm (xxv), 74-82 segments; body cylindrical in cross-section. First dorsal pore 12/13, spermathecal pores with thick lips paired in 7/8, 0.16 circumference apart, female pore single in xiv, openings of copulatory bursae paired in xviii, 0.13 circumference apart in 2 nd or 3 rd setal line, 0 (4 individuals) or 2 (1) setae between openings. Behind clitellum setae of adjacent segments offset, irregular, approaching quincunxial arrangement, 16-20 setae on vii, 20-30 setae on xx, 10-15 in tail segments, there widely spaced dorsally, laterally, with 3-5 closely spaced setae on ventral side ( Fig. 2C View Fig ); dorsal gaps irregular, no ventral gaps. Clitellum annular xiv-xvi; rugose epidermally thickened rectangular area covering ventral post-setal third of vii, most of viii ( Fig. 2A View Fig ).
Septa 4/5- 9/10 thin, 10/11-12/13 muscular. Nephridia in dense tufts on anterior faces of 5/6, 6/7; nephridia of postclitellar segments in pre-, post-septal rows near septum-body wall junction.
Large gizzard in viii, esophagus with vertical lamellae x-xiii, intestinal origin 1/2xvi, simple caeca originating in xxvii, extending forward to xxv; typhlosole rudimentary; 32 longitudinal vessels in intestinal wall.
Hearts x-xiii esophageal, commissural vessels vi, vii, ix lateral; viii to gizzard; ventral vessel with branches to body wall in viii, ix. Supra-esophageal vessel x-xiv; extraesophageal vessel not seen.
Ovaries and funnels free in xiii, spermathecae paired, preseptal in vii with nephridia on spermathecal ducts; each spermatheca with large rounded ampulla filling its side of vii, single stalked diverticulum terminating in pear-shaped receptacle, stalk tightly looped, first loop closely adherent to spermathecal duct ( Fig. 2B View Fig ). Male sexual system holandric, testes and funnels enclosed in paired sacs in x, xi; seminal vesicles xi, xii; vasa deferentia free from body wall en route to ental end of prostatic ducts; each prostate racemose occupying xvii-xix, stout muscular duct entering posterior side of copulatory bursa in xviii; coelomic and exterior surfaces of copulatory bursae lacking glandular or other projections; four pads flanking opening to exterior; no welldeveloped penis, but a small protuberance varying from Vshaped fold to low conical bump with male pore on apex.
Remarks. – Pheretima quincunxia keys to the P. sangirensis ( Michaelsen, 1891) , group in Sims & Easton (1972), which once consisted of three species, P. sangirensis , P. ceramensis ( Cognetti, 1922) , and P. crassicystis ( Michaelsen, 1896) , which in turn is divided into three subspecies, P. c. chica ( Michaelsen, 1896), P. c. crassicystis ( Michaelsen, 1896) and P. c. tobeloensis ( Michaelsen, 1896). However, Michaelsen (1900) synonymized the latter two subspecies of P. crassicystis under the name P. c. crassicystis , and included all crassicystis as subspecies of P. sangirensis . A third species in the group, P. unicystis ( Lee, 1981) , was described from Vanua Tu material ( Lee, 1981). The others are from Indonesia, in the Molucca Islands, Halmahera and North Celebes (Sulawesi). I examined material of P. sangirensis and P. ceramensis , and referred to the original descriptions for further information. The characteristics of P. sangirensis are given by Michaelsen (1900) as follows: anterior and ventral setae slightly enlarged, very slight dorsal gap present, male pores from 0.17 to 0.25 circumference apart, spermathecal pores one pair in 7/8, from 0.25 to 0.28 circumference apart, septa 8/9/10 lacking, caeca either simple or with short pockets on the ventral margins, holandric with paired testis sacs, copulatory pouches simple, with short conical penis. The subspecies vary in size (60-240 mm), color, shape of the spermathecal diverticula, and in the structure of the spermathecae.
Pheretima ceramensis is from the Indonesian island of Ceram, just west of New Guinea. It is a large worm, 140- 440 mm X 8-14 mm, 60 setae per segment, dark violet dorsal pigmentation, male pores about 0.2 circumference apart and spermathecal pores slightly closer set; no setae between male pores ( Cognetti, 1922). Interior characters are given in Cognetti (1922) as: septa 8/9/10 absent, 10/11-15/16 very muscular, intestinal origin xvi, caeca simple, last hearts xiii; testes sacs paired, unconnected and subesophageal, prostate glands divided in about 12 lobes, prostate ducts muscular, bent, entering large and robust copulatory pouch. Spermathecae one pair in vii, ampulla spherical on a large but short muscular duct, diverticulum chamber elongate. My examination of the type material, whose label data agree entirely with that reported in Cognetti (1922), show some slight differences: septum 8/9 present, intestinal origin 1 / 2 xv, one seta between male pores, prostate divided in 4 main lobes.
Lee (1981) distinguished Pheretima unicystis from the previous species and P. crassicystis by the presence of only one asymmetrically placed spermatheca, spermathecal characters, body size, and intestinal origin. My examination of previously undissected material of P. ceramensis shows an intestinal origin at 1 / 2 xv, rather than xvi as reported in Cognetti, and partly relied upon by Lee (1981).
Pheretima quincunxia , P. paucisetosa and P. alba below, are the only members of the species group lacking pigment, though P. unicystis is only weakly pigmented. Pheretima quincunxia has spermathecal and male pores more narrowly spaced than in other members of the species group, except for P. paucisetosa , P. alba and P. rubida , whose spacings are comparable ( Table 2). It also has fewer setae per segment than all others in the group and the setae have an irregular arrangement approaching quincunxial (alternating position in adjacent segments). It has all anterior septa present, unlike the others except P. unicystis . The stalk of the spermathecal diverticulum is not simple as in the P. sangirensis group but is tightly looped. While the spermathecal pores are in 7/8, indicating membership in the sangirensis group, the spermathecae are in vii, a state shared with the BMNH material of P. sangirensis , all P. ceramensis , and P. unicystis . The presence of other species in the region ( P. diesmosi , P. rugosa , P. monoporata , P. paucisetosa , P. alba , P. baungonensis , P. rubida ) with the same condition would indicate that the evolution of anteriorly displaced spermathecae took place in a common ancestor of these species and perhaps also P. ceramensis . In those sangirensis group species described here with spermathecae in vii, the spermathecae are placed in the posterior half of the segment, so I refer to such spermathecae as preseptal, indicating their unusual position within the segment. In all these cases the septal insertions are in line with the external intersegmental boundaries, with no evidence of displacement of septa either posteriorly or anteriorly. Therefore one must conclude that the spermathecae have migrated forwards, and that the simplest interpretation is that this change took place once in a common ancestor of the species in question.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |